Effect of Metallothionein-III on Mercury-Induced Chemokine Gene Expression

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Exposure Procedures
2.2. Real-Time Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
2.3. Analysis of Mercury Concentrations in Tissues
2.4. Statistical Analyses
3. Results
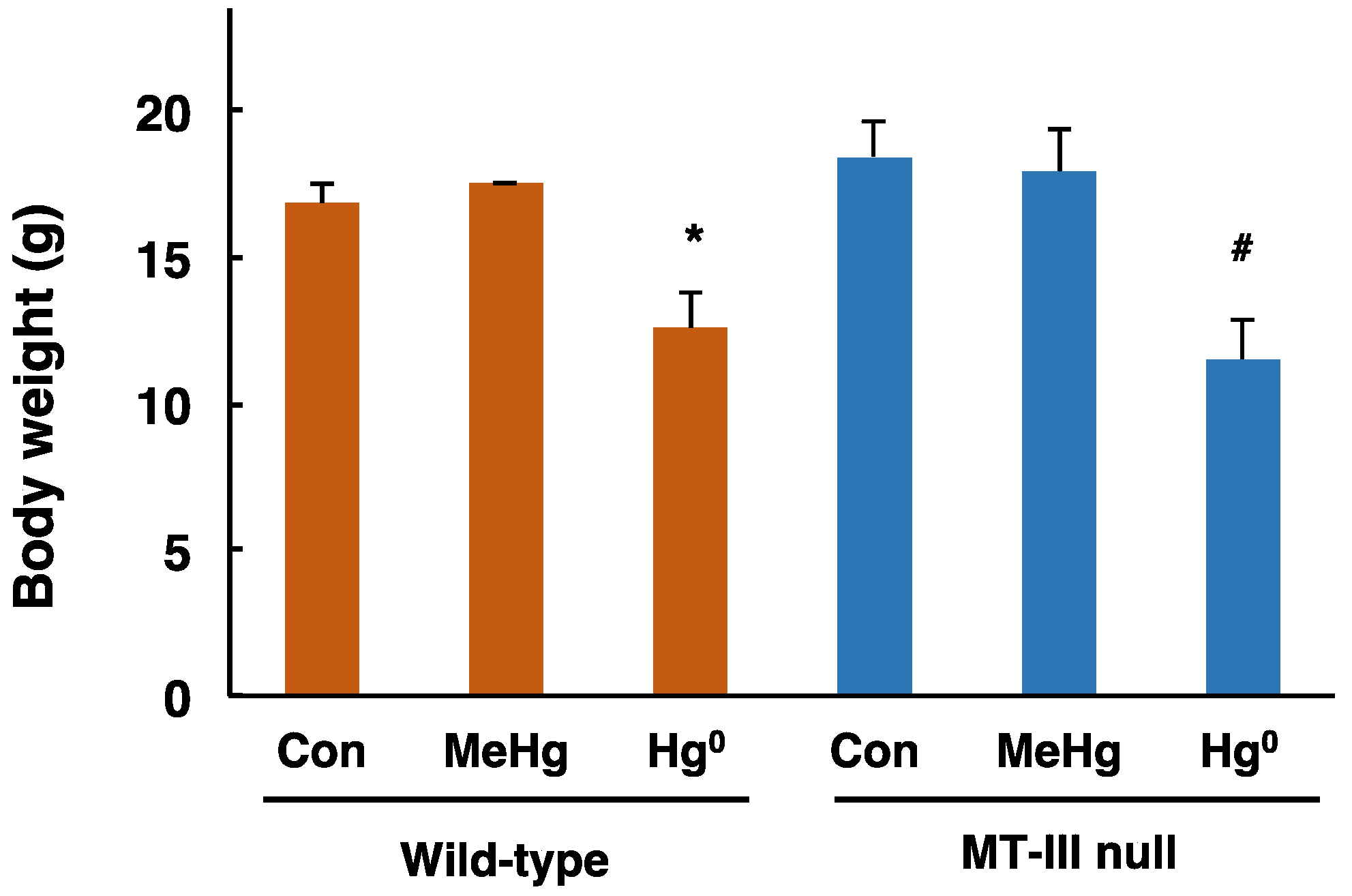
3.1. Body Weight Changes
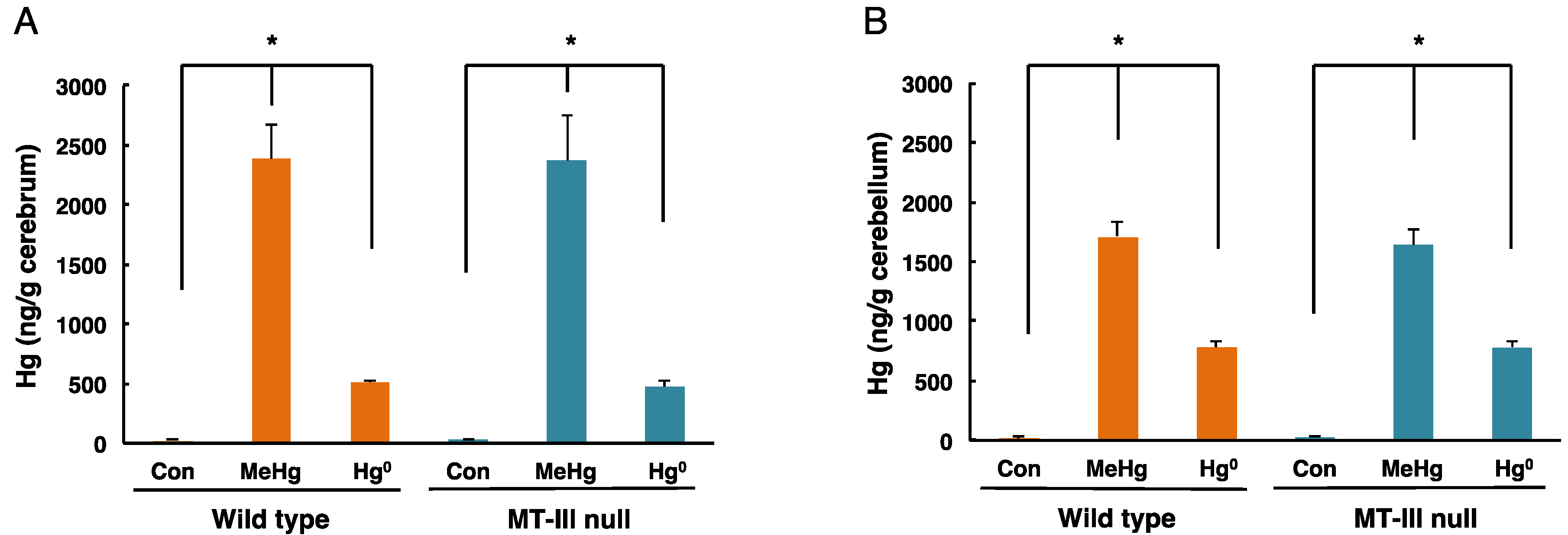
3.2. Total Mercury Concentrations in the Cerebrum and Cerebellum
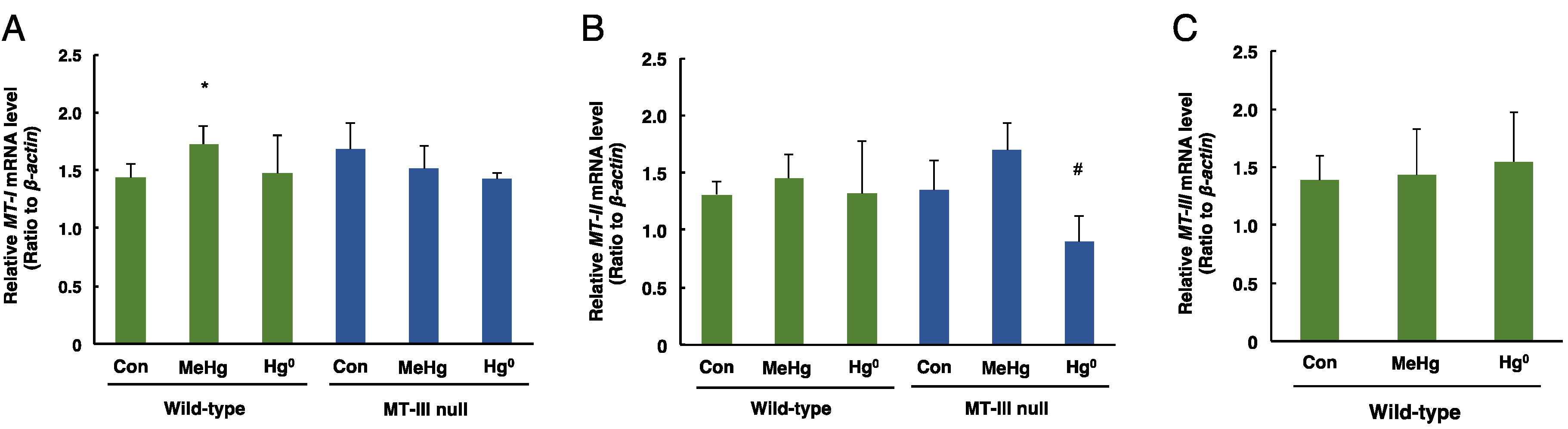
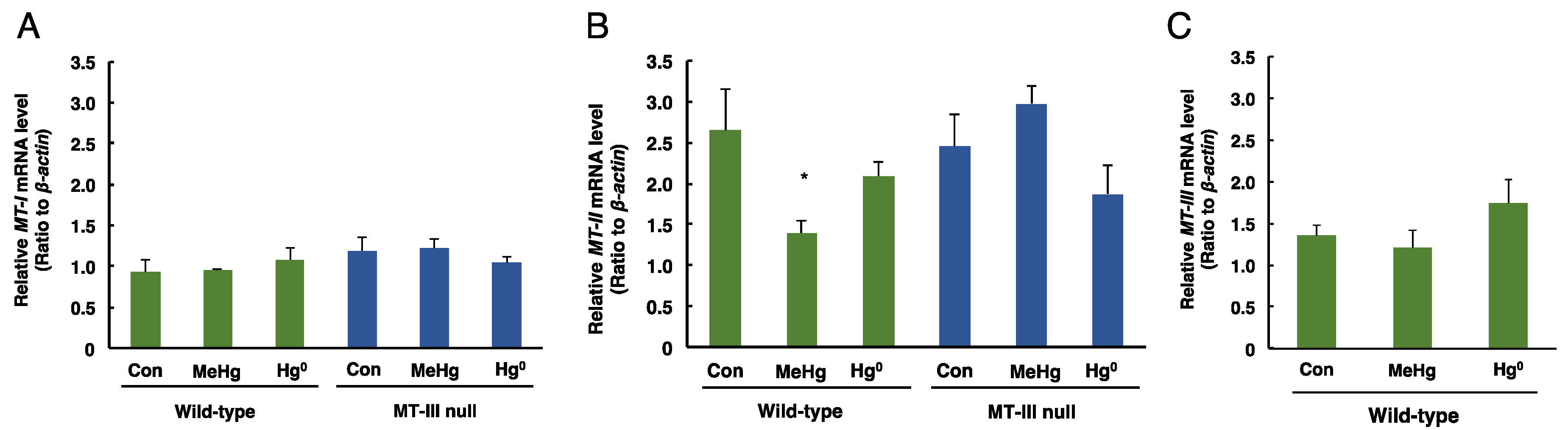
3.3. The Levels of MT-I, MT-II, MT-III mRNAs in the Cerebrum and Cerebellum after Mercury Exposure
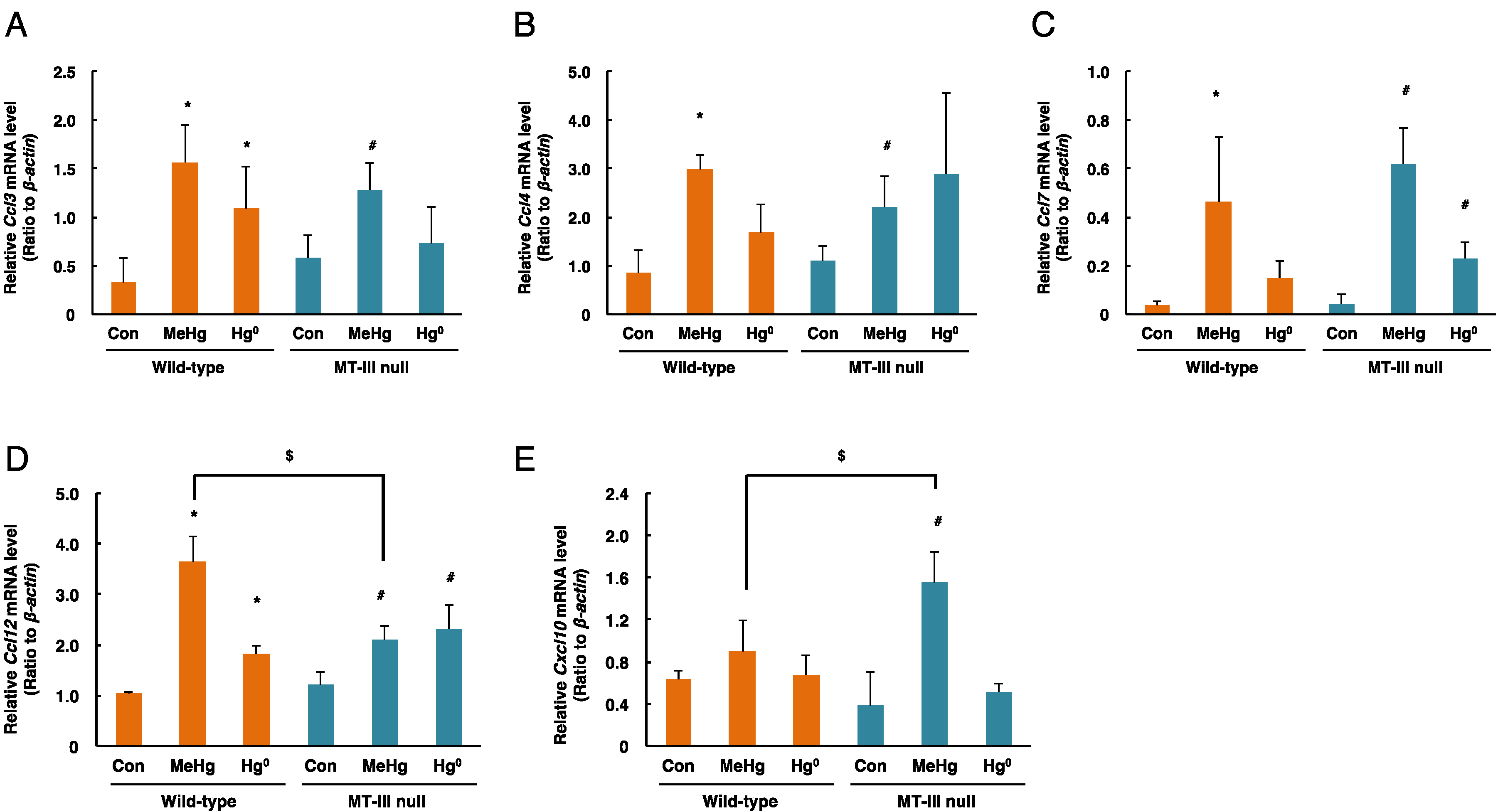
3.4. Changes in Expression of Chemokine Genes in the Cerebrum in Response to Mercury Exposure
3.5. Changes in Expression of Chemokine Genes in the Cerebellum after Mercury Exposure
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lee, J.Y.; Ishida, Y.; Kuge, S.; Naganuma, A.; Hwang, G.W. Identification of substrates of F-box protein involved in methylmercury toxicity in yeast cells. FEBS Lett. 2015, 589, 2720–2725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.Y.; Ishida, Y.; Takahashi, T.; Naganuma, A.; Hwang, G.W. Transport of pyruvate into mitochondria is involved in methylmercury toxicity. Sci. Rep. 2016, 6, 21528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tokumoto, M.; Lee, J.Y.; Shimada, A.; Tohyama, C.; Satoh, M. Glutathione has a more important role than metallothionein-I/II against inorganic mercury-induced acute renal toxicity. J. Toxicol. Sci. 2018, 43, 275–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, N.; Yasutake, A.; Marumoto, M.; Hirayama, K. Methylmercury inhibits electron transport chain activity and induces cytochrome c release in cerebellum mitochondria. J. Toxicol. Sci. 2011, 36, 253–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, J.; Nakamachi, T.; Ogawa, T.; Naganuma, A.; Nakamura, M.; Shioda, S.; Nakajo, S. Characterization of antioxidant protection of cultured neural progenitor cells (NPC) against methylmercury (MeHg) toxicity. J. Toxicol. Sci. 2009, 34, 315–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grandjean, P.; Weihe, P.; Jørgensen, P.J.; Clarkson, T.; Cernichiari, E.; Viderø, T. Impact of maternal seafood diet on fetal exposure to mercury, selenium, and lead. Arch. Environ. Health 1992, 47, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Grandjean, P.; Weihe, P.; White, R.F.; Debes, F.; Araki, S.; Yokoyama, K.; Murata, K.; Sørensen, N.; Dahl, R.; Jørgensen, P.J. Cognitive deficit in 7-year-old children with prenatal exposure to methylmercury. Neurotoxicol. Teratol. 1997, 19, 417–428. [Google Scholar] [CrossRef]
- Hinton, J.J.; Veiga, M.M.; Veiga, A.T.C. Clean artisanal gold mining: A utopian approach? J. Clean. Prod. 2003, 11, 99–115. [Google Scholar] [CrossRef]
- Veiga, M.M.; Maxson, P.A.; Hylander, L.D. Origin and consumption of mercury in small-scale gold mining. J. Clean. Prod. 2006, 14, 436–447. [Google Scholar] [CrossRef]
- Bose-O’Reilly, S.; Lettmeier, B.; Gothe, R.M.; Beinhoff, C.; Siebert, U.; Drasch, G. Mercury as a serious health hazard for children in gold mining areas. Environ. Res. 2008, 107, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Feng, X.; Qiu, G.; Shang, L.; Wang, S. Mercury exposure in the population from Wuchuan mercury mining area, Guizhou, China. Sci. Total Environ. 2008, 395, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Chan, P.H.Y.; Chan, M.H.M.; Li, A.M.; Cheung, R.C.K.; Yu, X.T.; Lam, H.S. Methylmercury levels in commonly consumed fish and methylmercury exposure of children and women of childbearing age in Hong Kong, a high fish consumption community. Environ. Res. 2018, 166, 418–426. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Nakai, K.; Sugawara, T.; Nakamura, T.; Ohba, T.; Shimada, M.; Hosokawa, T.; Okamura, K.; Sakai, T.; Kurokawa, N.; et al. Neurobehavioral effects of prenatal exposure to methylmercury and pcbs, and seafood intake: Neonatal behavioral assessment scale results of tohoku study of child development. Environ. Res. 2010, 110, 699–704. [Google Scholar] [CrossRef] [PubMed]
- Tatsuta, N.; Murata, K.; Iwai-Shimada, M.; Yaginuma-Sakurai, K.; Satoh, H.; Nakai, K. Psychomotor ability in children prenatally exposed to methylmercury: The 18-month follow-up of tohoku study of child development. Tohoku J. Exp. Med. 2017, 242, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Klaassen, C.D.; Liu, J.; Choudhuri, S. Metallothionein: An intracellular protein to protect against cadmium toxicity. Annu. Rev. Pharmacol. Toxicol. 1999, 39, 267–294. [Google Scholar] [CrossRef] [PubMed]
- Vašák, M.; Meloni, G. Chemistry and biology of mammalian metallothioneins. J. Biol. Inorg. Chem. 2011, 16, 1067–1078. [Google Scholar] [CrossRef] [PubMed]
- Uchida, Y.; Takio, K.; Titani, K.; Ihara, Y.; Tomonaga, M. The growth inhibitory factor that is deficient in the Alzheimer’s disease brain is a 68 amino acid metallothionein-like protein. Neuron 1991, 7, 337–347. [Google Scholar] [CrossRef]
- Erickson, J.C.; Hollopeter, G.; Thomas, S.A.; Froelick, G.J.; Palmiter, R.D. Disruption of the metallothionein-III gene in mice: Analysis of brain zinc, behavior, and neuron vulnerability to metals, aging, and seizures. J. Neurosci. 1997, 17, 1271–1281. [Google Scholar] [CrossRef] [PubMed]
- West, A.K.; Hidalgo, J.; Eddins, D.; Levin, E.D.; Aschner, M. Metallothionein in the central nervous system: Roles in protection, regeneration and cognition. Neurotoxicology 2008, 29, 489–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, G.W.; Lee, J.Y.; Ryoke, K.; Matsuyama, F.; Kim, J.M.; Takahashi, T.; Naganuma, A. Gene expression profiling using DNA microarray analysis of the cerebellum of mice treated with methylmercury. J. Toxicol. Sci. 2011, 36, 389–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.Y.; Hwang, G.W.; Kim, M.S.; Takahashi, T.; Naganuma, A. Methylmercury induces a brain-specific increase in chemokine CCL4 expression in mice. J. Toxicol. Sci. 2012, 37, 1279–1282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zlotnik, A.; Yoshie, O. Chemokines: A new classification system and their role in immunity. Immunity 2000, 12, 121–127. [Google Scholar] [CrossRef]
- Gorter, J.A.; van Vliet, E.A.; Aronica, E.; Breit, T.; Rauwerda, H.; Lopes da Silva, F.H.; Wadman, W.J. Potential new antiepileptogenic targets indicated by microarray analysis in a rat model for temporal lobe epilepsy. J. Neurosci. 2006, 26, 11083–11110. [Google Scholar] [CrossRef] [PubMed]
- Soria, G.; Ben-Baruch, A. The inflammatory chemokines CCL2 and CCL5 in breast cancer. Cancer Lett. 2008, 267, 271–285. [Google Scholar] [CrossRef] [PubMed]
- Saiman, Y.; Friedman, S.L. The role of chemokines in acute liver injury. Front. Physiol. 2012, 3, 213. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, H.; Bolati, D.; Higashiyama, Y.; Nishijima, F.; Shimizu, K.; Niwa, T. Indoxyl sulfate upregulates renal expression of MCP-1 via production of ROS and activation of NF-kappaB, p53, ERK, and JNK in proximal tubular cells. Life Sci. 2012, 90, 525–530. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Takahashi, T.; Lee, J.Y.; Hwang, G.W.; Naganuma, A. Global chemokine expression in methylmercury-treated mice: Methylmercury induces brain-specific expression of CCL3 and CCL4. J. Toxicol. Sci. 2013, 38, 925–929. [Google Scholar] [CrossRef] [PubMed]
- Koumura, A.; Hamanaka, J.; Shimazawa, M.; Honda, A.; Tsuruma, K.; Uchida, Y.; Hozumi, I.; Satoh, M.; Inuzuka, T.; Hara, H. Metallothionein-III knockout mice aggravates the neuronal damage after transient focal cerebral ischemia. Brain Res. 2009, 1292, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Poulsen, C.B.; Penkowa, M.; Borup, R.; Nielsen, F.C.; Cáceres, M.; Quintana, A.; Molinero, A.; Carrasco, J.; Giralt, M.; Hidalgo, J. Brain response to traumatic brain injury in wild-type and interleukin-6 knockout mice: A microarray analysis. J. Neurochem. 2005, 92, 417–432. [Google Scholar] [CrossRef] [PubMed]
- Scuderi, C.; Stecca, C.; Iacomino, A.; Steardo, L. Role of astrocytes in major neurological disorders: The evidence and implications. IUBMB Life 2013, 65, 957–961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duerr, G.D.; Dewald, D.; Schmitz, E.J.; Verfuerth, L.; Keppel, K.; Peigney, C.; Ghanem, A.; Welz, A.; Dewald, O. Metallothioneins 1 and 2 modulate inflammation and support remodeling in ischemic cardiomyopathy in mice. Mediat. Inflamm. 2016, 2016, 7174127. [Google Scholar] [CrossRef] [PubMed]
- Gamo, K.; Kiryu-Seo, S.; Konishi, H.; Aoki, S.; Matsushima, K.; Wada, K.; Kiyama, H. G-protein-coupled receptor screen reveals a role for chemokine receptor CCR5 in suppressing microglial neurotoxicity. J. Neurosci. 2008, 28, 11980–11988. [Google Scholar] [CrossRef] [PubMed]
- Adler, M.W.; Rogers, T.J. Are chemokines the third major system in the brain? J. Leukoc. Biol. 2005, 78, 1204–1209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, R.J.; Rostene, W.; Apartis, E.; Banisadr, G.; Biber, K.; Milligan, E.D.; White, F.A.; Zhang, J. Chemokine action in the nervous system. J. Neurosci. 2008, 28, 11792–11795. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, M.; Lee, J.Y.; Satoh, M.; Watanabe, C. Neurobehavioral effects of postnatal exposure to low-level mercury vapor and/or methylmercury in mice. J. Toxicol. Sci. 2018, 43, 11–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, M.; Watanabe, C.; Horie, K.; Satoh, M.; Sawada, M.; Shimada, A. Neurobehavioral changes in metallothionein-null mice prenatally exposed to mercury vapor. Toxicol. Lett. 2005, 155, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, M.; Watanabe, C.; Kishimoto, M.; Yasutake, A.; Satoh, M.; Sawada, M.; Akama, Y. Behavioral changes in metallothionein-null mice after the cessation of long-term, low-level exposure to mercury vapor. Toxicol. Lett. 2006, 161, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Yao, C.P.; Allen, J.W.; Mutkus, L.A.; Xu, S.B.; Tan, K.H.; Aschner, M. Foreign metallothionein-I expression by transient transfection in MT-I and MT-II null astrocytes confers increased protection against acute methylmercury cytotoxicity. Brain Res. 2000, 855, 32–38. [Google Scholar] [CrossRef]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.-Y.; Tokumoto, M.; Hwang, G.-W.; Kim, M.-S.; Takahashi, T.; Naganuma, A.; Yoshida, M.; Satoh, M. Effect of Metallothionein-III on Mercury-Induced Chemokine Gene Expression. Toxics 2018, 6, 48. https://doi.org/10.3390/toxics6030048
Lee J-Y, Tokumoto M, Hwang G-W, Kim M-S, Takahashi T, Naganuma A, Yoshida M, Satoh M. Effect of Metallothionein-III on Mercury-Induced Chemokine Gene Expression. Toxics. 2018; 6(3):48. https://doi.org/10.3390/toxics6030048
Chicago/Turabian StyleLee, Jin-Yong, Maki Tokumoto, Gi-Wook Hwang, Min-Seok Kim, Tsutomu Takahashi, Akira Naganuma, Minoru Yoshida, and Masahiko Satoh. 2018. "Effect of Metallothionein-III on Mercury-Induced Chemokine Gene Expression" Toxics 6, no. 3: 48. https://doi.org/10.3390/toxics6030048
APA StyleLee, J. -Y., Tokumoto, M., Hwang, G. -W., Kim, M. -S., Takahashi, T., Naganuma, A., Yoshida, M., & Satoh, M. (2018). Effect of Metallothionein-III on Mercury-Induced Chemokine Gene Expression. Toxics, 6(3), 48. https://doi.org/10.3390/toxics6030048