A Neurodevelopmental Model of Combined Pyrethroid and Chronic Stress Exposure


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
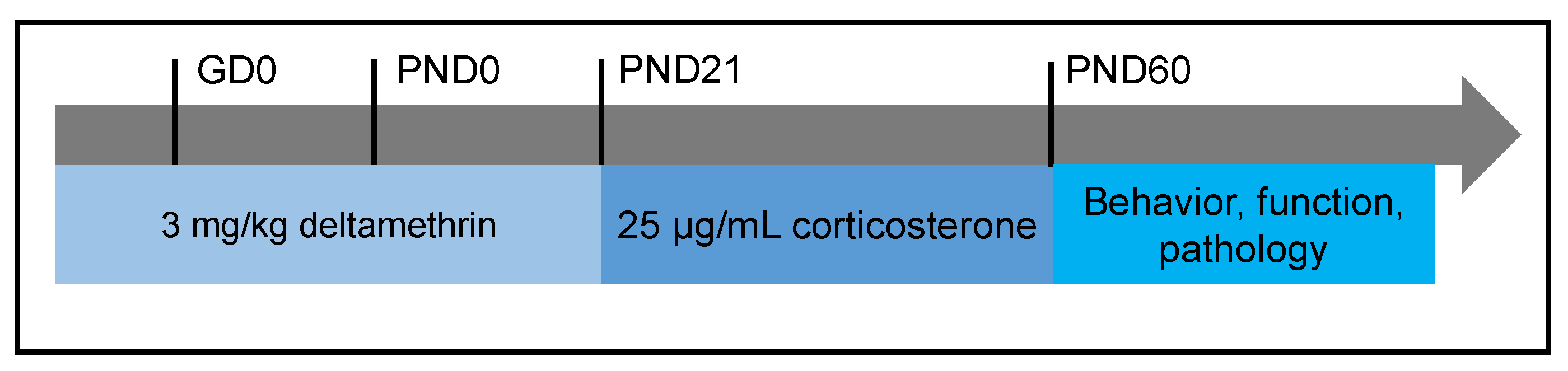
2.2. Exposure Paradigm
2.3. Behavioral Analyses
2.3.1. Locomotor Activity
2.3.2. Y Maze
2.3.3. Marble Burying
2.4. Serum Corticosterone Enzyme-Linked Immunsorbent Assay (ELISA)
2.5. mRNA Expression Analysis
2.6. Immunoblotting
2.7. Fast-Scan Cyclic Voltammetry
2.8. Statistical Analyses
3. Results
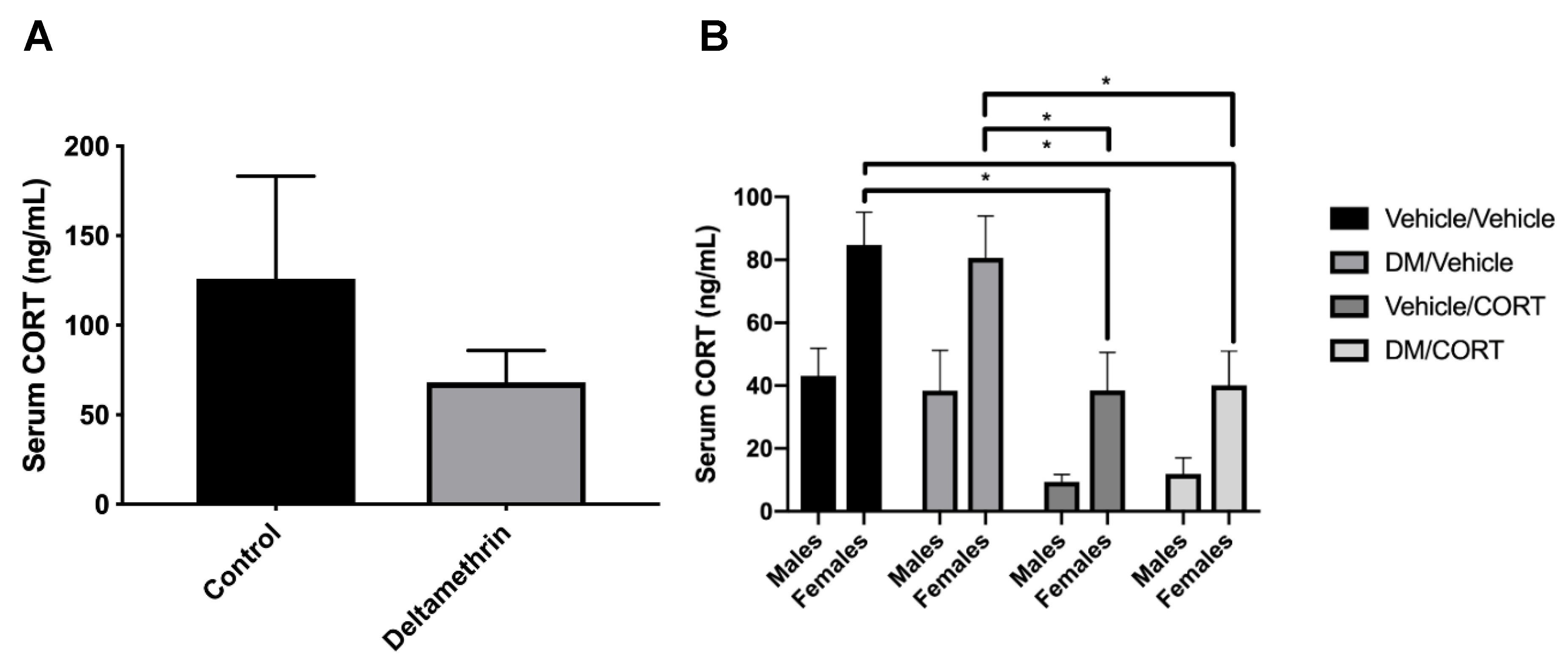
3.1. CORT Exposure Decreases Serum CORT Independent of Deltamethrin Exposure
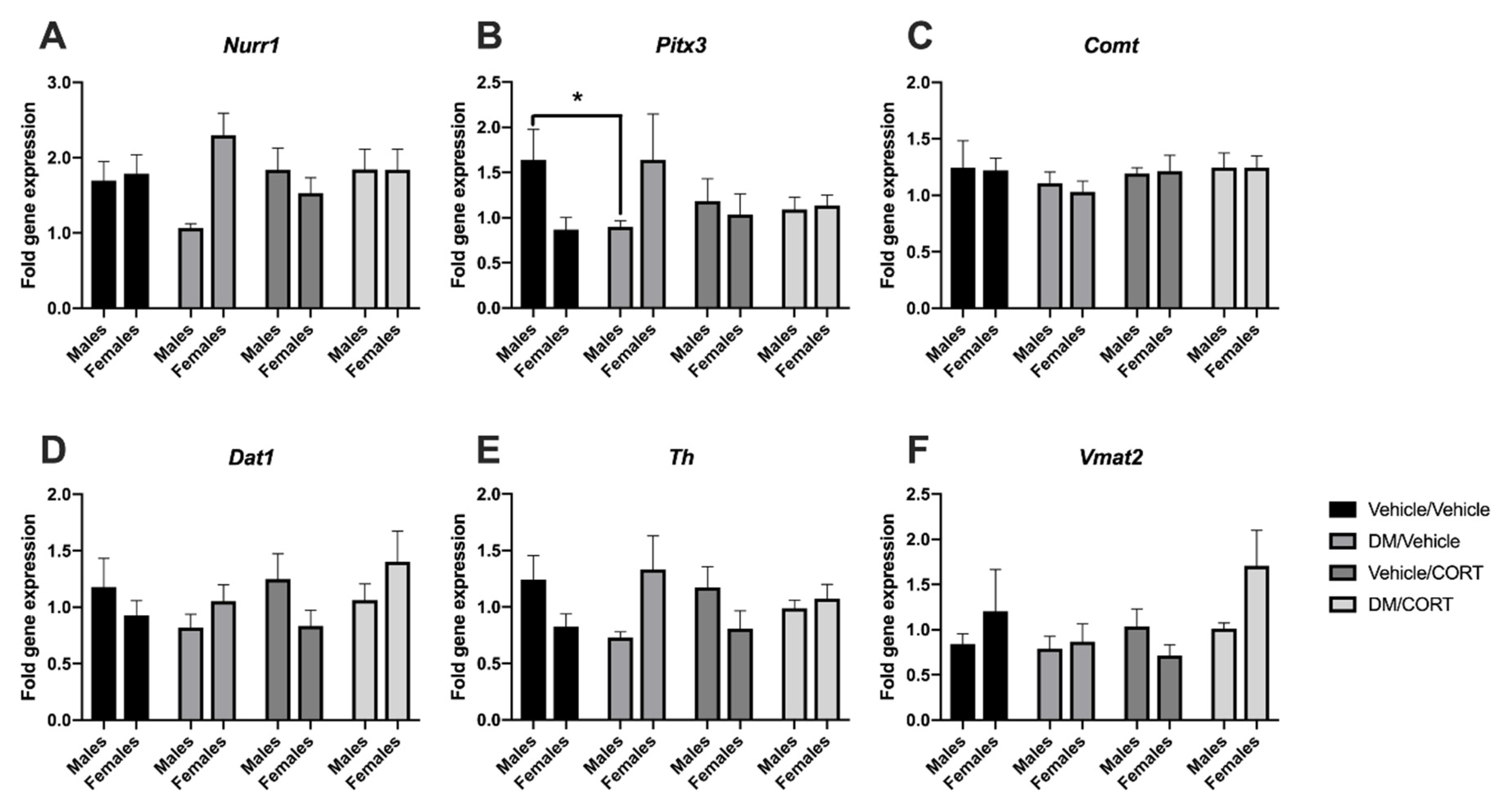
3.2. Sex-Specific Alterations in the Expression of Key Dopaminergic Genes
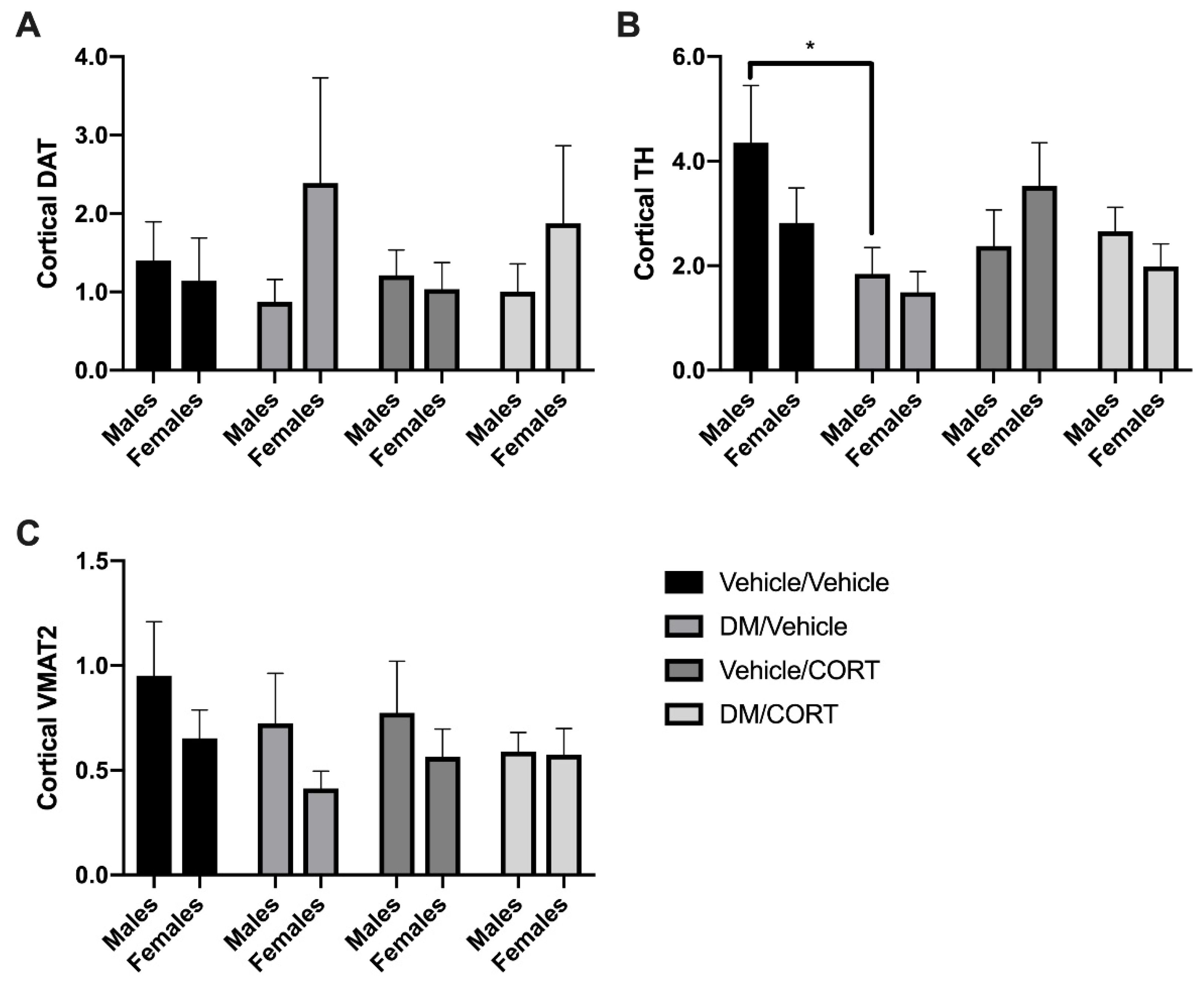
3.3. TH and VMAT2 Expression Were Significantly Decreased in Striatum of Females Exposed to Deltamethrin/CORT, and TH Expression Was Significantly Decreased in the Frontal Cortex of Males Exposed to Deltamethrin
3.4. Deltamethrin Exposure Significantly Slowed Striatal Dopamine Uptake Rate in Males
3.5. Males Exposed to Deltamethrin Display Increased Activity in Y Maze but Not Open Field
3.6. No Difference in Impulsivity as Measured Via Marble Burying in Animals Exposed to Deltamethrin and CORT
4. Discussion
4.1. Summary of Findings
4.2. Glucocorticoid Signaling and DM/CORT Exposure
4.3. Altered Midbrain RNA Expression
4.4. Altered Striatal and Cortical Protein Expression
4.5. Functional Consequences of Deltamethrin and CORT Exposure
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Thomas, R.; Sanders, S.; Doust, J.; Beller, E.; Glasziou, P. Prevalence of attention-deficit/hyperactivity disorder: A systematic review and meta-analysis. Pediatrics 2015, 135, e994–e1001. [Google Scholar] [CrossRef] [PubMed]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorder, 5th ed.; American Psychiatric Pub.: Arlington, VA, USA, 2013. [Google Scholar]
- Fayyad, J.; Sampson, N.A.; Hwang, I.; Adamowski, T.; Aguilar-Gaxiola, S.; Al-Hamzawi, A.; Andrade, L.H.; Borges, G.; de Girolamo, G.; Florescu, S.; et al. The descriptive epidemiology of DSM-IV Adult ADHD in the World Health Organization World Mental Health Surveys. Atten. Deficit Hyperact. Disord. 2017, 9, 47–65. [Google Scholar] [CrossRef]
- Gizer, I.R.; Ficks, C.; Waldman, I.D. Candidate gene studies of ADHD: A meta-analytic review. Hum. Genet. 2009, 126, 51–90. [Google Scholar] [CrossRef]
- Mortimer, N.; Ganster, T.; O’Leary, A.; Popp, S.; Freudenberg, F.; Reif, A.; Soler Artigas, M.; Ribases, M.; Ramos-Quiroga, J.A.; Lesch, K.P.; et al. Dissociation of impulsivity and aggression in mice deficient for the ADHD risk gene Adgrl3: Evidence for dopamine transporter dysregulation. Neuropharmacology 2019. [Google Scholar] [CrossRef]
- Grunblatt, E.; Werling, A.M.; Roth, A.; Romanos, M.; Walitza, S. Association study and a systematic meta-analysis of the VNTR polymorphism in the 3‘-UTR of dopamine transporter gene and attention-deficit hyperactivity disorder. J. Neural Transm. 2019, 126, 517–529. [Google Scholar] [CrossRef]
- Fraporti, T.T.; Contini, V.; Tovo-Rodrigues, L.; Recamonde-Mendoza, M.; Rovaris, D.L.; Rohde, L.A.; Hutz, M.H.; Salatino-Oliveira, A.; Genro, J.P. Synergistic effects between ADORA2A and DRD2 genes on anxiety disorders in children with ADHD. Prog. Neuropsychopharmacol. Biol. Psychiatry 2019. [Google Scholar] [CrossRef] [PubMed]
- Akutagava-Martins, G.C.; Salatino-Oliveira, A.; Kieling, C.; Genro, J.P.; Polanczyk, G.V.; Anselmi, L.; Menezes, A.M.; Goncalves, H.; Wehrmeister, F.C.; Barros, F.C.; et al. COMT and DAT1 genes are associated with hyperactivity and inattention traits in the 1993 Pelotas Birth Cohort: Evidence of sex-specific combined effect. J. Psychiatry Neurosci. Jpn. 2016, 41, 150270. [Google Scholar] [CrossRef] [PubMed]
- Maitra, S.; Sarkar, K.; Sinha, S.; Mukhopadhyay, K. The Dopamine Receptor D5 May Influence Age of Onset: An Exploratory Study on Indo-Caucasoid ADHD Subjects. J. Child Neurol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Van Mil, N.H.; Steegers-Theunissen, R.P.; Bouwland-Both, M.I.; Verbiest, M.M.; Rijlaarsdam, J.; Hofman, A.; Steegers, E.A.; Heijmans, B.T.; Jaddoe, V.W.; Verhulst, F.C.; et al. DNA methylation profiles at birth and child ADHD symptoms. J. Psychiatr. Res. 2014, 49, 51–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volkow, N.D.; Fowler, J.S.; Wang, G.; Ding, Y.; Gatley, S.J. Mechanism of action of methylphenidate: Insights from PET imaging studies. J. Atten. Disord. 2002, 6 (Suppl. 1), S31–S43. [Google Scholar] [CrossRef]
- Madras, B.K.; Miller, G.M.; Fischman, A.J. The dopamine transporter and attention-deficit/hyperactivity disorder. Biol. Psychiatry 2005, 57, 1397–1409. [Google Scholar] [CrossRef] [PubMed]
- Sakrikar, D.; Mazei-Robison, M.S.; Mergy, M.A.; Richtand, N.W.; Han, Q.; Hamilton, P.J.; Bowton, E.; Galli, A.; Veenstra-Vanderweele, J.; Gill, M.; et al. Attention deficit/hyperactivity disorder-derived coding variation in the dopamine transporter disrupts microdomain targeting and trafficking regulation. J. Neurosci. 2012, 32, 5385–5397. [Google Scholar] [CrossRef] [PubMed]
- Khadka, S.; Pearlson, G.D.; Calhoun, V.D.; Liu, J.; Gelernter, J.; Bessette, K.L.; Stevens, M.C. Multivariate Imaging Genetics Study of MRI Gray Matter Volume and SNPs Reveals Biological Pathways Correlated with Brain Structural Differences in Attention Deficit Hyperactivity Disorder. Front. Psychiatry 2016, 7, 128. [Google Scholar] [CrossRef]
- Fernandez-Jaen, A.; Albert, J.; Fernandez-Mayoralas, D.M.; Lopez-Martin, S.; Fernandez-Perrone, A.L.; Jimenez de la Pena, M.; Calleja-Perez, B.; Recio Rodriguez, M.; Lopez Arribas, S. Cingulate Cortical Thickness and Dopamine Transporter (DAT1) Genotype in Children and Adolescents With ADHD. J. Atten. Disord. 2016. [Google Scholar] [CrossRef]
- Gainetdinov, R.R.; Jones, S.R.; Caron, M.G. Functional hyperdopaminergia in dopamine transporter knock-out mice. Biol. Psychiatry 1999, 46, 303–311. [Google Scholar] [CrossRef]
- Salahpour, A.; Ramsey, A.J.; Medvedev, I.O.; Kile, B.; Sotnikova, T.D.; Holmstrand, E.; Ghisi, V.; Nicholls, P.J.; Wong, L.; Murphy, K.; et al. Increased amphetamine-induced hyperactivity and reward in mice overexpressing the dopamine transporter. Proc. Natl. Acad. Sci. USA 2008, 105, 4405–4410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leo, D.; Gainetdinov, R.R. Transgenic mouse models for ADHD. Cell Tissue Res. 2013, 354, 259–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gainetdinov, R.R.; Caron, M.G. An animal model of attention deficit hyperactivity disorder. Mol. Med. Today 2000, 6, 43–44. [Google Scholar] [CrossRef]
- Efimova, E.V.; Gainetdinov, R.R.; Budygin, E.A.; Sotnikova, T.D. Dopamine transporter mutant animals: A translational perspective. J. Neurogenet. 2016, 30, 5–15. [Google Scholar] [CrossRef]
- Kasparbauer, A.M.; Rujescu, D.; Riedel, M.; Pogarell, O.; Costa, A.; Meindl, T.; la Fougere, C.; Ettinger, U. Methylphenidate effects on brain activity as a function of SLC6A3 genotype and striatal dopamine transporter availability. Neuropsychopharmacology 2015, 40, 736–745. [Google Scholar] [CrossRef]
- Axelrod, J.; Weinshilboum, R. Catecholamines. N. Engl. J. Med. 1972, 287, 237–242. [Google Scholar] [CrossRef]
- Mulvihill, K.G. Presynaptic regulation of dopamine release: Role of the DAT and VMAT2 transporters. Neurochem. Int. 2019, 122, 94–105. [Google Scholar] [CrossRef]
- Klein, M.O.; Battagello, D.S.; Cardoso, A.R.; Hauser, D.N.; Bittencourt, J.C.; Correa, R.G. Dopamine: Functions, Signaling, and Association with Neurological Diseases. Cell. Mol. Neurobiol. 2019, 39, 31–59. [Google Scholar] [CrossRef]
- Purves, D. Neuroscience; Sinauer Associates: Sunderland, MA, USA, 2012. [Google Scholar]
- Nestler, E.J.; Hyman, S.E.; Malenka, R.C. Molecular Neuropharmacology: A Foundation for Clinical Neuroscience, 2nd ed.; McGraw-Hill Education: New York, NY, USA, 2008. [Google Scholar]
- Purper-Ouakil, D.; Ramoz, N.; Lepagnol-Bestel, A.-M.; Gorwood, P.; Simonneau, M. Neurobiology of Attention Deficit/Hyperactivity Disorder. Pediatr. Res. 2011, 69, 69R–76R. [Google Scholar] [CrossRef] [Green Version]
- Thapar, A.; Cooper, M.; Eyre, O.; Langley, K. What have we learnt about the causes of ADHD? J. Child Psychol. Psychiatry Allied Discip. 2013, 54, 3–16. [Google Scholar] [CrossRef]
- Neuman, R.J.; Lobos, E.; Reich, W.; Henderson, C.A.; Sun, L.W.; Todd, R.D. Prenatal smoking exposure and dopaminergic genotypes interact to cause a severe ADHD subtype. Biol. Psychiatry 2007, 61, 1320–1328. [Google Scholar] [CrossRef]
- Russell, A.E.; Ford, T.; Williams, R.; Russell, G. The Association Between Socioeconomic Disadvantage and Attention Deficit/Hyperactivity Disorder (ADHD): A Systematic Review. Child Psychiatry Hum. Dev. 2016, 47, 440–458. [Google Scholar] [CrossRef]
- Russell, A.E.; Ford, T.; Russell, G. Socioeconomic Associations with ADHD: Findings from a Mediation Analysis. PLoS ONE 2015, 10, e0128248. [Google Scholar] [CrossRef]
- Van der Meer, D.; Hoekstra, P.J.; van Donkelaar, M.; Bralten, J.; Oosterlaan, J.; Heslenfeld, D.; Faraone, S.V.; Franke, B.; Buitelaar, J.K.; Hartman, C.A. Predicting attention-deficit/hyperactivity disorder severity from psychosocial stress and stress-response genes: A random forest regression approach. Transl. Psychiatry 2017, 7, e1145. [Google Scholar] [CrossRef]
- Class, Q.A.; Abel, K.M.; Khashan, A.S.; Rickert, M.E.; Dalman, C.; Larsson, H.; Hultman, C.M.; Langstrom, N.; Lichtenstein, P.; D’Onofrio, B.M. Offspring psychopathology following preconception, prenatal and postnatal maternal bereavement stress. Psychol. Med. 2014, 44, 71–84. [Google Scholar] [CrossRef]
- Pessah, I.N.; Lein, P.J.; Seegal, R.F.; Sagiv, S.K. Neurotoxicity of polychlorinated biphenyls and related organohalogens. Acta Neuropathol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.H.; Yu, C.J.; Du, J.C.; Chiou, H.C.; Chen, H.C.; Yang, W.; Chung, M.Y.; Chen, Y.S.; Hwang, B.; Mao, I.F.; et al. The interactions among organophosphate pesticide exposure, oxidative stress, and genetic polymorphisms of dopamine receptor D4 increase the risk of attention deficit/hyperactivity disorder in children. Environ. Res. 2018, 160, 339–346. [Google Scholar] [CrossRef]
- Shen, H.; Main, K.M.; Virtanen, H.E.; Damggard, I.N.; Haavisto, A.M.; Kaleva, M.; Boisen, K.A.; Schmidt, I.M.; Chellakooty, M.; Skakkebaek, N.E.; et al. From mother to child: Investigation of prenatal and postnatal exposure to persistent bioaccumulating toxicants using breast milk and placenta biomonitoring. Chemosphere 2007, 67, S256–S262. [Google Scholar] [CrossRef] [PubMed]
- Landrigan, P.J.; Rauh, V.A.; Galvez, M.P. Environmental justice and the health of children. Mt. Sinai J. Med. N. Y. 2010, 77, 178–187. [Google Scholar] [CrossRef] [PubMed]
- Landrigan, P.J.; Goldman, L.R. Children’s vulnerability to toxic chemicals: A challenge and opportunity to strengthen health and environmental policy. Health Aff. (Proj. Hope) 2011, 30, 842–850. [Google Scholar] [CrossRef]
- Grandjean, P.; Landrigan, P.J. Neurobehavioural effects of developmental toxicity. Lancet. Neurol. 2014, 13, 330–338. [Google Scholar] [CrossRef] [Green Version]
- Pastor, P.N.; Reuben, C.A.; Duran, C.; Howie, L.D. Association between Diagnosed ADHD and Selected Characteristics among Children Aged 4–17 Years: United States, 2011–2013; NCHS Data Brief, No 201; National Center for Health Statistics: Hyattsville, MD, USA, 2015.
- Richardson, J.R.; Taylor, M.M.; Shalat, S.L.; Guillot, T.S., 3rd; Caudle, W.M.; Hossain, M.M.; Mathews, T.A.; Jones, S.R.; Cory-Slechta, D.A.; Miller, G.W. Developmental pesticide exposure reproduces features of attention deficit hyperactivity disorder. FASEB J. 2015. [Google Scholar] [CrossRef]
- Quiros-Alcala, L.; Mehta, S.; Eskenazi, B. Pyrethroid pesticide exposure and parental report of learning disability and attention deficit/hyperactivity disorder in U.S. children: NHANES 1999–2002. Environ. Health Perspect. 2014, 122, 1336–1342. [Google Scholar] [CrossRef]
- Larsson, H.; Sariaslan, A.; Langstrom, N.; D’Onofrio, B.; Lichtenstein, P. Family income in early childhood and subsequent attention deficit/hyperactivity disorder: A quasi-experimental study. J. Child Psychol. Psychiatry Allied Discip. 2014, 55, 428–435. [Google Scholar] [CrossRef]
- Bock, J.; Breuer, S.; Poeggel, G.; Braun, K. Early life stress induces attention-deficit hyperactivity disorder (ADHD)-like behavioral and brain metabolic dysfunctions: Functional imaging of methylphenidate treatment in a novel rodent model. Brain Struct. Funct. 2016. [Google Scholar] [CrossRef]
- Klaassen, C.D. Casarett and Doull’s Toxicology: The Basic Science of Poisons; McGraw-Hill: New York, NY, USA, 2013; Volume 1236. [Google Scholar]
- Burr, S.A.; Ray, D.E. Structure-activity and interaction effects of 14 different pyrethroids on voltage-gated chloride ion channels. Toxicol. Sci. 2004, 77, 341–346. [Google Scholar] [CrossRef]
- Bloomquist, J.R.; Barlow, R.L.; Gillette, J.S.; Li, W.; Kirby, M.L. Selective effects of insecticides on nigrostriatal dopaminergic nerve pathways. Neurotoxicology 2002, 23, 537–544. [Google Scholar] [CrossRef]
- Gillette, J.S.; Bloomquist, J.R. Differential up-regulation of striatal dopamine transporter and alpha-synuclein by the pyrethroid insecticide permethrin. Toxicol. Appl. Pharm. 2003, 192, 287–293. [Google Scholar] [CrossRef]
- Elwan, M.A.; Richardson, J.R.; Guillot, T.S.; Caudle, W.M.; Miller, G.W. Pyrethroid pesticide-induced alterations in dopamine transporter function. Toxicol. Appl. Pharm. 2006, 211, 188–197. [Google Scholar] [CrossRef]
- Mubarak Hossain, M.; Suzuki, T.; Sato, N.; Sato, I.; Takewaki, T.; Suzuki, K.; Tachikawa, E.; Kobayashi, H. Differential effects of pyrethroid insecticides on extracellular dopamine in the striatum of freely moving rats. Toxicol. Appl. Pharm. 2006, 217, 25–34. [Google Scholar] [CrossRef]
- Wagner-Schuman, M.; Richardson, J.R.; Auinger, P.; Braun, J.M.; Lanphear, B.P.; Epstein, J.N.; Yolton, K.; Froehlich, T.E. Association of pyrethroid pesticide exposure with attention-deficit/hyperactivity disorder in a nationally representative sample of U.S. children. Environ. Health Glob. Access Sci. Source 2015, 14, 44. [Google Scholar] [CrossRef]
- Russell, G.; Ford, T.; Rosenberg, R.; Kelly, S. The association of attention deficit hyperactivity disorder with socioeconomic disadvantage: Alternative explanations and evidence. J. Child Psychol. Psychiatry Allied Discip. 2014, 55, 436–445. [Google Scholar] [CrossRef]
- Steptoe, A.; Feldman, P.J. Neighborhood problems as sources of chronic stress: Development of a measure of neighborhood problems, and associations with socioeconomic status and health. Ann. Behav. Med. Publ. Soc. Behav. Med. 2001, 23, 177–185. [Google Scholar] [CrossRef]
- Baum, A.; Garofalo, J.P.; Yali, A.M. Socioeconomic status and chronic stress. Does stress account for SES effects on health? Ann. N. Y. Acad. Sci. 1999, 896, 131–144. [Google Scholar] [CrossRef]
- McEwen, B.S.; Gianaros, P.J. Central role of the brain in stress and adaptation: Links to socioeconomic status, health, and disease. Ann. N. Y. Acad. Sci. 2010, 1186, 190–222. [Google Scholar] [CrossRef] [Green Version]
- McEwen, B.S.; Tucker, P. Critical biological pathways for chronic psychosocial stress and research opportunities to advance the consideration of stress in chemical risk assessment. Am. J. Public Health 2011, 101 (Suppl. 1), S131–S139. [Google Scholar] [CrossRef]
- Vaghri, Z.; Guhn, M.; Weinberg, J.; Grunau, R.E.; Yu, W.; Hertzman, C. Hair cortisol reflects socio-economic factors and hair zinc in preschoolers. Psychoneuroendocrinology 2013, 38, 331–340. [Google Scholar] [CrossRef]
- Vliegenthart, J.; Noppe, G.; van Rossum, E.F.; Koper, J.W.; Raat, H.; van den Akker, E.L. Socioeconomic status in children is associated with hair cortisol levels as a biological measure of chronic stress. Psychoneuroendocrinology 2016, 65, 9–14. [Google Scholar] [CrossRef]
- Butts, K.A.; Phillips, A.G. Glucocorticoid receptors in the prefrontal cortex regulate dopamine efflux to stress via descending glutamatergic feedback to the ventral tegmental area. Int. J. Neuropsychopharmacol. 2013, 16, 1799–1807. [Google Scholar] [CrossRef] [Green Version]
- Lucas, L.R.; Celen, Z.; Tamashiro, K.L.; Blanchard, R.J.; Blanchard, D.C.; Markham, C.; Sakai, R.R.; McEwen, B.S. Repeated exposure to social stress has long-term effects on indirect markers of dopaminergic activity in brain regions associated with motivated behavior. Neuroscience 2004, 124, 449–457. [Google Scholar] [CrossRef]
- Hensleigh, E.; Pritchard, L.M. Glucocorticoid receptor expression and sub-cellular localization in dopamine neurons of the rat midbrain. Neurosci. Lett. 2013, 556, 191–195. [Google Scholar] [CrossRef]
- Brown, P. Race, Class, and Environmental Health: A Review and Systematization of the Literature. Environ. Res. 1995, 69, 15–30. [Google Scholar] [CrossRef]
- Lu, C.; Adamkiewicz, G.; Attfield, K.R.; Kapp, M.; Spengler, J.D.; Tao, L.; Xie, S.H. Household pesticide contamination from indoor pest control applications in urban low-income public housing dwellings: A community-based participatory research. Environ. Sci. Technol. 2013, 47, 2018–2025. [Google Scholar] [CrossRef]
- Julien, R.; Adamkiewicz, G.; Levy, J.I.; Bennett, D.; Nishioka, M.; Spengler, J.D. Pesticide loadings of select organophosphate and pyrethroid pesticides in urban public housing. J. Expo. Sci. Environ. Epidemiol. 2008, 18, 167–174. [Google Scholar] [CrossRef]
- Wason, S.C.; Julien, R.; Perry, M.J.; Smith, T.J.; Levy, J.I. Modeling exposures to organophosphates and pyrethroids for children living in an urban low-income environment. Environ. Res. 2013, 124, 13–22. [Google Scholar] [CrossRef]
- Castorina, R.; Bradman, A.; Fenster, L.; Barr, D.B.; Bravo, R.; Vedar, M.G.; Harnly, M.E.; McKone, T.E.; Eisen, E.A.; Eskenazi, B. Comparison of current-use pesticide and other toxicant urinary metabolite levels among pregnant women in the CHAMACOS cohort and NHANES. Environ. Health Perspect. 2010, 118, 856–863. [Google Scholar] [CrossRef]
- Berkowitz, G.S.; Obel, J.; Deych, E.; Lapinski, R.; Godbold, J.; Liu, Z.; Landrigan, P.J.; Wolff, M.S. Exposure to indoor pesticides during pregnancy in a multiethnic, urban cohort. Environ. Health Perspect. 2003, 111, 79–84. [Google Scholar] [CrossRef]
- Barr, D.B.; Olsson, A.O.; Wong, L.Y.; Udunka, S.; Baker, S.E.; Whitehead, R.D.; Magsumbol, M.S.; Williams, B.L.; Needham, L.L. Urinary concentrations of metabolites of pyrethroid insecticides in the general U.S. population: National Health and Nutrition Examination Survey 1999–2002. Environ. Health Perspect. 2010, 118, 742–748. [Google Scholar] [CrossRef]
- Panuwet, P.; Prapamontol, T.; Chantara, S.; Barr, D.B. Urinary pesticide metabolites in school students from northern Thailand. Int. J. Hyg. Environ. Health 2009, 212, 288–297. [Google Scholar] [CrossRef]
- Lu, C.; Barr, D.B.; Pearson, M.A.; Walker, L.A.; Bravo, R. The attribution of urban and suburban children’s exposure to synthetic pyrethroid insecticides: A longitudinal assessment. J. Expo. Sci. Environ. Epidemiol. 2009, 19, 69–78. [Google Scholar] [CrossRef]
- Perez, J.J.; Williams, M.K.; Weerasekera, G.; Smith, K.; Whyatt, R.M.; Needham, L.L.; Barr, D.B. Measurement of pyrethroid, organophosphorus, and carbamate insecticides in human plasma using isotope dilution gas chromatography-high resolution mass spectrometry. J. Chromatogr. BAnal. Technol. Biomed. Life Sci. 2010, 878, 2554–2562. [Google Scholar] [CrossRef]
- Lu, D.; Wang, D.; Feng, C.; Jin, Y.; Zhou, Z.; Wu, C.; Lin, Y.; Wang, G. Urinary concentrations of metabolites of pyrethroid insecticides in textile workers, Eastern China. Environ. Int. 2013, 60, 137–144. [Google Scholar] [CrossRef]
- Schettgen, T.; Heudorf, U.; Drexler, H.; Angerer, J. Pyrethroid exposure of the general population-is this due to diet. Toxicol. Lett. 2002, 134, 141–145. [Google Scholar] [CrossRef]
- Morgan, M.K. Children’s exposures to pyrethroid insecticides at home: A review of data collected in published exposure measurement studies conducted in the United States. Int. J. Environ. Res. Public Health 2012, 9, 2964–2985. [Google Scholar] [CrossRef]
- Gong, S.; Miao, Y.-L.; Jiao, G.-Z.; Sun, M.-J.; Li, H.; Lin, J.; Luo, M.-J.; Tan, J.-H. Dynamics and Correlation of Serum Cortisol and Corticosterone under Different Physiological or Stressful Conditions in Mice. PLoS ONE 2015, 10, e0117503. [Google Scholar] [CrossRef]
- Yin, X.; Guven, N.; Dietis, N. Stress-based animal models of depression: Do we actually know what we are doing? Brain Res. 2016, 1652, 30–42. [Google Scholar] [CrossRef]
- Gourley, S.L.; Taylor, J.R. Recapitulation and reversal of a persistent depression-like syndrome in rodents. Curr. Protoc. Neurosci. 2009. [Google Scholar] [CrossRef]
- Caudle, W.M.; Richardson, J.R.; Wang, M.Z.; Taylor, T.N.; Guillot, T.S.; McCormack, A.L.; Colebrooke, R.E.; Di Monte, D.A.; Emson, P.C.; Miller, G.W. Reduced vesicular storage of dopamine causes progressive nigrostriatal neurodegeneration. J. Neurosci. 2007, 27, 8138–8148. [Google Scholar] [CrossRef]
- Cliburn, R.A.; Dunn, A.R.; Stout, K.A.; Hoffman, C.A.; Lohr, K.M.; Bernstein, A.I.; Winokur, E.J.; Burkett, J.; Schmitz, Y.; Caudle, W.M.; et al. Immunochemical localization of vesicular monoamine transporter 2 (VMAT2) in mouse brain. J. Chem. Neuroanat. 2017, 83–84, 82–90. [Google Scholar] [CrossRef]
- Dunn, A.R.; Stout, K.A.; Ozawa, M.; Lohr, K.M.; Hoffman, C.A.; Bernstein, A.I.; Li, Y.; Wang, M.; Sgobio, C.; Sastry, N.; et al. Synaptic vesicle glycoprotein 2C (SV2C) modulates dopamine release and is disrupted in Parkinson disease. Proc. Natl. Acad. Sci. USA 2017, 114, E2253–E2262. [Google Scholar] [CrossRef]
- Lohr, K.M.; Bernstein, A.I.; Stout, K.A.; Dunn, A.R.; Lazo, C.R.; Alter, S.P.; Wang, M.; Li, Y.; Fan, X.; Hess, E.J.; et al. Increased vesicular monoamine transporter enhances dopamine release and opposes Parkinson disease-related neurodegeneration in vivo. Proc. Natl. Acad. Sci. USA 2014, 111, 9977–9982. [Google Scholar] [CrossRef] [Green Version]
- Yorgason, J.T.; Espana, R.A.; Jones, S.R. Demon voltammetry and analysis software: Analysis of cocaine-induced alterations in dopamine signaling using multiple kinetic measures. J. Neurosci. Methods 2011, 202, 158–164. [Google Scholar] [CrossRef] [Green Version]
- Caudle, W.M.; Richardson, J.R.; Wang, M.; Miller, G.W. Perinatal heptachlor exposure increases expression of presynaptic dopaminergic markers in mouse striatum. Neurotoxicology 2005, 26, 721–728. [Google Scholar] [CrossRef]
- Richardson, J.R.; Caudle, W.M.; Wang, M.; Dean, E.D.; Pennell, K.D.; Miller, G.W. Developmental exposure to the pesticide dieldrin alters the dopamine system and increases neurotoxicity in an animal model of Parkinson’s disease. FASEB J. 2006, 20, 1695–1697. [Google Scholar] [CrossRef]
- Wilson, W.W.; Shapiro, L.P.; Bradner, J.M.; Caudle, W.M. Developmental exposure to the organochlorine insecticide endosulfan damages the nigrostriatal dopamine system in male offspring. Neurotoxicology 2014, 44, 279–287. [Google Scholar] [CrossRef] [Green Version]
- Richardson, J.R.; Caudle, W.M.; Wang, M.Z.; Dean, E.D.; Pennell, K.D.; Miller, G.W. Developmental heptachlor exposure increases susceptibility of dopamine neurons to N-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)in a gender-specific manner. Neurotoxicology 2008, 29, 855–863. [Google Scholar] [CrossRef] [PubMed]
- Isovich, E.; Mijnster, M.J.; Flugge, G.; Fuchs, E. Chronic psychosocial stress reduces the density of dopamine transporters. Eur. J. Neurosci. 2000, 12, 1071–1078. [Google Scholar] [CrossRef]
- Graf, E.N.; Wheeler, R.A.; Baker, D.A.; Ebben, A.L.; Hill, J.E.; McReynolds, J.R.; Robble, M.A.; Vranjkovic, O.; Wheeler, D.S.; Mantsch, J.R.; et al. Corticosterone acts in the nucleus accumbens to enhance dopamine signaling and potentiate reinstatement of cocaine seeking. J. Neurosci. 2013, 33, 11800–11810. [Google Scholar] [CrossRef]
- Stout, K.A.; Dunn, A.R.; Lohr, K.M.; Alter, S.P.; Cliburn, R.A.; Guillot, T.S.; Miller, G.W. Selective Enhancement of Dopamine Release in the Ventral Pallidum of Methamphetamine-Sensitized Mice. ACS Chem. Neurosci. 2016. [Google Scholar] [CrossRef] [PubMed]
- Wallis, D.; Hill, D.S.; Mendez, I.A.; Abbott, L.C.; Finnell, R.H.; Wellman, P.J.; Setlow, B. Initial characterization of mice null for Lphn3, a gene implicated in ADHD and addiction. Brain Res. 2012, 1463, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Kim, P.; Choi, C.S.; Park, J.H.; Joo, S.H.; Kim, S.Y.; Ko, H.M.; Kim, K.C.; Jeon, S.J.; Park, S.H.; Han, S.H.; et al. Chronic exposure to ethanol of male mice before mating produces attention deficit hyperactivity disorder-like phenotype along with epigenetic dysregulation of dopamine transporter expression in mouse offspring. J. Neurosci. Res. 2014, 92, 658–670. [Google Scholar] [CrossRef] [PubMed]
- Majdak, P.; Ossyra, J.R.; Ossyra, J.M.; Cobert, A.J.; Hofmann, G.C.; Tse, S.; Panozzo, B.; Grogan, E.L.; Sorokina, A.; Rhodes, J.S. A new mouse model of ADHD for medication development. Sci. Rep. 2016, 6, 39472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Itohara, S.; Kobayashi, Y.; Nakashiba, T. Genetic factors underlying attention and impulsivity: Mouse models of attention-deficit/hyperactivity disorder. Curr. Opin. Behav. Sci. 2015, 2, 46–51. [Google Scholar] [CrossRef]
- Barros, V.G.; Boado, L.A.; Adamo, A.M.; Caviedes, R.; Caviedes, P.; Antonelli, M.C. Corticosterone down-regulates dopamine D4 receptor in a mouse cerebral cortex neuronal cell line. Neurotox. Res. 2003, 5, 369–373. [Google Scholar] [CrossRef]
- Hu, L.; Yang, J.; Song, T.; Hou, N.; Liu, Y.; Zhao, X.; Zhang, D.; Wang, L.; Wang, T.; Huang, C. A new stress model, a scream sound, alters learning and monoamine levels in rat brain. Physiol. Behav. 2014, 123, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Mora, F.; Segovia, G.; Del Arco, A.; de Blas, M.; Garrido, P. Stress, neurotransmitters, corticosterone and body-brain integration. Brain Res. 2012, 1476, 71–85. [Google Scholar] [CrossRef] [PubMed]
- Gourley, S.L.; Kedves, A.T.; Olausson, P.; Taylor, J.R. A history of corticosterone exposure regulates fear extinction and cortical NR2B, GluR2/3, and BDNF. Neuropsychopharmacology 2009, 34, 707–716. [Google Scholar] [CrossRef] [PubMed]
- Mekiri, M.; Gardier, A.M.; David, D.J.; Guilloux, J.P. Chronic corticosterone administration effects on behavioral emotionality in female c57bl6 mice. Exp. Clin. Psychopharmacol. 2017, 25, 94–104. [Google Scholar] [CrossRef] [PubMed]
- Van Bodegom, M.; Homberg, J.R.; Henckens, M. Modulation of the Hypothalamic-Pituitary-Adrenal Axis by Early Life Stress Exposure. Front. Cell. Neurosci. 2017, 11, 87. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.J.; Liu, M.Y.; Li, H.; Liu, X.; Chen, C.; Han, Z.; Wu, H.Y.; Jing, X.; Zhou, H.H.; Suh, H.; et al. The different roles of glucocorticoids in the hippocampus and hypothalamus in chronic stress-induced HPA axis hyperactivity. PLoS ONE 2014, 9, e97689. [Google Scholar] [CrossRef] [PubMed]
- Herman, J.P.; McKlveen, J.M.; Ghosal, S.; Kopp, B.; Wulsin, A.; Makinson, R.; Scheimann, J.; Myers, B. Regulation of the Hypothalamic-Pituitary-Adrenocortical Stress Response. Compr. Physiol. 2016, 6, 603–621. [Google Scholar]
- Shahanoor, Z.; Sultana, R.; Baker, M.R.; Romeo, R.D. Neuroendocrine stress reactivity of male C57BL/6N mice following chronic oral corticosterone exposure during adulthood or adolescence. Psychoneuroendocrinology 2017, 86, 218–224. [Google Scholar] [CrossRef]
- Ulrich-Lai, Y.M.; Herman, J.P. Neural regulation of endocrine and autonomic stress responses. Nat. Rev. Neurosci. 2009, 10, 397–409. [Google Scholar] [CrossRef]
- Robinson, S.A.; Brookshire, B.R.; Lucki, I. Corticosterone exposure augments sensitivity to the behavioral and neuroplastic effects of fluoxetine in C57BL/6 mice. Neurobiol. Stress 2016, 3, 34–42. [Google Scholar] [CrossRef] [Green Version]
- Perlmann, T.; Wallen-Mackenzie, A. Nurr1, an orphan nuclear receptor with essential functions in developing dopamine cells. Cell Tissue Res. 2004, 318, 45–52. [Google Scholar] [CrossRef]
- Smidt, M.P.; Smits, S.M.; Burbach, J.P. Homeobox gene Pitx3 and its role in the development of dopamine neurons of the substantia nigra. Cell Tissue Res. 2004, 318, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Kadkhodaei, B.; Ito, T.; Joodmardi, E.; Mattsson, B.; Rouillard, C.; Carta, M.; Muramatsu, S.; Sumi-Ichinose, C.; Nomura, T.; Metzger, D.; et al. Nurr1 is required for maintenance of maturing and adult midbrain dopamine neurons. J. Neurosci. 2009, 29, 15923–15932. [Google Scholar] [CrossRef]
- Smidt, M.P.; Smits, S.M.; Bouwmeester, H.; Hamers, F.P.; van der Linden, A.J.; Hellemons, A.J.; Graw, J.; Burbach, J.P. Early developmental failure of substantia nigra dopamine neurons in mice lacking the homeodomain gene Pitx3. Development 2004, 131, 1145–1155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smidt, M.P.; van Schaick, H.S.; Lanctôt, C.; Tremblay, J.J.; Cox, J.J.; van der Kleij, A.A.; Wolterink, G.; Drouin, J.; Burbach, J.P. A homeodomain gene Ptx3 has highly restricted brain expression in mesencephalic dopaminergic neurons. Proc. Natl. Acad. Sci. USA 1997, 94, 13305–13310. [Google Scholar] [CrossRef] [Green Version]
- Smits, S.M.; Smidt, M.P. The role of Pitx3 in survival of midbrain dopaminergic neurons. J. Neural Transm. Suppl. 2006, 70, 57–60. [Google Scholar]
- Bissonette, G.B.; Roesch, M.R. Development and function of the midbrain dopamine system: What we know and what we need to. Genes Brain Behav. 2016, 15, 62–73. [Google Scholar] [CrossRef]
- Kas, M.J.; van der Linden, A.J.; Oppelaar, H.; von Oerthel, L.; Ramakers, G.M.; Smidt, M.P. Phenotypic segregation of aphakia and Pitx3-null mutants reveals that Pitx3 deficiency increases consolidation of specific movement components. Behav. Brain Res. 2008, 186, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, F.M.; van Erp, S.; van der Linden, A.J.; von Oerthel, L.; Burbach, J.P.; Smidt, M.P. Pitx3 potentiates Nurr1 in dopamine neuron terminal differentiation through release of SMRT-mediated repression. Development 2009, 136, 531–540. [Google Scholar] [CrossRef] [Green Version]
- Volpicelli, F.; De Gregorio, R.; Pulcrano, S.; Perrone-Capano, C.; di Porzio, U.; Bellenchi, G.C. Direct regulation of Pitx3 expression by Nurr1 in culture and in developing mouse midbrain. PLoS ONE 2012, 7, e30661. [Google Scholar] [CrossRef]
- Smidt, M.P.; Burbach, J.P. Terminal differentiation of mesodiencephalic dopaminergic neurons: The role of Nurr1 and Pitx3. Adv. Exp. Med. Biol. 2009, 651, 47–57. [Google Scholar]
- Luk, K.C.; Rymar, V.V.; van den Munckhof, P.; Nicolau, S.; Steriade, C.; Bifsha, P.; Drouin, J.; Sadikot, A.F. The transcription factor Pitx3 is expressed selectively in midbrain dopaminergic neurons susceptible to neurodegenerative stress. J. Neurochem. 2013, 125, 932–943. [Google Scholar] [CrossRef]
- SR, J.C.J. Fast Scan Cyclic Voltammetry of Dopamine and Serotonin in Mouse Brain Slices. In Electrochemical Methods for Neuroscience; Michael, A.C., Borland, L.M., Eds.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2007. [Google Scholar]
- Huang, E.Y.; Tsui, P.F.; Kuo, T.T.; Tsai, J.J.; Chou, Y.C.; Ma, H.I.; Chiang, Y.H.; Chen, Y.H. Amantadine ameliorates dopamine-releasing deficits and behavioral deficits in rats after fluid percussion injury. PLoS ONE 2014, 9, e86354. [Google Scholar] [CrossRef] [PubMed]
- Brodnik, Z.D.; Ferris, M.J.; Jones, S.R.; Espana, R.A. Reinforcing Doses of Intravenous Cocaine Produce Only Modest Dopamine Uptake Inhibition. ACS Chem. Neurosci. 2017, 8, 281–289. [Google Scholar] [CrossRef]
- Giros, B.; Jaber, M.; Jones, S.R.; Wightman, R.M.; Caron, M.G. Hyperlocomotion and indifference to cocaine and amphetamine in mice lacking the dopamine transporter. Nature 1996, 379, 606–612. [Google Scholar] [CrossRef] [PubMed]
- Rastedt, D.E.; Vaughan, R.A.; Foster, J.D. Palmitoylation mechanisms in dopamine transporter regulation. J. Chem. Neuroanat. 2017, 83–84, 3–9. [Google Scholar] [CrossRef]
- Lohr, K.M.; Masoud, S.T.; Salahpour, A.; Miller, G.W. Membrane transporters as mediators of synaptic dopamine dynamics: Implications for disease. Eur. J. Neurosci. 2017, 45, 20–33. [Google Scholar] [CrossRef]
- Loland, C.J.; Desai, R.I.; Zou, M.F.; Cao, J.; Grundt, P.; Gerstbrein, K.; Sitte, H.H.; Newman, A.H.; Katz, J.L.; Gether, U. Relationship between conformational changes in the dopamine transporter and cocaine-like subjective effects of uptake inhibitors. Mol Pharm. 2008, 73, 813–823. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, K.C.; Reith, M.E. The atypical stimulant and nootropic modafinil interacts with the dopamine transporter in a different manner than classical cocaine-like inhibitors. PLoS ONE 2011, 6, e25790. [Google Scholar] [CrossRef]
- Hamilton, P.J.; Campbell, N.G.; Sharma, S.; Erreger, K.; Herborg Hansen, F.; Saunders, C.; Belovich, A.N.; Sahai, M.A.; Cook, E.H.; Gether, U.; et al. De novo mutation in the dopamine transporter gene associates dopamine dysfunction with autism spectrum disorder. Mol. Psychiatry 2013, 18, 1315–1323. [Google Scholar] [CrossRef] [Green Version]
- Hansen, F.H.; Skjorringe, T.; Yasmeen, S.; Arends, N.V.; Sahai, M.A.; Erreger, K.; Andreassen, T.F.; Holy, M.; Hamilton, P.J.; Neergheen, V.; et al. Missense dopamine transporter mutations associate with adult parkinsonism and ADHD. J. Clin. Investig. 2014, 124, 3107–3120. [Google Scholar] [CrossRef] [Green Version]
- Kraft, A.D.; Aschner, M.; Cory-Slechta, D.A.; Bilbo, S.D.; Caudle, W.M.; Makris, S.L. Unmasking silent neurotoxicity following developmental exposure to environmental toxicants. Neurotoxicol. Teratol. 2016, 55, 38–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faraone, S.V.; Biederman, J.; Morley, C.P.; Spencer, T.J. Effect of stimulants on height and weight: A review of the literature. J. Am. Acad. Child Adolesc. Psychiatry 2008, 47, 994–1009. [Google Scholar] [CrossRef] [PubMed]
- Urban, K.R.; Gao, W.J. Methylphenidate and the juvenile brain: Enhancement of attention at the expense of cortical plasticity? Med. Hypotheses 2013, 81, 988–994. [Google Scholar] [CrossRef] [PubMed]
- Mannuzza, S.; Klein, R.G.; Truong, N.L.; Moulton, J.L., 3rd; Roizen, E.R.; Howell, K.H.; Castellanos, F.X. Age of methylphenidate treatment initiation in children with ADHD and later substance abuse: Prospective follow-up into adulthood. Am. J. Psychiatry 2008, 165, 604–609. [Google Scholar] [CrossRef] [PubMed]
- Klassen, A.F. Quality of life of children with attention deficit hyperactivity disorder. Expert Rev. Pharm. Outcomes Res. 2005, 5, 95–103. [Google Scholar] [CrossRef] [PubMed]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vester, A.I.; Chen, M.; Marsit, C.J.; Caudle, W.M. A Neurodevelopmental Model of Combined Pyrethroid and Chronic Stress Exposure. Toxics 2019, 7, 24. https://doi.org/10.3390/toxics7020024
Vester AI, Chen M, Marsit CJ, Caudle WM. A Neurodevelopmental Model of Combined Pyrethroid and Chronic Stress Exposure. Toxics. 2019; 7(2):24. https://doi.org/10.3390/toxics7020024
Chicago/Turabian StyleVester, Aimée I., Merry Chen, Carmen J. Marsit, and W. Michael Caudle. 2019. "A Neurodevelopmental Model of Combined Pyrethroid and Chronic Stress Exposure" Toxics 7, no. 2: 24. https://doi.org/10.3390/toxics7020024
APA StyleVester, A. I., Chen, M., Marsit, C. J., & Caudle, W. M. (2019). A Neurodevelopmental Model of Combined Pyrethroid and Chronic Stress Exposure. Toxics, 7(2), 24. https://doi.org/10.3390/toxics7020024