Lethal and Sublethal Effects of Pyriproxyfen on Apis and Non-Apis Bees

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Effects of Pyriproxyfen on Apis Bees
2.1. Mortality
2.2. Effects on Development
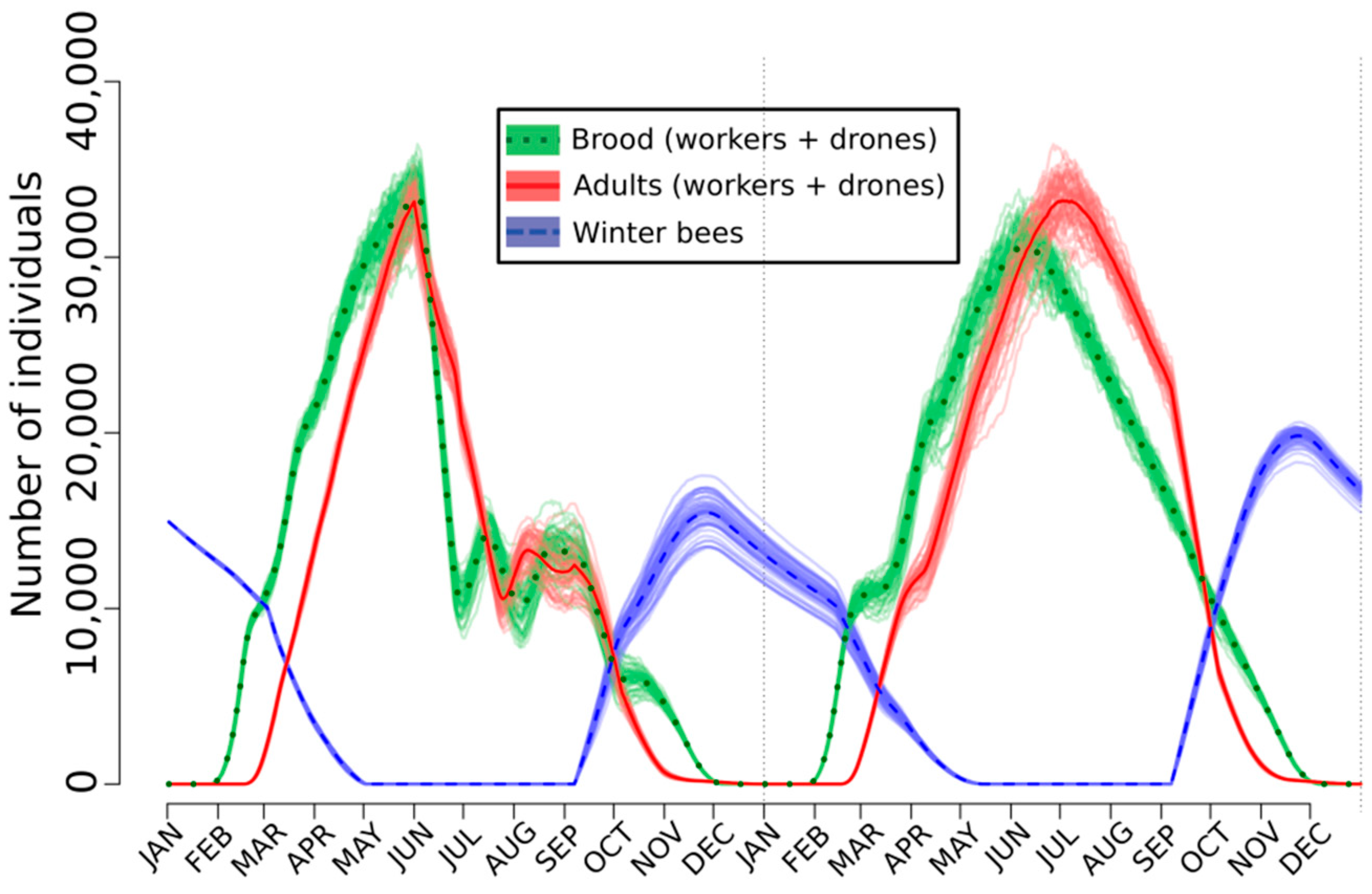
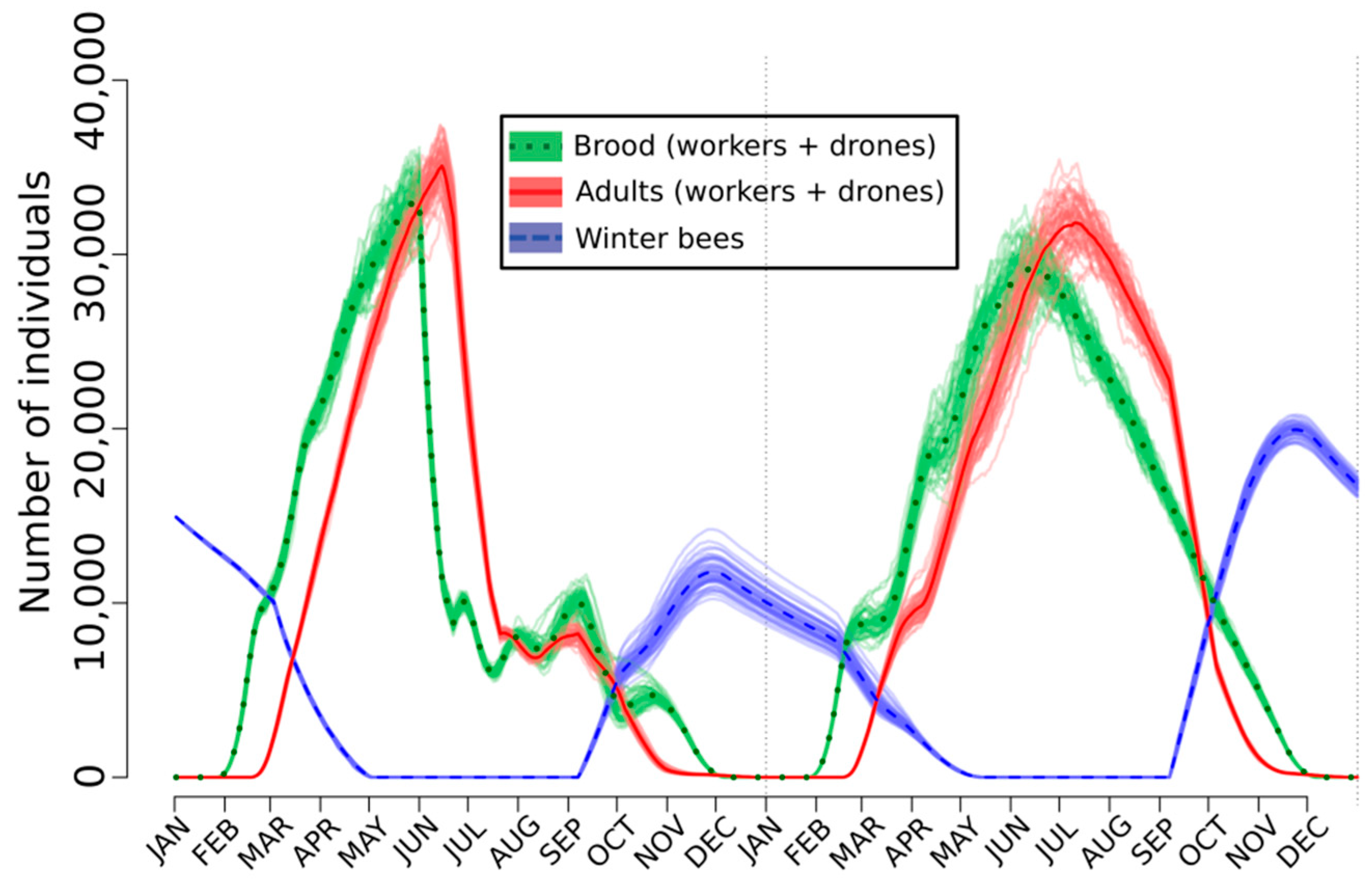
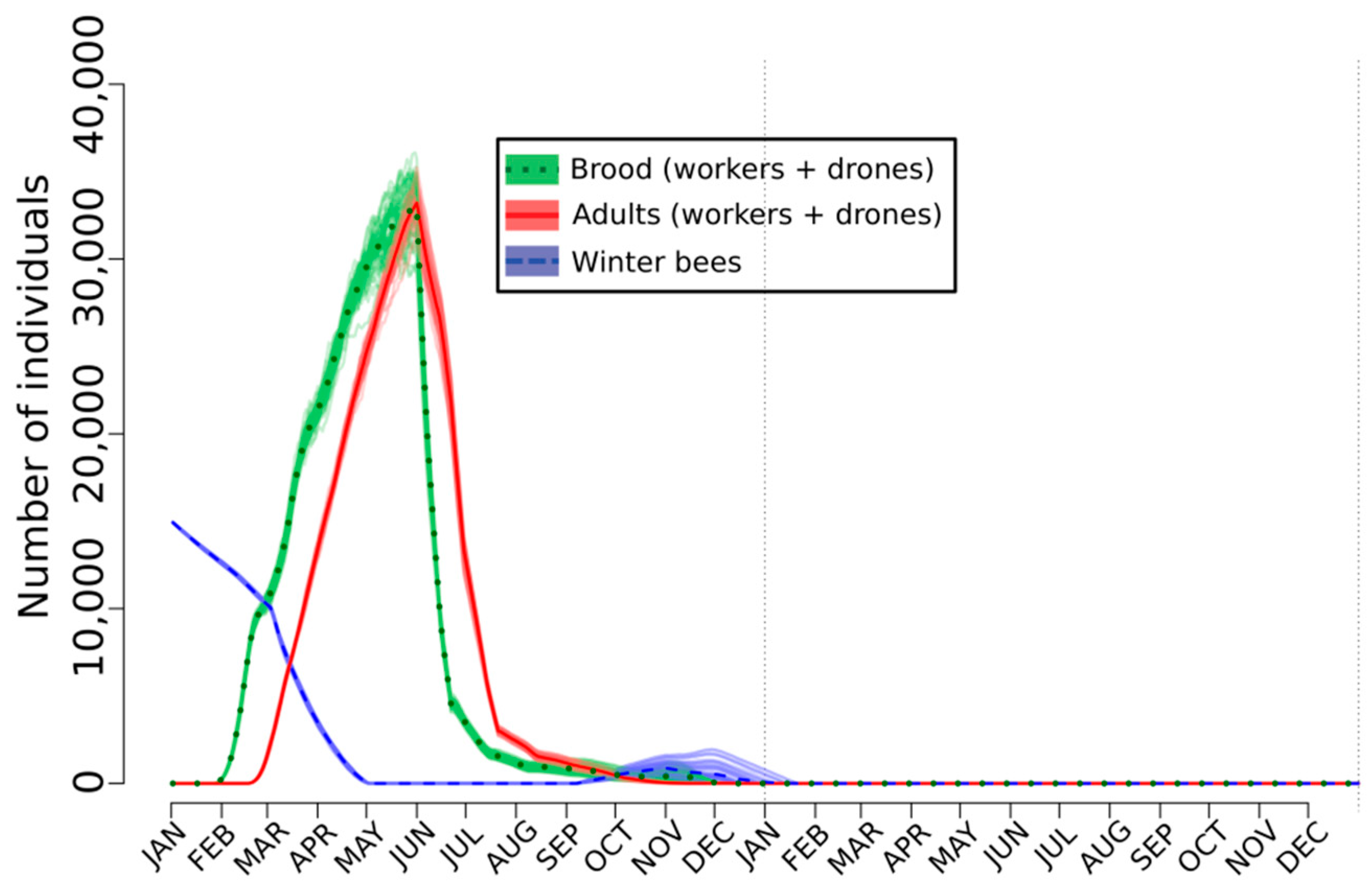
2.3. Lethal, Sublethal, and Behavioral Effects at the Hive Level
3. Effects of Pyriproxyfen on Non-Apis Bees
4. Discussion
5. Conclusions and Recommendations
Author Contributions
Funding
Conflicts of Interest
References
- Bawa, K.S. Plant-pollinator interactions in tropical rain forests. Ann. Rev. Ecol. Syst. 1990, 21, 399–422. [Google Scholar] [CrossRef]
- Roubik, D.W. Pollination of Cultivated Plants in the Tropics; FAO Agricultural Services Bulletin 118; FAO: Rome, Italy, 1995. [Google Scholar]
- Potts, S.G.; Imperatriz-Fonseca, V.L.; Ngo, H.T. IPBE S: The Assessment Report of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services on Pollinators, Pollination and Food Production; Secretariat of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services: Bonn, Germany, 2016; Available online: https://ipbes.net/assessment-reports/pollinators (accessed on 5 August 2020).
- Klein, A.M.; Vaissière, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc. Biol. Sci. 2007, 274, 303–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ollerton, J.; Winfree, R.; Tarrant, S. How many flowering plants are pollinated by animals? Oikos 2011, 120, 321–326. [Google Scholar] [CrossRef]
- Devillers, J.; Morlot, M.; Pham-Delègue, M.H.; Doré, J.C. Classification of monofloral honeys based on their quality control data. Food Chem. 2004, 86, 305–312. [Google Scholar] [CrossRef]
- Ahmad, M.; Bodlah, I.; Mehmood, K.; Sheikh, U.A.A.; Aziz, M.A. Pollination and foraging potential of European bumblebee, Bombus terrestris (Hymenoptera: Apidae) on tomato crop under greenhouse system. Pak. J. Zool. 2015, 47, 1279–1285. [Google Scholar]
- Velthuis, H.H.W.; van Doorn, A. A century of advances in bumblebee domestication and the economic and environmental aspects of its commercialization for pollination. Apidologie 2006, 37, 421–451. [Google Scholar] [CrossRef] [Green Version]
- Heard, T.A. The role of stingless bees in crop pollination. Annu. Rev. Entomol. 1999, 44, 183–206. [Google Scholar] [CrossRef]
- Gonzalez, V.H.; Amith, J.D.; Stein, T.J. Nesting ecology and the cultural importance of stingless bees to speakers of Yoloxóchitl Mixtec, an endangered language in Guerrero, Mexico. Apidologie 2018, 49, 625–636. [Google Scholar] [CrossRef] [Green Version]
- Cruz, D.O.; Freitas, B.M.; da Silva, L.A.; da Silva, E.M.S.; Bomfim, I.G.A. Pollination efficiency of the stingless bee Melipona subnitida on greenhouse sweet pepper. Pesq. Agropecu. Bras. 2005, 40, 1197–1201. [Google Scholar] [CrossRef]
- Wille, A.; Orozco, E.; Raabe, C. Polinización del chayote Sechium edule (Jacq.) Swartz en Costa Rica. Rev. Biol. Trop. 1983, 31, 145–154. [Google Scholar]
- Slaa, E.J.; Sanchez Chaves, L.A.; Malagodi-Braga, K.S.; Hofstede, F.E. Stingless bees in applied pollination: Practice and perspectives. Apidologie 2006, 37, 293–315. [Google Scholar] [CrossRef] [Green Version]
- Pitts-Singer, T.L.; Cane, J.H. The alfalfa leafcutting bee, Megachile rotundata: The world’s most intensively managed solitary bee. Annu. Rev. Entomol. 2011, 56, 221–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cane, J.H. A native ground-nesting bee (Nomia melanderi) sustainably managed to pollinate alfalfa across an intensively agricultural landscape. Apidologie 2008, 39, 315–323. [Google Scholar] [CrossRef] [Green Version]
- Bosch, J.; Kemp, W.P. Developing and establishing bee species as crop pollinators: The example of Osmia spp. (Hymenoptera: Megachilidae) and fruit trees. Bull. Entomol. Res. 2002, 92, 3–16. [Google Scholar]
- Maccagnani, B.; Ladurner, E.; Santi, F.; Burgio, G. Osmia cornuta (Hymenoptera, Megachilidae) as a pollinator of pear (Pyrus communis): Fruit- and seed-set. Apidologie 2003, 34, 207–216. [Google Scholar] [CrossRef] [Green Version]
- Bosch, J.; Kemp, W.P.; Trostle, G.E. Cherry yields and nesting success in an orchard pollinated with Osmia lignaria (Hymenoptera: Megachilidae). J. Econ. Entomol. 2006, 99, 408–413. [Google Scholar] [CrossRef]
- Gruber, B.; Eckel, K.; Everaars, J.; Dorman, C.F. On managing the red mason bee (Osmia bicornis) in apple orchards. Apidologie 2011, 42, 564–576. [Google Scholar] [CrossRef] [Green Version]
- Sedivy, C.; Dorn, S. Towards a sustainable management of bees of the subgenus Osmia (Megachilidae; Osmia) as fruit tree pollinators. Apidologie 2014, 45, 88–105. [Google Scholar] [CrossRef] [Green Version]
- Straub, L.; Williams, G.R.; Pettis, J.; Fries, I.; Neumann, P. Superorganism resilience: Eusociality and susceptibility of ecosystem service providing insects to stressors. Curr. Opin. Insect Sci. 2015, 12, 109–112. [Google Scholar] [CrossRef] [Green Version]
- Villanueva, R.; Roubik, D.W.; Colli-Ucan, W. Extinction of Melipona beecheii and traditional beekeeping in the Yucatán peninsula. Bee World 2005, 86, 35–41. [Google Scholar] [CrossRef]
- Biesmeijer, J.C.; Roberts, S.P.; Reemer, M.; Ohlemüller, R.; Edwards, M.; Peeters, T.; Schaffers, A.P.; Potts, S.G.; Kleukers, R.; Thomas, C.D.; et al. Parallel declines in pollinators and insect-pollinated plants in Britain and The Netherlands. Science 2006, 313, 351–354. [Google Scholar] [CrossRef] [PubMed]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef]
- Winfree, R. The conservation and restoration of wild bees. Ann. N. Y. Acad. Sci. 2010, 1195, 169–197. [Google Scholar] [CrossRef] [PubMed]
- Cameron, S.A.; Lozier, J.D.; Strange, J.P.; Koch, J.B.; Cordes, N.; Solter, L.F.; Griswold, T.L. Patterns of widespread decline in North American bumble bees. Proc. Natl. Acad. Sci. USA 2011, 108, 662–667. [Google Scholar] [CrossRef] [Green Version]
- Smith, K.M.; Loh, E.H.; Rostal, M.K.; Zambrana-Torrelio, C.M.; Mendiola, L.; Daszak, P. Pathogens, pests, and economics: Drivers of honey bee colony declines and losses. Ecohealth 2014, 10, 434–445. [Google Scholar] [CrossRef] [PubMed]
- Goulson, D.; Nicholls, E.; Botías, C.; Rotheray, E.L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 2015, 347, 1255957. [Google Scholar] [CrossRef] [PubMed]
- McMahon, D.P.; Fürst, M.A.; Caspar, J.; Theodorou, P.; Brown, M.J.F.; Paxton, R.J. A sting in the spit: Widespread cross-infection of multiple RNA viruses across wild and managed bees. J. Anim. Ecol. 2015, 84, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Geldmann, J.; González-Varo, J.P. Conserving honey bees does not help wildlife. Science 2018, 359, 392–393. [Google Scholar] [CrossRef]
- Le Féon, V.; Burel, F.; Chifflet, R.; Henry, M.; Ricroch, A.; Vaissière, B.E.; Baudry, J. Solitary bee abundance and species richness in dynamic agricultural landscapes. Agric. Ecosyst. Environ. 2013, 166, 94–101. [Google Scholar] [CrossRef]
- Guzman, A.; Chase, M.; Kremen, C. On-farm diversification in an agriculturally-dominated landscape positively influences specialist pollinators. Front. Sustain. Food Syst. 2019, 15, 10. [Google Scholar] [CrossRef]
- Linsley, E.G.; MacSwain, J.W. The effects of DDT and certain other insecticides on alfalfa pollinators. J. Econ. Entomol. 1947, 40, 358–363. [Google Scholar] [CrossRef] [PubMed]
- Way, M.J.; Synge, A.D. The effects of D.D.T. and of benzene hexachloride on bees. Ann. Appl. Biol. 1948, 35, 94–109. [Google Scholar] [CrossRef] [PubMed]
- Devillers, J.; Pham-Delègue, M.H. Honey Bees: Estimating the Environmental Impact of Chemicals; Taylor & Francis: London, UK, 2002. [Google Scholar]
- Tasei, J.N.; Ripault, G.; Rivault, E. Hazards of imidacloprid seed coating to Bombus terrestris (Hymenoptera: Apidae) when applied to sunflower. J. Econ. Entomol. 2001, 94, 623–627. [Google Scholar] [CrossRef] [PubMed]
- Gels, J.A.; Held, D.W.; Potter, D.A. Hazards of insecticides to the bumble bees Bombus impatiens (Hymenoptera: Apidae) foraging on flowering white clover in turf. J. Econ. Entomol. 2002, 95, 722–728. [Google Scholar] [CrossRef]
- Devillers, J.; Decourtye, A.; Budzinski, H.; Pham-Delègue, M.H.; Cluzeau, S.; Maurin, M. Comparative toxicity and hazards of pesticides to Apis and non-Apis bees. A chemometrical study. SAR QSAR Environ. Res. 2003, 14, 389–403. [Google Scholar] [CrossRef]
- Morandin, L.A.; Winston, M.L.; Franklin, M.T.; Abbott, V.A. Lethal and sub-lethal effects of spinosad on bumble bees (Bombus impatiens Cresson). Pest Manag. Sci. 2005, 61, 619–626. [Google Scholar] [CrossRef]
- Scott-Dupree, C.D.; Conroy, L.; Harris, C.R. Impact of currently used or potentially useful insecticides for canola agroecosystems on Bombus impatiens (Hymenoptera: Apidae), Megachile rotundata (Hymentoptera: Megachilidae), and Osmia lignaria (Hymenoptera: Megachilidae). J. Econ. Entomol. 2009, 102, 177–182. [Google Scholar] [CrossRef] [Green Version]
- Whitehorn, P.R.; O’Connor, S.; Wackers, F.L.; Goulson, D. Neonicotinoid pesticide reduces bumble bee colony growth and queen production. Science 2012, 336, 351–352. [Google Scholar] [CrossRef] [Green Version]
- Baron, G.L.; Raine, N.E.; Brown, M.J.F. Impact of chronic exposure to a pyrethroid pesticide on bumblebees and interactions with a trypanosome parasite. J. Appl. Ecol. 2014, 51, 460–469. [Google Scholar] [CrossRef] [Green Version]
- Laycock, I.; Cotterell, K.C.; O’Shea-Wheller, T.A.; Cresswell, J.E. Effects of the neonicotinoid pesticide thiamethoxam at field-realistic levels on microcolonies of Bombus terrestris worker bumble bees. Ecotoxicol. Environ. Saf. 2014, 100, 153–158. [Google Scholar] [CrossRef] [Green Version]
- Barbosa, W.F.; De Meyer, L.; Guedes, R.N.; Smagghe, G. Lethal and sublethal effects of azadirachtin on the bumblebee Bombus terrestris (Hymenoptera: Apidae). Ecotoxicology 2015, 24, 130–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanley, D.A.; Russell, A.L.; Morrison, S.J.; Rogers, C.; Raine, N.E. Investigating the impacts of field-realistic exposure to a neonicotinoid pesticide on bumblebee foraging, homing ability and colony growth. J. Appl. Ecol. 2016, 53, 1440–1449. [Google Scholar] [CrossRef] [PubMed]
- Botías, C.; David, A.; Hill, E.M.; Goulson, D. Quantifying exposure of wild bumblebees to mixtures of agrochemicals in agricultural and urban landscapes. Environ. Pollut. 2017, 222, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Siviter, H.; Brown, M.J.F.; Leadbeater, E. Sulfoxaflor exposure reduces bumblebee reproductive success. Nature 2018, 561, 109–112. [Google Scholar] [CrossRef]
- Colgan, T.J.; Fletcher, I.K.; Arce, A.N.; Gill, A.N.; Rodrigues, A.R.; Stolle, E.; Chittka, L.; Wurm, Y. Caste- and pesticide-specific effects of neonicotinoid pesticide exposure on gene expression in bumblebees. Mol. Ecol. 2019, 28, 1964–1974. [Google Scholar] [CrossRef] [Green Version]
- Kenna, D.; Cooley, H.; Pretelli, I.; Ramos Rodrigues, A.; Gill, S.D.; Gill, R.J. Pesticide exposure affects flight dynamics and reduces flight endurance in bumblebees. Ecol. Evol. 2019, 9, 5637–5650. [Google Scholar] [CrossRef] [Green Version]
- Camp, A.A.; Batres, M.A.; Williams, W.C.; Lehmann, D.M. Impact of diflubenzuron on Bombus impatiens (Hymenoptera: Apidae) microcolony development. Environ. Entomol. 2020, 49, 203–210. [Google Scholar] [CrossRef]
- Valdovinos-Núñez, G.R.; Quezada-Euán, J.J.; Ancona-Xiu, P.; Moo-Valle, H.; Carmona, A.; Ruiz Sanchez, E. Comparative toxicity of pesticides to stingless bees (Hymenoptera: Apidae: Meliponini). J. Econ. Entomol. 2009, 102, 1737–1742. [Google Scholar] [CrossRef]
- Lourenço, C.T.; Carvalho, S.M.; Malaspina, O.; Nocelli, R.C. Oral toxicity of fipronil insecticide against the stingless bee Melipona scutellaris (Latreille, 1811). Bull. Environ. Contam. Toxicol. 2012, 89, 921–924. [Google Scholar] [CrossRef]
- Barbosa, W.F.; Tomé, H.V.; Bernardes, R.C.; Siqueira, M.A.; Smagghe, G.; Guedes, R.N. Biopesticide-induced behavioral and morphological alterations in the stingless bee Melipona quadrifasciata. Environ. Toxicol. Chem. 2015, 34, 2149–2158. [Google Scholar] [CrossRef]
- Moreno, S.C.; Silvério, F.O.; Lopes, M.C.; Ramos, R.S.; Alvarenga, E.S.; Picanço, M.C. Toxicity of new pyrethroid in pest insects Ascia monuste and Diaphania hyalinata, predator Solenopsis saevissima and stingless bee Tetragonisca angustula. J. Environ. Sci. Health B 2017, 52, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Bernardes, R.C.; Barbosa, W.F.; Martins, G.F.; Lima, M.A.P. The reduced-risk insecticide azadirachtin poses a toxicological hazard to stingless bee Partamona helleri (Friese, 1900) queens. Chemosphere 2018, 201, 550–556. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Escobar, E.; Liedo, P.; Montoya, P.; Méndez-Villarreal, A.; Guzman, M.; Vandame, R.; Sanchez, D. Effect of GF-120 (Spinosad) aerial sprays on colonies of the stingless bee Scaptotrigona mexicana (Hymenoptera: Apidae) and the honey bee (Hymenoptera: Apidae). J. Econ. Entomol. 2018, 111, 1711–1715. [Google Scholar] [CrossRef] [PubMed]
- Padilha, A.C.; Piovesan, B.; Morais, M.C.; de Pazini, J.; Zotti, M.J.; Botton, M.; Grützmacher, A.D. Toxicity of insecticides on Neotropical stingless bees Plebeia emerina (Friese) and Tetragonisca fiebrigi (Schwarz) (Hymenoptera: Apidae: Meliponini). Ecotoxicology 2020, 29, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Torchio, P.F. Relative toxicity of insecticides to the honey bee, alkali bee, and alfalfa leafcutting bee (Hymenoptera: Apidae, Halictidae, Megachilidae). J. Kans. Entomol. Soc. 1973, 46, 446–453. [Google Scholar]
- Johansen, C.A.; Rincker, C.M.; George, D.A.; Mayer, D.F.; Kious, C.W. Effects of aldicarb and its biologically-active metabolites on bees. Environ. Entomol. 1984, 13, 1386–1398. [Google Scholar] [CrossRef]
- Tasei, J.N.; Carre, S.; Bosio, P.G.; Debray, P.; Hariot, J. Effects of the pyrethroid insecticide, WL85871 and phoalone on adults and progeny of the leaf-cutting bee, Megachile rotundata F, pollinator of lucerne. Pestic. Sci. 1987, 21, 119–128. [Google Scholar] [CrossRef]
- Peach, M.L.; Alston, A.G.; Tepedino, V.J. Sublethal effects of carbaryl bran bait on nesting performance, parental investment, and offspring size and sex ratio of the alfalfa leafcutting bee (Hymenoptera: Megachilidae). Environ. Entomol. 1995, 24, 34–39. [Google Scholar] [CrossRef]
- Abbott, V.A.; Nadeau, J.L.; Higo, H.A.; Winston, M.L. Lethal and sublethal effects of imidacloprid on Osmia lignaria and clothianidin on Megachile rotundata (Hymenoptera: Megachilidae). J. Econ. Entomol. 2008, 101, 784–796. [Google Scholar] [CrossRef]
- Huntzinger, C.I.; James, R.R.; Bosch, J.; Kemp, W.P. Fungicide tests on adult alfalfa leafcutting bees (Hymenoptera: Megachilidae). J. Econ. Entomol. 2008, 101, 1088–1094. [Google Scholar] [CrossRef]
- Hodgson, E.W.; Pitts-Singer, T.L.; Barbour, J.D. Effects of the insect growth regulator, novaluron on immature alfalfa leafcutting bees, Megachile rotundata. J. Insect Sci. 2011, 11, 43. [Google Scholar] [CrossRef] [Green Version]
- Artz, D.R.; Pitts-Singer, T.L. Effects of fungicide and adjuvant sprays on nesting behavior in two managed solitary bees, Osmia lignaria and Megachile rotundata. PLoS ONE 2015, 10, e0135688. [Google Scholar] [CrossRef] [PubMed]
- Anderson, N.L.; Harmon-Threatt, A.N. Chronic contact with realistic soil concentrations of imidacloprid affects the mass, immature development speed, and adult longevity of solitary bees. Sci. Rep. 2019, 9, 3724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tesoriero, D.; Maccagnani, B.; Santi, F.; Celli, G. Toxicity of three pesticides on larval instars of Osmia cornuta: Preliminary results. Bull. Insectol. 2003, 56, 169–171. [Google Scholar]
- Ladurner, E.; Bosch, J.; Maini, S.; Kemp, W.P. Assessing delayed and acute toxicity of five formulated fungicides to Osmia lignaria Say and Apis mellifera. Apidologie 2005, 36, 449–460. [Google Scholar] [CrossRef] [Green Version]
- Ladurner, E.; Bosch, J.; Kemp, W.P.; Maini, S. Foraging and nesting behavior of Osmia lignaria (Hymenoptera: Megachilidae) in the presence of fungicides: Cage studies. J. Econ. Entomol. 2008, 101, 647–653. [Google Scholar] [CrossRef]
- Biddinger, D.J.; Robertson, J.L.; Mullin, C.; Frazier, J.; Ashcraft, S.A.; Rajotte, E.G.; Joshi, N.K.; Vaughn, M. Comparative toxicities and synergism of apple orchard pesticides to Apis mellifera (L.) and Osmia cornifrons (Radoszkowski). PLoS ONE 2013, 8, e72587. [Google Scholar] [CrossRef] [Green Version]
- Sandrock, C.; Tanadini, L.G.; Pettis, J.S.; Biesmeijer, J.C.; Potts, S.G. Sublethal neonicotinoid insecticide exposure reduces solitary bee reproductive success. Agric. For. Entomol. 2014, 16, 119–128. [Google Scholar] [CrossRef]
- Jin, N.; Klein, S.; Leimig, F.; Bischoff, G.; Menzel, R. The neonicotinoid clothianidin interferes with navigation of the solitary bee Osmia cornuta in a laboratory test. J. Exp. Biol. 2015, 218, 2821–2825. [Google Scholar] [CrossRef] [Green Version]
- Sgolastra, F.; Tosi, S.; Medrzycki, P.; Porrini, C.; Burgio, G. Toxicity of spirotetramat on solitary bee larvae, Osmia cornuta (Hymenoptera: Megachilidae), in laboratory conditions. J. Apic. Sci. 2015, 59, 73–83. [Google Scholar] [CrossRef] [Green Version]
- Peters, B.; Gao, Z.; Zumkier, U. Large-scale monitoring of effects of clothianidin-dressed oilseed rape seeds on pollinating insects in Northern Germany: Effects on red mason bees (Osmia bicornis). Ecotoxicology 2016, 25, 1679–1690. [Google Scholar] [CrossRef] [Green Version]
- Shaw, F.R. Bee poisoning: A review of the more important literature. J. Econ. Entomol. 1941, 34, 16–21. [Google Scholar] [CrossRef]
- Devillers, J. Acute toxicity of pesticides to honey bees. In Honey Bees: Estimating the Environmental Impact of Chemicals; Devillers, J., Pham-Delègue, M.H., Eds.; Taylor and Francis: London, UK, 2002; pp. 56–66. [Google Scholar]
- Decourtye, A.; Devillers, J.; Cluzeau, S.; Charreton, M.; Pham-Delègue, M.H. Effects of imidacloprid and deltamethrin on associative learning in honeybees under semi-field and laboratory conditions. Ecotoxicol. Environ. Saf. 2004, 57, 410–419. [Google Scholar] [CrossRef]
- Decourtye, A.; Devillers, J.; Genecque, E.; Lemenach, K.; Budzinski, H.; Cluzeau, S.; Pham-Delègue, M.H. Comparative sublethal toxicity of nine pesticides on olfactory learning performances of honeybee Apis mellifera. Arch. Environ. Contam. Toxicol. 2005, 48, 242–250. [Google Scholar] [CrossRef] [PubMed]
- Decourtye, A.; Lefort, S.; Devillers, J.; Gauthier, M.; Aupinel, P.; Tisseur, M. Sublethal effects of fipronil on the ability of honeybees (Apis mellifera L.) to orientate in a complex maze. Julius-Kühn-Archiv 2009, 423, 75–83. [Google Scholar]
- Decourtye, A.; Devillers, J.; Aupinel, P.; Brun, F.; Bagnis, C.; Fourrier, J.; Gauthier, M. Honeybee tracking with microchips: A new methodology to measure the effects of pesticides. Ecotoxicology 2011, 20, 429–437. [Google Scholar] [CrossRef]
- Pham-Delègue, M.H.; Decourtye, A.; Kaiser, L.; Devillers, J. Behavioural methods to assess the effects of pesticides on honey bees. Apidologie 2002, 33, 425–432. [Google Scholar] [CrossRef] [Green Version]
- Devillers, J. Endocrine Disruption Modeling; CRC Press: Boca Raton, FL, USA, 2009. [Google Scholar]
- Horowitz, A.R.; Kontsedalov, S.; Denholm, I.; Ishaaya, I. Dynamics of insecticide resistance in Bemisia tabaci: A case study with the insect growth regulator pyriproxyfen. Pest Manag. Sci. 2002, 58, 1096–1100. [Google Scholar] [CrossRef] [PubMed]
- Richardson, M.L.; Lagos, D.M. Effects of a juvenile hormone analogue, pyriproxyfen, on the apterous form of soybean aphid (Aphis glycines). J. Appl. Entomol. 2007, 131, 297–302. [Google Scholar] [CrossRef]
- Rill, S.; Grafton-Cardwell, E.E.; Morse, J.G. Effects of pyriproxyfen on California red scale (Hemiptera: Diaspididae) development and reproduction. J. Econ. Entomol. 2007, 100, 1435–1443. [Google Scholar] [CrossRef]
- Boina, D.R.; Rogers, M.E.; Wang, N.; Stelinski, L.L. Effect of pyriproxyfen, a juvenile hormone mimic, on egg hatch, nymph development, adult emergence and reproduction of the Asian citrus psyllid, Diaphorina citri Kuwayama. Pest Manag. Sci. 2010, 66, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Liu, N. Insecticide resistance in mosquitoes: Impact, mechanisms, and research directions. Annu. Rev. Entomol. 2015, 60, 537–559. [Google Scholar] [CrossRef] [PubMed]
- Devillers, J.; Larghi, A.; Lagneau, C. QSAR modelling of synergists to increase the efficacy of deltamethrin against pyrethroid-resistant Aedes aegypti mosquitoes. SAR QSAR Environ. Res. 2018, 29, 613–629. [Google Scholar] [CrossRef] [PubMed]
- Devillers, J. 2D and 3D structure-activity modelling of mosquito repellents: A review. SAR QSAR Environ. Res. 2018, 29, 693–723. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.K. Field evaluation of an insect growth regulator, pyriproxyfen, against Aedes togoi larvae in brackish water in South Korea. J. Vector Ecol. 2001, 26, 39–42. [Google Scholar] [PubMed]
- Yapabandara, A.M.; Curtis, C.F. Laboratory and field comparisons of pyriproxyfen, polystyrene beads and other larvicidal methods against malaria vectors in Sri Lanka. Acta Trop. 2002, 81, 211–223. [Google Scholar] [CrossRef]
- Jambulingam, P.; Sadanandane, C.; Boopathi Doss, P.S.; Subramanian, S.; Zaim, M. Field evaluation of an insect growth regulator, pyriproxyfen 0.5% GR against Culex quinquefasciatus, the vector of Bancroftian filariasis in Pondicherry, India. Acta Trop. 2008, 107, 20–24. [Google Scholar] [CrossRef]
- Caputo, B.; Ienco, A.; Cianci, D.; Pombi, M.; Petrarca, V.; Baseggio, A.; Devine, G.J.; Della Torre, A. The “auto-dissemination” approach: A novel concept to fight Aedes albopictus in urban areas. PLoS Negl. Trop. Dis. 2012, 6, e1793. [Google Scholar] [CrossRef]
- Suman, D.S.; Wang, Y.; Faraji, A.; Williams, G.M.; Williges, E.; Gaugler, R. Seasonal field efficacy of pyriproxyfen autodissemination stations against container-inhabiting mosquito Aedes albopictus under different habitat conditions. Pest Manag. Sci. 2018, 74, 885–895. [Google Scholar] [CrossRef]
- Lwetoijera, D.; Kiware, S.; Okumu, F.; Devine, G.J.; Majambere, S. Autodissemination of pyriproxyfen suppresses stable populations of Anopheles arabiensis under semi-controlled settings. Malar. J. 2019, 18, 166. [Google Scholar] [CrossRef] [Green Version]
- Devillers, J. Fate and ecotoxicological effects of pyriproxyfen in aquatic ecosystems. Environ. Sci. Pollut. Res. 2020, 27, 16052–16068. [Google Scholar] [CrossRef] [PubMed]
- Devillers, J. Fate of pyriproxyfen in soils and plants. Toxics 2020, 8, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyamoto, J.; Hirano, H.; Takimoto, Y.; Hatakoshi, M. Insect growth regulators for pest control, with emphasis on juvenile hormone analogs. Present status and future prospect. In Pest Control with Enhanced Environmental Safety; Duke, S.O., Menn, J.J., Plimmer, J.R., Eds.; ACS Syposium Series 524; American Chemical Society: Washington, DC, USA, 1993; pp. 144–168. [Google Scholar]
- WHO. WHO Specifications and Evaluations for Public Health Pesticides: Pyriproxyfen, 4-Phenoxyphenyl (RS)-2-(2-Pyridyloxy)Propyl Ether; WHO Press, World Health Organization: Geneva, Switzerland, 2006. [Google Scholar]
- EC. Directive 98/8/EC Concerning the Placing of Biocidal Products on the Market. Inclusion of Active Substances in Annex I to Directive 98/8/EC. Assessment Report Pyriproxyfen Product-Type 18 (Insecticides, Acaricides and Products to Control Other Arthropods), 21 September 2012, Annex I, The Netherlands. 2012. Available online: http://dissemination.echa.europa.eu/Biocides/ActiveSubstances/0061-18/0061-18_Assessment_Report.pdf (accessed on 5 August 2020).
- EFSA. Conclusion on the Peer Review of the Pesticide Risk Assessment of the Active Substance Pyriproxyfen. EFSA J. 2019, 17, e05732. Available online: https://doi.org/10.2903/j.efsa.2019.5732 (accessed on 5 August 2020).
- Machado Baptista, A.P.; Carvalho, G.A.; Carvalho, S.M.; Carvalho, C.F.; de Souza Bueno Filho, J.S. Toxicidade de produtos fitossanitários utilizados em citros para Apis mellifera. Ciênc. Rural Santa Maria 2009, 39, 955–961. [Google Scholar] [CrossRef]
- Costa, E.M.; Araujo, E.L.; Maia, A.V.P.; Silva, F.E.L.; Bezerra, C.E.S.; Silva, J.G. Toxicity of insecticides used in the Brazilian melon crop to the honey bee Apis mellifera under laboratory condition. Apidologie 2013, 45, 34–44. [Google Scholar] [CrossRef] [Green Version]
- Naiara Gomes, I.; Ingred Castelan Vieira, K.; Moreira Gontijo, L.; Canto Resende, H. Honeybee survival and flight capacity are compromised by insecticides used for controlling melon pests in Brazil. Ecotoxicology 2020, 29, 97–107. [Google Scholar] [CrossRef]
- Fisher, A., 2nd; Colman, C.; Hoffmann, C.; Fritz, B.; Rangel, J. The effects of the insect growth regulators methoxyfenozide and pyriproxyfen and the acaricide bifenazate on honey bee (Hymenoptera: Apidae) forager survival. J. Econ. Entomol. 2018, 111, 510–516. [Google Scholar] [CrossRef]
- Phillips, J.E. Absorption, Accumulation and Sub-Lethal Effects of Juvenile Hormone Analogues in Apis mellifera Honeybees; B Sc Report; University of Canberra: Canberra, Australia, 2013. [Google Scholar]
- Poquet, Y.; Bodin, L.; Tchamitchian, M.; Fusellier, M.; Giroud, B.; Lafay, F.; Buleté, A.; Tchamitchian, S.; Cousin, M.; Pélissier, M.; et al. A pragmatic approach to assess the exposure of the honey bee (Apis mellifera) when subjected to pesticide spray. PLoS ONE 2014, 9, e113728. [Google Scholar] [CrossRef] [Green Version]
- Cunha, J.P.; Chueca, P.; Garcerá, C.; Molto, E. Risk assessment of pesticide spray drift from citrus applications with air-blast sprayers in Spain. Crop Protect. 2012, 42, 116–123. [Google Scholar] [CrossRef]
- Bitondi, M.M.; Mora, I.M.; Simões, Z.L.; Figueiredo, V.L. The Apis mellifera pupal melanization program is affected by treatment with a juvenile hormone analogue. J. Insect Physiol. 1998, 44, 499–507. [Google Scholar] [CrossRef]
- Zufelato, M.S.; Bitondi, M.M.; Simões, Z.L.; Hartfelder, K. The juvenile hormone analog pyriproxyfen affects ecdysteroid-dependent cuticle melanization and shifts the pupal ecdysteroid peak in the honey bee (Apis mellifera). Arthropod Struct. Dev. 2000, 29, 111–119. [Google Scholar] [CrossRef]
- Santos, A.E.; Bitondi, M.M.; Simões, Z.L. Hormone-dependent protein patterns in integument and cuticular pigmentation in Apis mellifera during pharate adult development. J. Insect Physiol. 2001, 47, 1275–1282. [Google Scholar] [CrossRef]
- Boleli, I.C.; Bitondi, M.M.G.; Figueiredo, V.L.C.; Simões, Z.L.P. Mispatterning in the ommatidia of Apis mellifera pupae treated with a juvenile hormone analogue. J. Morphol. 2001, 249, 89–99. [Google Scholar] [CrossRef]
- Pinto, L.Z.; Bitondi, M.M.G.; Simões, Z.L.P. Inhibition of vitellogenin synthesis in Apis mellifera workers by a juvenile hormone analogue, pyriproxyfen. J. Insect Physiol. 2000, 46, 153–160. [Google Scholar] [CrossRef]
- Corrêa Fernandez, F.; Da Cruz-Landim, C.; Malaspina, O. Influence of the insecticide pyriproxyfen on the flight muscle differentiation of Apis mellifera (Hymenoptera, Apidae). Microsc. Res. Tech. 2012, 75, 844–848. [Google Scholar] [CrossRef]
- Aupinel, P.; Fortini, D.; Dufour, H.; Tasei, J.N.; Michaud, B.; Odoux, J.F.; Pham Delègue, M.H. Improvement of artificial feeding in a standard in vitro method for rearing Apis mellifera larvae. Bull. Insectol. 2005, 58, 107–111. [Google Scholar]
- Aupinel, P.; Fortini, D.; Dufour, H.; Michaud, B.; Marolleau, F.; Tasei, J.N.; Odoux, J.F. Toxicity of dimethoate and fenoxycarb to honey bee brood (Apis mellifera), using a new in vitro standardized feeding method. Pest Manag. Sci. 2007, 63, 1090–1094. [Google Scholar] [CrossRef]
- Aupinel, P.; Fortini, D.; Michaud, B.; Mdrzycki, P.; Padovani, E.; Przygoda, D.; Maus, C.; Charrière, J.D.; Kilchenmann, V.; Riesberger-Galle, U.; et al. Honey bee brood ring-test: Method for testing pesticide toxicity on honeybee brood in laboratory conditions. Julius-Kühn-Archiv 2009, 423, 96–102. [Google Scholar]
- Deseyn, J.; Billen, J. Age-dependent morphology and ultrastructure of the hypopharyngeal gland of Apis mellifera workers (Hymenoptera, Apidae). Apidologie 2005, 36, 49–57. [Google Scholar] [CrossRef] [Green Version]
- Fortini, D.; Michaud, B.; Aupinel, P. Comparison of two methods to assess effects of insecticides on hypopharyngeal gland development of honey bee. In Proceedings of the 41st Apimondia Congress, International Federation of Beekeepers’s Associations, Montpellier, France, 26 September–30 October 2009. [Google Scholar]
- Heylen, K.; Gobin, B.; Arckens, L.; Huybrechts, R.; Billen, J. The effects of four crop protection products on the morphology and ultrastructure of the hypopharyngeal gland of the European honeybee, Apis mellifera. Apidologie 2011, 42, 103–116. [Google Scholar] [CrossRef] [Green Version]
- Devillers, J.; Devillers, H.; Decourtye, A.; Fourrier, J.; Aupinel, P.; Fortini, D. Agent-based modeling of the long term effects of pyriproxyfen on honey bee population. In In Silico Bees; Devillers, J., Ed.; CRC Press: Boca Raton, FL, USA, 2014; pp. 179–208. [Google Scholar]
- Utsumi, T.; Miyamoto, M.; Katagi, T. Ecotoxicological risk assessment of pesticides in terrestrial ecosystems. Sumitomo Kagaku 2011, 1, 1–19. [Google Scholar]
- Chen, Y.W.; Wu, P.S.; Yang, E.C.; Nai, Y.S.; Huang, Z.Y. The impact of pyriproxyfen on the development of honey bee (Apis mellifera L.) colony in field. J. Asia Pac. Entomol. 2016, 19, 589–594. [Google Scholar] [CrossRef]
- Ko, C.Y.; Chen, Y.W.; Nai, Y.S. Evaluating the effect of environmental chemicals on honey bee development from the individual to colony level. J. Vis. Exp. 2017, 122, 55296. [Google Scholar] [CrossRef] [PubMed]
- Fourrier, J.; Deschamps, M.; Droin, L.; Alaux, C.; Fortini, D.; Beslay, D.; Le Conte, Y.; Devillers, J.; Aupinel, P.; Decourtye, A. Larval exposure to the juvenile hormone analog pyriproxyfen disrupts acceptance and social behavior performance in adult honeybees. PLoS ONE 2015, 10, e0132985. [Google Scholar] [CrossRef]
- Dani, F.R.; Jones, G.R.; Corsi, S.; Beard, R.; Pradella, D.; Turillazzi, S. Nestmate recognition cues in the honey bee: Differential importance of cuticular alkanes and alkenes. Chem. Senses 2005, 30, 477–489. [Google Scholar] [CrossRef] [Green Version]
- Fléché, C.; Clément, M.C.; Zeggane, S.; Faucon, J.P. Contamination des produits de la ruche et risques pour la santé humaine: Situation en France. Rev. Sci. Tech. Off. Int. Epiz. 1997, 16, 609–619. [Google Scholar] [CrossRef]
- Mullin, C.A.; Frazier, M.; Frazier, J.L.; Ashcraft, S.; Simonds, R.; van Engelsdorp, D.; Pettis, J.S. High levels of miticides and agrochemicals in North American apiaries: Implications for honey bee health. PLoS ONE 2010, 5, e9754. [Google Scholar] [CrossRef] [Green Version]
- Calatayud-Vernich, P.; Calatayud, F.; Simó, E.; Picó, Y. Occurrence of pesticide residues in Spanish beeswax. Sci. Total Environ. 2017, 605–606, 745–754. [Google Scholar] [CrossRef]
- Wiest, L.; Buleté, A.; Giroud, B.; Fratta, C.; Amic, S.; Lambert, O.; Pouliquen, H.; Arnaudguilhem, C. Multi-residue analysis of 80 environmental contaminants in honeys, honeybees and pollens by one extraction procedure followed by liquid and gas chromatography coupled with mass spectrometric detection. J. Chromatogr. A 2011, 1218, 5743–5756. [Google Scholar] [CrossRef]
- Lambert, O.; Piroux, M.; Puyo, S.; Thorin, C.; L’Hostis, M.; Wiest, L.; Buleté, A.; Delbac, F.; Pouliquen, H. Widespread occurrence of chemical residues in beehive matrices from apiaries located in different landscapes of Western France. PLoS ONE 2013, 8, e67007. [Google Scholar] [CrossRef]
- Vidau, C. Surveillance de l’Exposition de l’Abeille aux Pesticides. In Proceedings of the Rencontres Scientifiques de l’Anses, Paris, France, 5 December 2017. [Google Scholar]
- Calatayud-Vernich, P.; Calatayud, F.; Simó, E.; Picó, Y. Pesticide residues in honey bees, pollen and beeswax: Assessing beehive exposure. Environ. Pollut. 2018, 241, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Calatayud-Vernich, P.; Calatayud, F.; Simó, E.; Aguilar, J.A.P.; Picó, Y. Two-year monitoring of pesticide hazard in-hive: High honey bee mortality rates during insecticide poisoning episodes in apiaries located near agricultural settings. Chemosphere 2019, 232, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Valcárcel, A.I.; Martinez-Ferrer, M.T.; Campos-Rivela, J.M.; Hernando Guil, M.D. Analysis of pesticide residues in honeybee (Apis mellifera L.) and in corbicular pollen. Exposure in citrus orchard with an integrated pest management system. Talanta 2019, 204, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Nai, Y.S.; Chen, T.Y.; Chen, Y.C.; Chen, C.T.; Chen, B.Y.; Chen, Y.W. Revealing pesticide residues under high pesticide stress in Taiwan’s agricultural environment probed by fresh honey bee (Hymenoptera: Apidae) pollen. J. Econ. Entomol. 2017, 110, 1947–1958. [Google Scholar] [CrossRef]
- Martinello, M.; Manzinello, C.; Borin, A.; Avram, L.E.; Dainese, N.; Giuliato, H.; Gallina, A.; Mutinelli, F. A Survey from 2015 to 2019 to investigate the occurrence of pesticide residues in dead honeybees and other matrices related to honeybee mortality incidents in Italy. Diversity 2020, 12, 15. [Google Scholar] [CrossRef] [Green Version]
- Devillers, H.; Lobry, J.R.; Menu, F. An agent-based model for predicting the prevalence of Trypanosoma cruzi I and II in their host and vector populations. J. Theor. Biol. 2008, 255, 307–315. [Google Scholar] [CrossRef]
- Devillers, J.; Devillers, H.; Decourtye, A.; Aupinel, P. Internet resources for agent-based modelling. SAR QSAR Environ. Res. 2010, 21, 337–350. [Google Scholar] [CrossRef]
- Beaudouin, R.; Goussen, B.; Piccini, B.; Augustine, S.; Devillers, J.; Brion, F.; Péry, A.R.R. An individual-based model of zebrafish population dynamics accounting for energy dynamics. PLoS ONE 2015, 10, e0125841. [Google Scholar] [CrossRef] [Green Version]
- Devillers, J.; Devillers, H. Population dynamics models for assessing the endocrine disruption potential of juvenile hormone analogues on nontarget species. In Juvenile Hormones and Juvenoids. Modeling Biological Effects and Environmental Fate; Devillers, J., Ed.; CRC Press: Boca Raton, FL, USA, 2013; pp. 115–132. [Google Scholar]
- de Wael, L.; de Greef, M.; van Laere, O. Toxicity of pyriproxifen and fenoxycarb to bumble bee brood using a new method for testing insect growth regulators. J. Apicul. Res. 1995, 34, 3–8. [Google Scholar] [CrossRef]
- Mommaerts, V.; Sterk, G.; Smagghe, G. Bumblebees can be used in combination with juvenile hormone analogues and ecdysone agonists. Ecotoxicology 2006, 15, 513–521. [Google Scholar] [CrossRef]
- Pinto, L.Z.; Hartfelder, K.; Bitondi, M.M.; Simões, Z.L. Ecdysteroid titers in pupae of highly social bees relate to distinct modes of caste development. J. Insect Physiol. 2002, 48, 783–790. [Google Scholar] [CrossRef]
- Mayer, D.F.; Johansen, C.A. How to Reduce Bee Poisoning from Pesticides; A Pacific Northwest Extension Publication: Pullman, WA, USA, 1999. [Google Scholar]
- Gradish, A.E.; van der Steen, J.; Scott-Dupree, C.D.; Cabrera, A.R.; Cutler, G.C.; Goulson, D.; Klein, O.; Lehmann, D.M.; Lückmann, J.; O’Neill, B.; et al. Comparison of pesticide exposure in honey bees (Hymenoptera: Apidae) and bumble bees (Hymenoptera: Apidae): Implications for risk assessments. Environ. Entomol. 2018, 20, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Boyle, N.K.; Pitts-Singer, T.L.; Abbott, J.; Alix, A.; Cox-Foster, D.L.; Hinarejos, S.; Lehmann, D.M.; Morandin, L.; O’Neill, B.; Raine, N.E.; et al. Workshop on pesticide exposure assessment paradigm for non-Apis bees: Foundation and summaries. Environ. Entomol. 2019, 48, 4–11. [Google Scholar] [CrossRef] [Green Version]
- Cham, K.O.; Nocelli, R.C.F.; Borges, L.O.; Viana-Silva, F.E.C.; Tonelli, C.A.M.; Malaspina, O.; Menezes, C.; Fontana, A.S.; Blochtein, B.; Freitas, B.M.; et al. Pesticide exposure assessment paradigm for stingless bees. Environ. Entomol. 2019, 48, 36–48. [Google Scholar] [CrossRef] [PubMed]
- Sgolastra, F.; Hinarejos, S.; Pitts-Singer, T.L.; Boyle, N.K.; Joseph, T.; Lūckmann, J.; Raine, N.E.; Singh, R.; Williams, N.M.; Bosch, J. Pesticide exposure assessment paradigm for solitary bees. Environ. Entomol. 2019, 48, 22–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabino, W.D.E.; Antonini, Y. Nest architecture, life cycle, and natural enemies of the neotropical leafcutting bee Megachile (Moureapis) maculata (Hymenoptera: Megachilidae) in a montane forest. Apidologie 2017, 48, 450–460. [Google Scholar] [CrossRef] [Green Version]
- Ramussen, C.; Camargo, J.M.F. A molecular phylogeny and the evolution of nest architecture and behavior in Trigona s.s. (Hymenoptera: Apidae: Meliponini). Apidologie 2008, 39, 102–118. [Google Scholar] [CrossRef]
- Remillet, M. Catalogue des Insectes Ravageurs des Cultures en Guyane Française; Editions de l’ORSTOM: Paris, France, 1988. [Google Scholar]
- EFSA. Collection and Analysis of Pesticide Residue Data for Pollen and Nectar. Final Report. EFSA Supporting Publication, EN-13032017. 2017. Available online: https://efsa.onlinelibrary.wiley.com/doi/10.2903/sp.efsa.2017.EN-1303 (accessed on 5 August 2020).
- Gierer, F.; Vaughan, S.; Slater, M.; Thompson, H.M.; Elmore, J.S.; Girling, R.D. A review of the factors that influence pesticide residues in pollen and nectar: Future research requirements for optimising the estimation of pollinator exposure. Environ. Pollut. 2019, 249, 236–247. [Google Scholar] [CrossRef] [Green Version]
- Field, J. The evolution of progressive provisioning. Behav. Ecol. 2005, 16, 770–778. [Google Scholar] [CrossRef] [Green Version]
- Neff, J.L. Components of nest provisioning behavior in solitary bees (Hymenoptera: Apoidea). Apidologie 2008, 39, 30–45. [Google Scholar] [CrossRef] [Green Version]
- Dorigo, A.S.; Rosa-Fontana, A.S.; Soares-Lima, H.M.; Galaschi-Teixeira, J.S.; Nocelli, R.C.F.; Malaspina, O. In vitro larval rearing protocol for the stingless bee species Melipona scutellaris for toxicological studies. PLoS ONE 2019, 14, e0213109. [Google Scholar]
- Dharampal, P.S.; Carlson, C.M.; Diaz-Garcia, L.; Steffan, S.A. In vitro rearing of solitary bees: A tool for assessing larval risk factors. J. Vis. Exp. 2018, 137, e57876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, Y.; Diao, Q. Current knowledge of detoxification mechanisms of xenobiotic in honey bees. Ecotoxicology 2017, 26, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Troczka, B.J.; Homem, R.A.; Reid, R.; Beadle, K.; Kolher, M.; Zaworra, M.; Field, L.M.; Williamson, M.S.; Nauen, R.; Emyr Davies, T.G. Identification and functional characterisation of a novel N-cyanoamidine neonicotinoid metabolising cytochrome P450, CYP9Q6, from the buff-tailed bumblebee Bombus terrestris. Insect Biochem. Mol. Biol. 2019, 111, 103171. [Google Scholar] [CrossRef] [PubMed]
- Beadle, K.; Singh, K.S.; Troczka, B.J.; Randall, E.; Zaworra, M.; Zimmer, C.T.; Hayward, A.; Reid, R.; Kor, L.; Kohler, M.; et al. Genomic insights into neonicotinoid sensitivity in the solitary bee Osmia bicornis. PLoS Genet. 2019, 15, e1007903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayward, A.; Beadle, K.; Singh, K.S.; Exeler, N.; Zaworra, M.; Almanza, M.T.; Nikolakis, A.; Garside, C.; Glaubitz, J.; Bass, C.; et al. The leafcutter bee, Megachile rotundata, is more sensitive to N-cyanoamidine neonicotinoid and butenolide insecticides than other managed bees. Nat. Ecol. Evol. 2019, 3, 1521–1524. [Google Scholar] [CrossRef]
- Thompson, H.M.; Wilkins, S.; Battersby, A.H.; Waite, R.J.; Wilkinson, D. The effects of four insect growth-regulating (IGR) insecticides on honeybee (Apis mellifera L.) colony development, queen rearing and drone sperm production. Ecotoxicology 2005, 14, 757–769. [Google Scholar] [CrossRef]
- Kairo, G.; Provost, B.; Tchamitchian, S.; Abdelkader, F.B.; Bonnet, M.; Cousin, M.; Sénéchal, J.; Benet, P.; Kretzschmar, A.; Belzunces, L.P.; et al. Drone exposure to the systemic insecticide fipronil indirectly impairs queen reproductive potential. Sci. Rep. 2016, 6, 31904. [Google Scholar] [CrossRef] [Green Version]
- Milchreit, K.; Ruhnke, H.; Wegener, J.; Bienefeld, K. Effects of an insect growth regulator and a solvent on honeybee (Apis mellifera L.) brood development and queen viability. Ecotoxicology 2016, 25, 530–537. [Google Scholar] [CrossRef]
- Fisher, A., 2nd; Rangel, J. Exposure to pesticides during development negatively affects honey bee (Apis mellifera) drone sperm viability. PLoS ONE 2018, 13, e0208630. [Google Scholar] [CrossRef]
- Alford, D.V. A study of the hibernation of bumblebees (Hymenoptera: Bombidae) in Southern England. J. Anim. Ecol. 1969, 38, 149–170. [Google Scholar] [CrossRef]
- Liu, H.; Yi, X.; Bi, J.; Wang, P.; Liu, N.; Zhou, Z. The enantioselective environmental behavior and toxicological effects of pyriproxyfen in soil. J. Hazard. Mater. 2019, 365, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Becher, M.A.; Twiston-Davies, G.; Penny, T.D.; Goulson, D.; Rotheray, E.L.; Osborne, J.L. Bumble-BEEHAVE: A systems model for exploring multifactorial causes of bumblebee decline at individual, colony, population and community level. J. Appl. Ecol. 2018, 55, 2790–2801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Everaars, J.; Dormann, C.F. Simulation of solitary (non-Apis) bees competing for pollen. In In Silico Bees; Devillers, J., Ed.; CRC Press: Boca Raton, FL, USA, 2014; pp. 209–268. [Google Scholar]
- Everaars, J.; Settele, J.; Dormann, C.F. Fragmentation of nest and foraging habitat affects time budgets of solitary bees, their fitness and pollination services, depending on traits: Results from an individual-based model. PLoS ONE 2018, 13, e0188269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crall, J.D.; de Bivort, B.L.; Dey, B.; Ford Versypt, A.N. Social buffering of pesticides in bumblebees: Agent-based modeling of the effects of colony size and neonicotinoid exposure on behavior within nests. Front. Ecol. Evol. 2019, 7, 51. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Devillers, J.; Devillers, H. Lethal and Sublethal Effects of Pyriproxyfen on Apis and Non-Apis Bees. Toxics 2020, 8, 104. https://doi.org/10.3390/toxics8040104
Devillers J, Devillers H. Lethal and Sublethal Effects of Pyriproxyfen on Apis and Non-Apis Bees. Toxics. 2020; 8(4):104. https://doi.org/10.3390/toxics8040104
Chicago/Turabian StyleDevillers, James, and Hugo Devillers. 2020. "Lethal and Sublethal Effects of Pyriproxyfen on Apis and Non-Apis Bees" Toxics 8, no. 4: 104. https://doi.org/10.3390/toxics8040104
APA StyleDevillers, J., & Devillers, H. (2020). Lethal and Sublethal Effects of Pyriproxyfen on Apis and Non-Apis Bees. Toxics, 8(4), 104. https://doi.org/10.3390/toxics8040104