Effects of Fluoride Exposure on Primary Human Melanocytes from Dark and Light Skin


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture
2.3. MTS Cytotoxicity Assay
2.4. LDH Membrane Damage Assay
2.5. Melanin Production Assay
2.6. Intracellular Tyrosinase Activity
2.7. Intracellular Reactive Oxygen Species (ROS) Levels
2.8. Intracellular Mitochondrial Membrane Potential (MMP) Levels
2.9. Statistical Analysis
3. Results
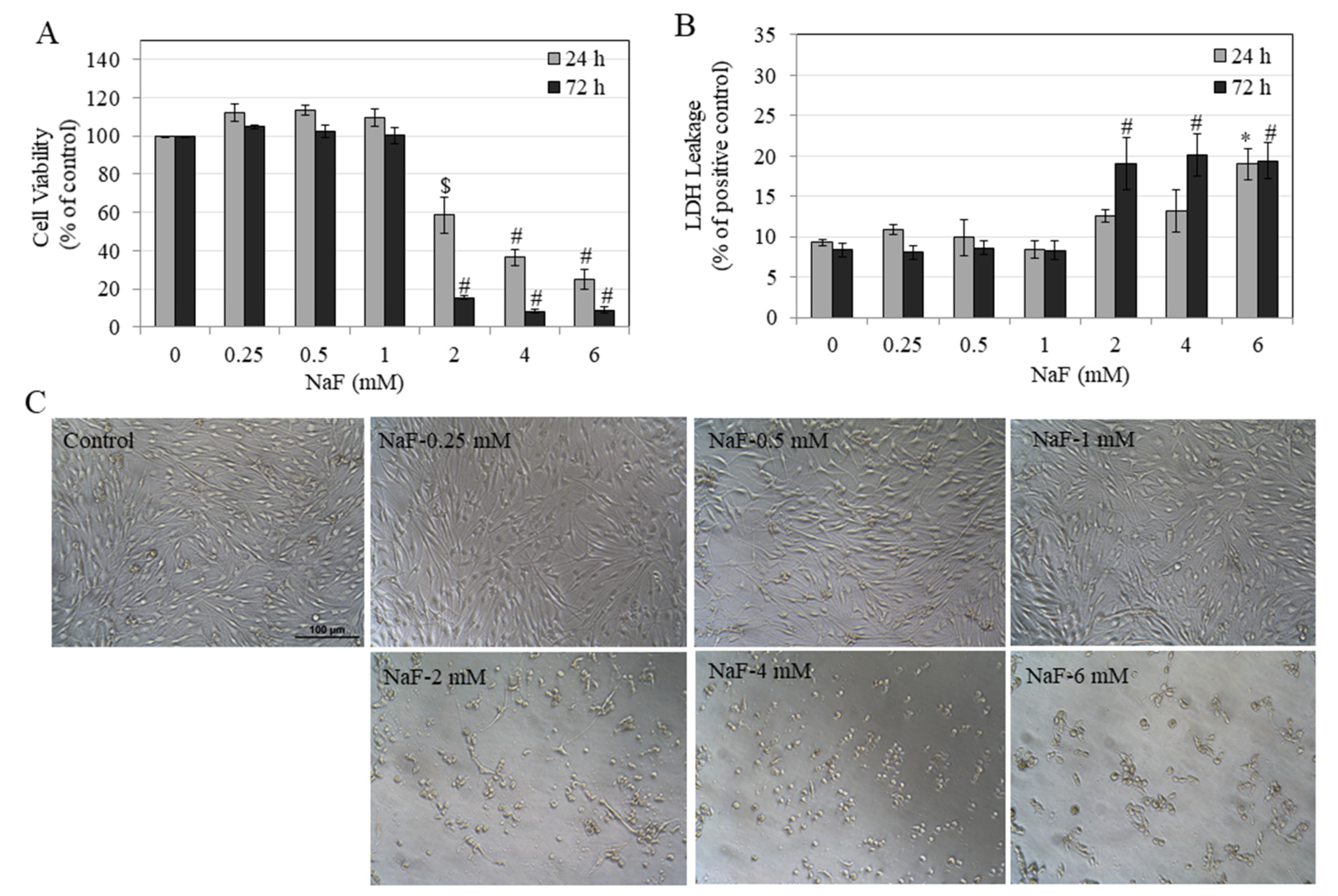
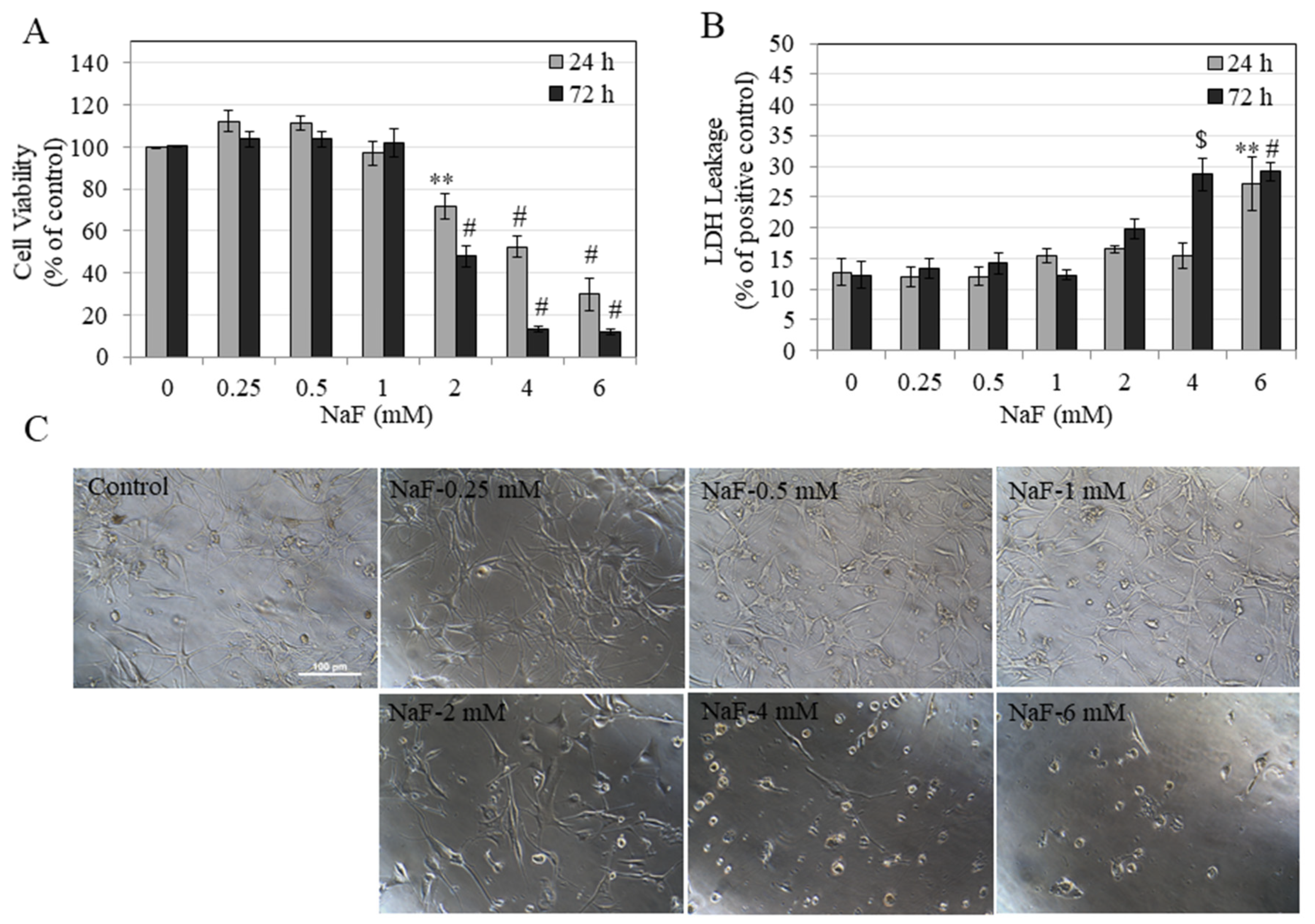
3.1. Effects of NaF on Metabolic Activity and Membrane Inegrity of HEMn-LP Cells
3.2. Effects of NaF on Metabolic Activity and Membrane Integrity of HEMn-DP Cells
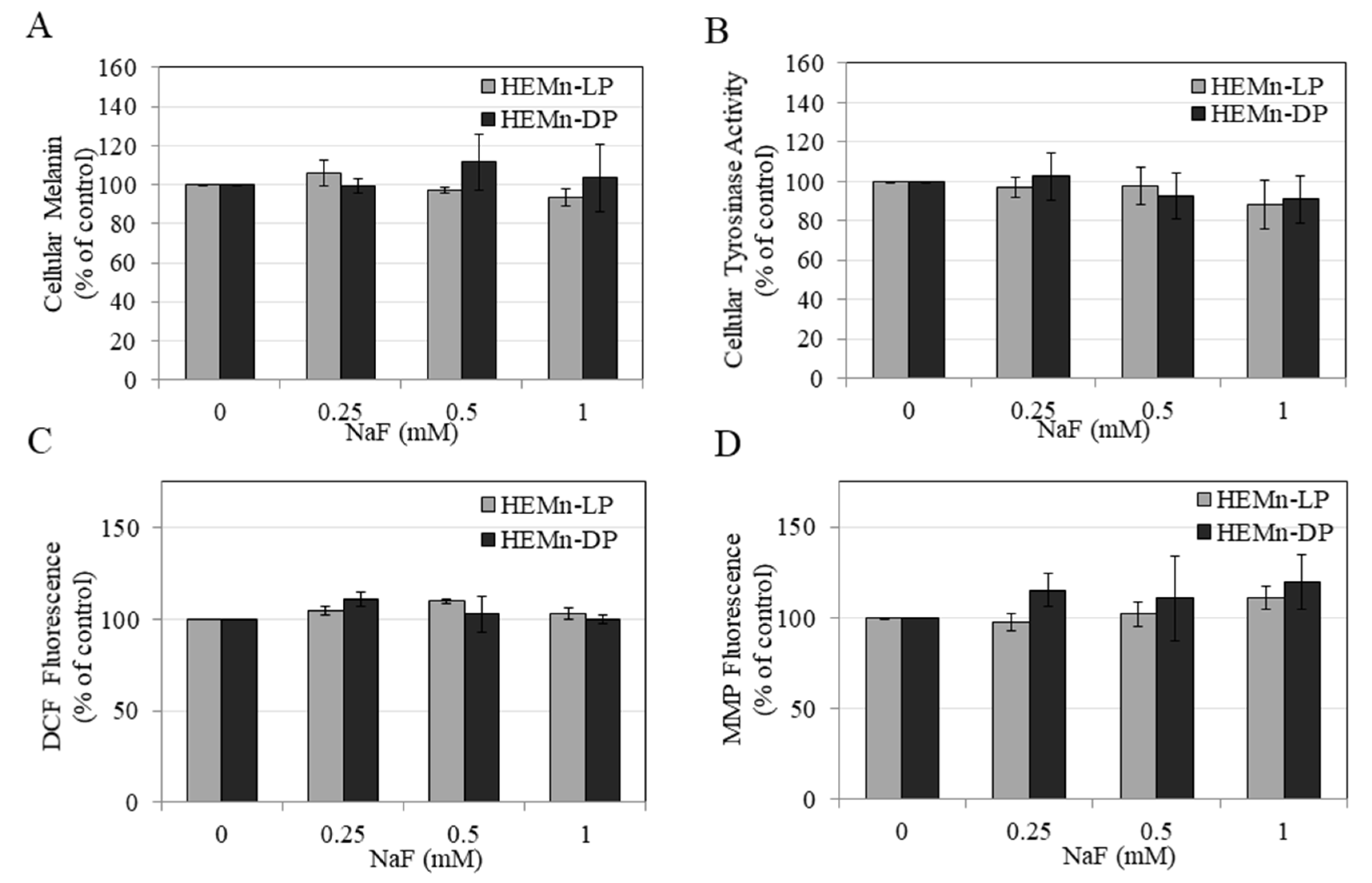
3.3. Effects of NaF on Intracellular Melanin of HEMn-LP and HEMn-DP Cells
3.4. Effects of NaF on Intracellular Tyrosinase Activity in HEMn-LP and HEMn-DP Cells
3.5. Effects of NaF on Intracellular ROS Generation and MMP Levels in HEMn-LP and HEMn-DP Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Zuo, H.; Chen, L.; Kong, M.; Qiu, L.; Lü, P.; Wu, P.; Yang, Y.; Chen, K. Toxic effects of fluoride on organisms. Life Sci. 2018, 198, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Kabir, H.; Gupta, A.K.; Tripathy, S. Fluoride and human health: Systematic appraisal of sources, exposures, metabolism, and toxicity. Crit. Rev. Environ. Sci. Technol. 2020, 50, 1116–1193. [Google Scholar] [CrossRef]
- Spittle, B. The effects of fluoride on inflammation and cancer. Fluoride 2019, 52, 7–8. [Google Scholar]
- Aulestia, F.J.; Groeling, J.; Bomfim, G.H.S.; Costiniti, V.; Manikandan, V.; Chaloemtoem, A.; Concepcion, A.R.; Li, Y.; Wagner, L.E., II; Idaghdour, Y.; et al. Fluoride exposure alters Ca(2+) signaling and mitochondrial function in enamel cells. Sci. Signal. 2020, 13. [Google Scholar] [CrossRef]
- Liao, Y.; Brandt, B.W.; Li, J.; Crielaard, W.; Van Loveren, C.; Deng, D.M. Fluoride resistance in Streptococcus mutans: A mini review. J. Oral Microbiol. 2017, 9, 1344509. [Google Scholar] [CrossRef] [Green Version]
- Strunecka, A.; Strunecky, O. Mechanisms of Fluoride Toxicity: From Enzymes to Underlying Integrative Networks. Appl. Sci. 2020, 10, 7100. [Google Scholar] [CrossRef]
- Johnston, N.R.; Strobel, S.A. Principles of fluoride toxicity and the cellular response: A review. Arch. Toxicol. 2020, 94, 1051–1069. [Google Scholar] [CrossRef]
- Wang, J.; Sánchez-Roselló, M.; Aceña, J.L.; del Pozo, C.; Sorochinsky, A.E.; Fustero, S.; Soloshonok, V.A.; Liu, H. Fluorine in pharmaceutical industry: Fluorine-containing drugs introduced to the market in the last decade (2001–2011). Chem. Rev. 2014, 114, 2432–2506. [Google Scholar] [CrossRef]
- Pan, Y. The Dark Side of Fluorine; ACS Publications: Washington, DC, USA, 2019. [Google Scholar]
- Wermers, R.A.; Cooper, K.; Razonable, R.R.; Deziel, P.J.; Whitford, G.M.; Kremers, W.K.; Moyer, T.P. Fluoride excess and periostitis in transplant patients receiving long-term voriconazole therapy. Clin. Infect. Dis. 2011, 52, 604–611. [Google Scholar] [CrossRef]
- Ron, M.; Singer, L.; Menczel, J.; Kidroni, G. Fluoride concentration in amniotic fluid and fetal cord and maternal plasma. Eur. J. Obstet. Gynecol. Reprod. Biol. 1986, 21, 213–218. [Google Scholar] [CrossRef]
- Jiménez, L.V.; Guzmán, O.L.; Flores, M.C.; Costilla-Salazar, R.; Hernández, J.C.; Contreras, Y.A.; Rocha-Amador, D. In utero exposure to fluoride and cognitive development delay in infants. Neurotoxicology 2017, 59, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Green, R.; Lanphear, B.; Hornung, R.; Flora, D.; Martinez-Mier, E.A.; Neufeld, R.; Ayotte, P.; Muckle, G.; Till, C. Association between maternal fluoride exposure during pregnancy and IQ scores in offspring in Canada. JAMA Pediatr. 2019, 173, 940–948. [Google Scholar] [CrossRef] [PubMed]
- Choi, A.L.; Sun, G.; Zhang, Y.; Grandjean, P. Developmental fluoride neurotoxicity: A systematic review and meta-analysis. Environ. Health Perspect. 2012, 120, 1362–1368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiorillo, L.; Cervino, G.; Herford, A.S.; Laino, L.; Cicciù, M. Stannous fluoride effects on enamel: A systematic review. Biomimetics 2020, 5, 41. [Google Scholar] [CrossRef]
- Croll, T.P.; Nicholson, J.W. Glass ionomer cements in pediatric dentistry: Review of the literature. Pediatr. Dent. 2002, 24, 423–429. [Google Scholar]
- Attar, N.; Turgut, M.D. Fluoride release and uptake capacities of fluoride-releasing restorative materials. Oper. Dent. 2003, 28, 395–402. [Google Scholar]
- Forsten, L. Fluoride release and uptake by glass ionomers. Eur. J. Oral Sci. 1991, 99, 241–245. [Google Scholar] [CrossRef]
- Forsten, L. Short-and long-term fluoride release from glass ionomers and other fluoride-containing filling materials in vitro. Eur. J. Oral Sci. 1990, 98, 179–185. [Google Scholar] [CrossRef]
- Levy, S.M.; McGrady, J.A.; Bhuridej, P.; Warren, J.J.; Heilman, J.R.; Wefel, J.S. Factors affecting dentifrice use and ingestion among a sample of US preschoolers. Pediatr. Dent. 2000, 22, 389–394. [Google Scholar]
- Bentley, E.; Ellwood, R.; Davies, R. Fluoride ingestion from toothpaste by young children. Br. Dent. J. 1999, 186, 460–462. [Google Scholar] [CrossRef]
- Dourson, M.; Charnley, G.; Scheuplein, R. Differential sensitivity of children and adults to chemical toxicity. II. Risk and regulation. Regul. Toxicol. Pharm. 2002, 35, 448–467. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, J.; Newton, R. Serum antioxidant activity in neonates. Arch. Dis. In Child. 1988, 63, 748–750. [Google Scholar] [CrossRef] [Green Version]
- Johanson, C.E. Permeability and vascularity of the developing brain: Cerebellum vs. cerebral cortex. Brain Res. 1980, 190, 3–16. [Google Scholar] [CrossRef]
- Waugh, D.T. Fluoride exposure induces inhibition of sodium-and potassium-activated adenosine triphosphatase (Na+, K+-ATPase) enzyme activity: Molecular mechanisms and implications for public health. Int. J. Environ. Res. Public Health 2019, 16, 1427. [Google Scholar] [CrossRef] [Green Version]
- Fein, S.B.; Grummer-Strawn, L.M.; Raju, T.N. Infant feeding and care practices in the United States: Results from the Infant Feeding Practices Study II. Pediatrics 2008, 122, S25–S27. [Google Scholar] [CrossRef] [Green Version]
- Tapia, J.L.; Neiders, M.E.; Aguirre, A.; McCall, W.D., Jr.; Cohen, R.E. Inflammatory response to dental polishing and prophylaxis materials in rats. J. Int. Acad. Periodontol. 2011, 13, 86–92. [Google Scholar]
- Essman, E.; Essman, W.; Valderrama, E. Histaminergic mediation of the response of rat skin to topical fluorides. Arch. Dermatol. Res. 1981, 271, 325–340. [Google Scholar] [CrossRef]
- Dogan, S.; Gunay, H.; Leyhausen, G.; Geurtsen, W. Chemical-biological interactions of NaF with three different cell lines and the caries pathogen Streptococcus sobrinus. Clin. Oral Investig. 2002, 6, 92–97. [Google Scholar] [CrossRef]
- Prado, E.; Wurtz, T.; Ferbus, D.; Shabana, E.-H.; Forest, N.; Berdal, A. Sodium fluoride influences the expression of keratins in cultured keratinocytes. Cell Biol. Toxicol. 2011, 27, 69–81. [Google Scholar] [CrossRef] [Green Version]
- Cichorek, M.; Wachulska, M.; Stasiewicz, A.; Tymińska, A. Skin melanocytes: Biology and development. Adv. Dermatol. Allergol. 2013, 30, 30. [Google Scholar] [CrossRef]
- Bowman, S.L.; Bi-Karchin, J.; Le, L.; Marks, M.S. The road to lysosome-related organelles: Insights from Hermansky-Pudlak syndrome and other rare diseases. Traffic 2019, 20, 404–435. [Google Scholar] [CrossRef] [Green Version]
- Chang, T.S. An updated review of tyrosinase inhibitors. Int. J. Mol. Sci. 2009, 10, 2440–2475. [Google Scholar] [CrossRef] [Green Version]
- Decker, H.; Tuczek, F. Tyrosinase/catecholoxidase activity of hemocyanins: Structural basis and molecular mechanism. Trends Biochem. Sci. 2000, 25, 392–397. [Google Scholar] [CrossRef]
- Costin, G.-E.; Hearing, V.J. Human skin pigmentation: Melanocytes modulate skin color in response to stress. FASEB J. 2007, 21, 976–994. [Google Scholar] [CrossRef] [PubMed]
- Vancoillie, G.; Lambert, J.; Nayaert, J. Melanocyte biology and its implications for the clinician. Eur. J. Dermatol. 1999, 9, 241–251. [Google Scholar]
- Brenner, M.; Hearing, V.J. The protective role of melanin against UV damage in human skin. Photochem. Photobiol. 2008, 84, 539–549. [Google Scholar] [CrossRef] [Green Version]
- EIObeid, A.S.; Kamal-Eldin, A.; Abdelhalim, M.A.K.; Haseeb, A.M. Pharmacological properties of melanin and its function in health. Basic Clin. Pharmacol. Toxicol. 2017, 120, 515–522. [Google Scholar] [CrossRef]
- Lucky, P.A.; Nordlund, J.J. The biology of the pigmentary system and its disorders. Dermatol. Clin. 1985, 3, 197–216. [Google Scholar] [CrossRef]
- Nicolaidou, E.; Katsambas, A.D. Pigmentation disorders: Hyperpigmentation and hypopigmentation. Clin. Dermatol. 2014, 32, 66–72. [Google Scholar] [CrossRef]
- Lerner, E.A.; Sober, A.J. Chemical and pharmacologic agents that cause hyperpigmentation or hypopigmentation of the skin. Dermatol. Clin. 1988, 6, 327–337. [Google Scholar] [CrossRef]
- Nilima, S.; Vandana, K. Melanin: A scavenger in gingival inflammation. Indian J. Dent. Res. 2011, 22, 38. [Google Scholar]
- Eisen, D. Disorders of pigmentation in the oral cavity. Clin. Dermatol. 2000, 18, 579–587. [Google Scholar] [CrossRef]
- Korzeniewski, C.; Callewaert, D.M. An enzyme-release assay for natural cytotoxicity. J. Immunol. Methods 1983, 64, 313–320. [Google Scholar] [CrossRef]
- Endo, K.; Mizutani, T.; Okano, Y.; Masaki, H. A red pumpkin seed extract reduces melanosome transfer to keratinocytes by activation of Nrf2 signaling. J. Cosmet. Dermatol. 2019, 18, 827–834. [Google Scholar] [CrossRef]
- Scaduto, R.C., Jr.; Grotyohann, L.W. Measurement of mitochondrial membrane potential using fluorescent rhodamine derivatives. Biophys. J. 1999, 76, 469–477. [Google Scholar] [CrossRef] [Green Version]
- Chan, F.K.-M.; Moriwaki, K.; De Rosa, M.J. Detection of necrosis by release of lactate dehydrogenase activity. In Immune Homeostasis; Springer: Berlin/Heidelberg, Germany, 2013; pp. 65–70. [Google Scholar]
- Agalakova, N.I.; Gusev, G.P. Molecular mechanisms of cytotoxicity and apoptosis induced by inorganic fluoride. ISRN Cell Biol. 2012, 2012. [Google Scholar] [CrossRef] [Green Version]
- Inkielewicz-Stepniak, I.; Santos-Martinez, M.J.; Medina, C.; Radomski, M.W. Pharmacological and toxicological effects of co-exposure of human gingival fibroblasts to silver nanoparticles and sodium fluoride. Int. J. Nanomed. 2014, 9, 1677. [Google Scholar]
- Dzierżęga-Lęcznar, A.; Kurkiewicz, S.; Tam, I.; Marek, Ł.; Stępień, K. Pheomelanin content of cultured human melanocytes from lightly and darkly pigmented skin: A pyrolysis-gas chromatography/tandem mass spectrometry study. J. Anal. Appl. Pyrolysis 2017, 124, 349–354. [Google Scholar] [CrossRef]
- Wakamatsu, K.; Kavanagh, R.; Kadekaro, A.L.; Terzieva, S.; Sturm, R.A.; Leachman, S.; Abdel-Malek, Z.; Ito, S. Diversity of pigmentation in cultured human melanocytes is due to differences in the type as well as quantity of melanin. Pigment. Cell Res. 2006, 19, 154–162. [Google Scholar] [CrossRef]
- Alaluf, S.; Heath, A.; Carter, N.; Atkins, D.; Mahalingam, H.; Barrett, K.; Kolb, R.; Smit, N. Variation in melanin content and composition in type V and VI photoexposed and photoprotected human skin: The dominant role of DHI. Pigment. Cell Res. 2001, 14, 337–347. [Google Scholar] [CrossRef]
- Sato, T.; Yagori, A.; Niwa, M. Low sensitivity of cultured human young adult and adult gingival fibroblasts to fluoride. I. Relation to doubling time. Pharmacol. Toxicol. 1987, 61, 313–315. [Google Scholar] [CrossRef]
- Wang, Z.-H.; Li, X.-L.; Yang, Z.-Q.; Xu, M. Fluorine-induced apoptosis and lipid peroxidation in human hair follicles in vitro. Biol. Trace Elem. Res. 2010, 137, 280–288. [Google Scholar] [CrossRef]
- Jiang, P.; Li, G.; Zhou, X.; Wang, C.; Qiao, Y.; Liao, D.; Shi, D. Chronic fluoride exposure induces neuronal apoptosis and impairs neurogenesis and synaptic plasticity: Role of GSK-3β/β-catenin pathway. Chemosphere 2019, 214, 430–435. [Google Scholar] [CrossRef]
- Chen, L.; Ning, H.; Yin, Z.; Song, X.; Feng, Y.; Qin, H.; Li, Y.; Wang, J.; Ge, Y.; Wang, W. The effects of fluoride on neuronal function occurs via cytoskeleton damage and decreased signal transmission. Chemosphere 2017, 185, 589–594. [Google Scholar] [CrossRef]
- Guan, Z. Morphology of the brain of the offspring of rats with chronic fluorosis. Chin. J. Pathol. 1986, 15, 297. [Google Scholar]
- Barbier, O.; Arreola-Mendoza, L.; Del Razo, L.M. Molecular mechanisms of fluoride toxicity. Chem. Biol. Interact. 2010, 188, 319–333. [Google Scholar] [CrossRef]
- Lerner, A.B. Mammalian tyrosinase: Effect of ions on enzyme action. Arch. Biochem. Biophys. 1952, 36, 473–481. [Google Scholar] [CrossRef] [Green Version]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Izquierdo-Vega, J.A.; Sanchez-Gutierrez, M.; Del Razo, L.M. Decreased in vitro fertility in male rats exposed to fluoride-induced oxidative stress damage and mitochondrial transmembrane potential loss. Toxicol. Appl. Pharm. 2008, 230, 352–357. [Google Scholar] [CrossRef]
- Naumova, E.A.; Dickten, C.; Jung, R.; Krauss, F.; Rübesamen, H.; Schmütsch, K.; Sandulescu, T.; Zimmer, S.; Arnold, W.H. Dynamics of fluoride bioavailability in the biofilms of different oral surfaces after amine fluoride and sodium fluoride application. Sci. Rep. 2016, 6, 18729. [Google Scholar] [CrossRef] [Green Version]
- Larsen, L.S.; Baelum, V.; Richards, A.; Nyvad, B. Fluoride in saliva and oral mucosa after brushing with 1450 or 5000 ppm fluoride toothpaste. Caries Res. 2019, 53, 675–681. [Google Scholar]
- Comar, L.P.; de Souza, B.M.; Grizzo, L.T.; Buzalaf, M.A.R.; Magalhaes, A.C. Evaluation of fluoride release from experimental TiF4 and NaF varnishes in vitro. J. Appl. Oral Sci. 2014, 22, 138–143. [Google Scholar] [CrossRef] [Green Version]
- Guth, S.; Hüser, S.; Roth, A.; Degen, G.; Diel, P.; Edlund, K.; Eisenbrand, G.; Engel, K.-H.; Epe, B.; Grune, T. Toxicity of fluoride: Critical evaluation of evidence for human developmental neurotoxicity in epidemiological studies, animal experiments and in vitro analyses. Arch. Toxicol. 2020, 94, 1375–1415. [Google Scholar] [CrossRef]
- Comar, L.P.; Wiegand, A.; Moron, B.M.; Rios, D.; Buzalaf, M.A.; Buchalla, W.; Magalhaes, A.C. In situ effect of sodium fluoride or titanium tetrafluoride varnish and solution on carious demineralization of enamel. Eur. J. Oral Sci. 2012, 120, 342–348. [Google Scholar] [CrossRef]
- Salomão, P.M.A.; de Oliveira, F.A.; Rodrigues, P.D.; Al-Ahj, L.P.; da Silva Gasque, K.C.; Jeggle, P.; Buzalaf, M.A.R.; de Oliveira, R.C.; Edwardson, J.M.; Magalhães, A.C. The cytotoxic effect of TiF4 and NaF on fibroblasts is influenced by the experimental model, fluoride concentration and exposure time. PLoS ONE 2017, 12, e0179471. [Google Scholar] [CrossRef]
- Seiberg, M. Keratinocyte–melanocyte interactions during melanosome transfer. Pigment. Cell Res. 2001, 14, 236–242. [Google Scholar] [CrossRef]
- Gordon, P.R.; Mansur, C.P.; Gilchrest, B.A. Regulation of human melanocyte growth, dendricity, and melanization by keratinocyte derived factors. J. Investig. Dermatol. 1989, 92, 565–572. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Li, G.; Li, Y.; Zhao, Y.; Manthari, R.K.; Wang, J. The Effects of Fluoride on the Gap-Junctional Intercellular Communication of Rats’ Osteoblast. Biol. Trace Elem. Res. 2020, 193, 195–203. [Google Scholar] [CrossRef]
- Hsu, M.; Andl, T.; Li, G.; Meinkoth, J.L.; Herlyn, M. Cadherin repertoire determines partner-specific gap junctional communication during melanoma progression. J. Cell Sci. 2000, 113, 1535–1542. [Google Scholar]
- Tam, I.; Dzierzega-Lecznar, A.; Stepien, K. Differential expression of inflammatory cytokines and chemokines in lipopolysaccharide-stimulated melanocytes from lightly and darkly pigmented skin. Exp. Derm. 2019, 28, 551–560. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Cell Type | IC50, 24 h (mM) | IC50, 72 h (mM) |
|---|---|---|
| HEMn-LP | 3.09 ± 0.45 | 1.68 ± 0.09 |
| HEMn-DP | 3.97 ± 0.61 | 2.05 ± 0.13 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goenka, S.; Simon, S.R. Effects of Fluoride Exposure on Primary Human Melanocytes from Dark and Light Skin. Toxics 2020, 8, 114. https://doi.org/10.3390/toxics8040114
Goenka S, Simon SR. Effects of Fluoride Exposure on Primary Human Melanocytes from Dark and Light Skin. Toxics. 2020; 8(4):114. https://doi.org/10.3390/toxics8040114
Chicago/Turabian StyleGoenka, Shilpi, and Sanford R. Simon. 2020. "Effects of Fluoride Exposure on Primary Human Melanocytes from Dark and Light Skin" Toxics 8, no. 4: 114. https://doi.org/10.3390/toxics8040114
APA StyleGoenka, S., & Simon, S. R. (2020). Effects of Fluoride Exposure on Primary Human Melanocytes from Dark and Light Skin. Toxics, 8(4), 114. https://doi.org/10.3390/toxics8040114