Comparative Analysis of Neurotoxicity of Six Phthalates in Zebrafish Embryos



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Zebrafish Husbandry
2.2. Chemical Exposure and Developmental Toxicity
2.3. Locomotor Response Analysis
2.4. Tg Zebrafish Larvae Assay
2.5. Gene Expression Analysis
2.6. Statistical Analysis
3. Results
3.1. Developmental Toxicity Profiles of Phthalates
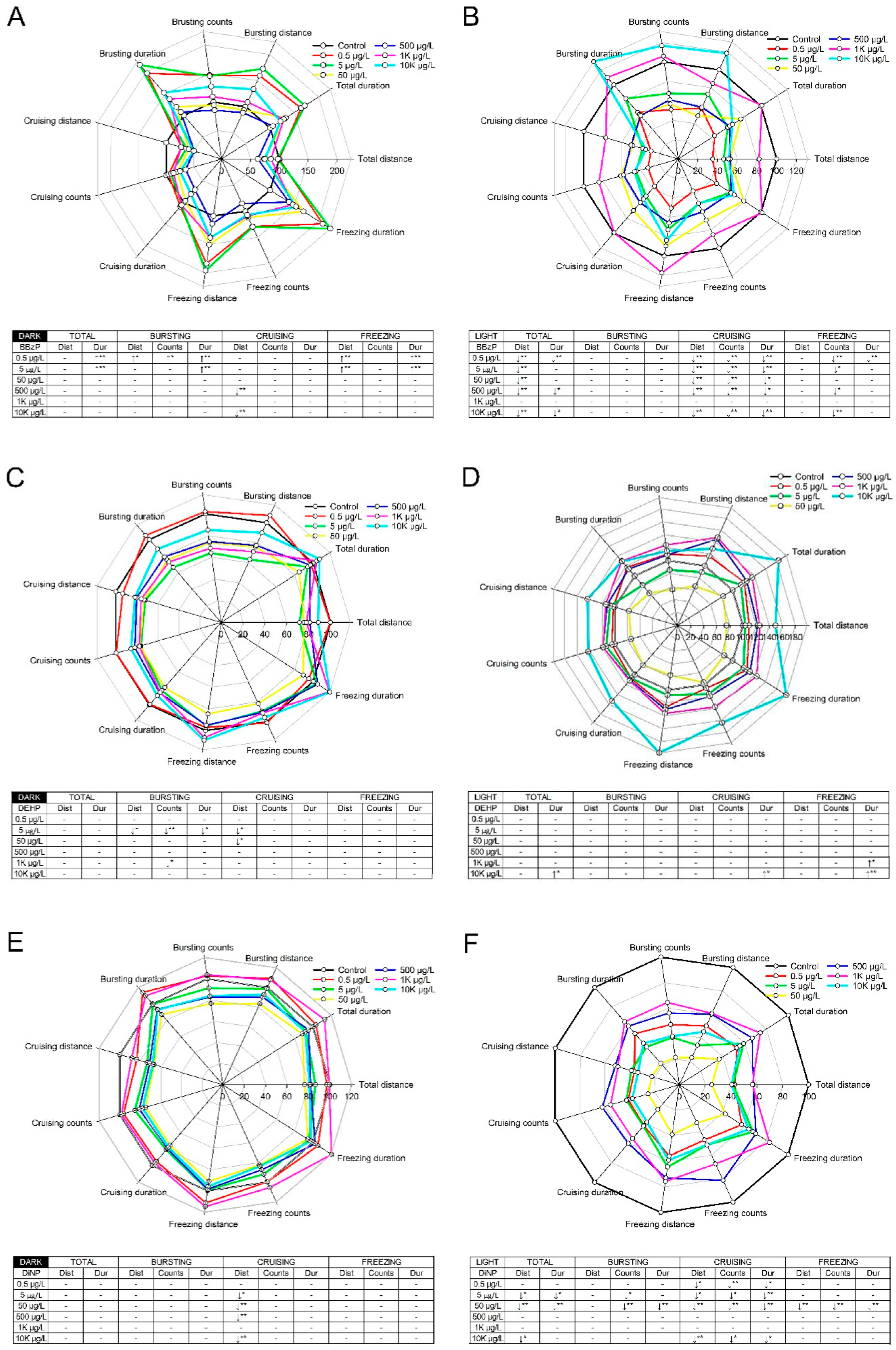
3.2. Neurobehavioral Locomotor Response
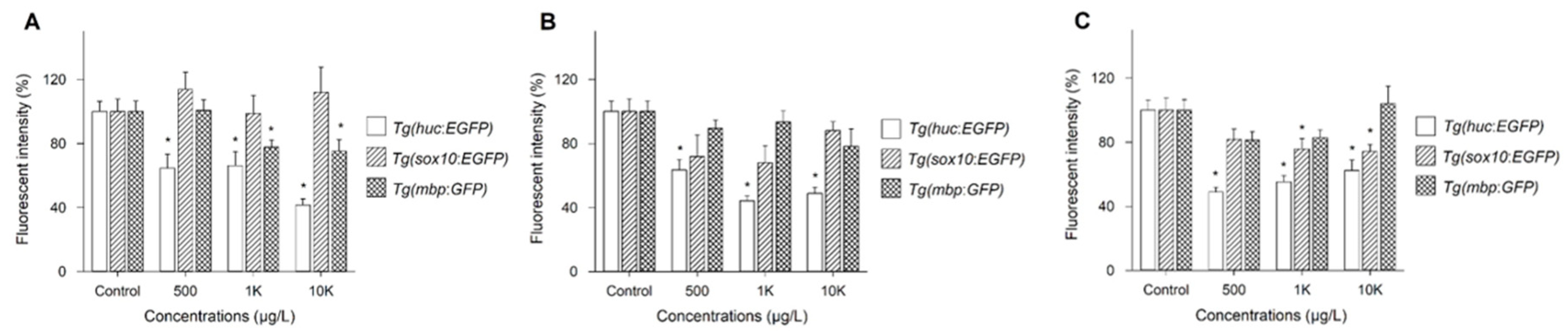
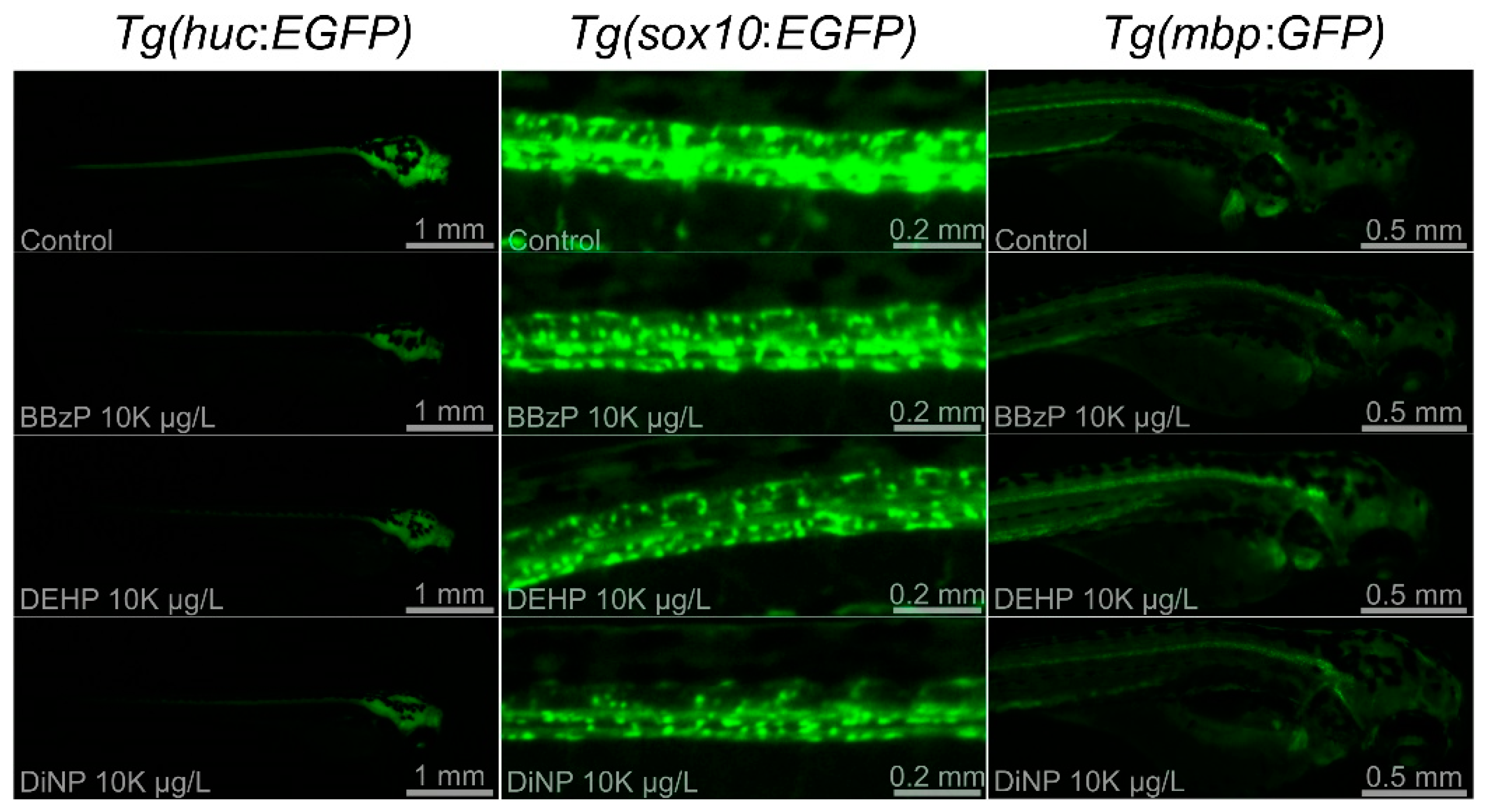
3.3. Changes in Fluorescence Intensity of Three Tg Lines
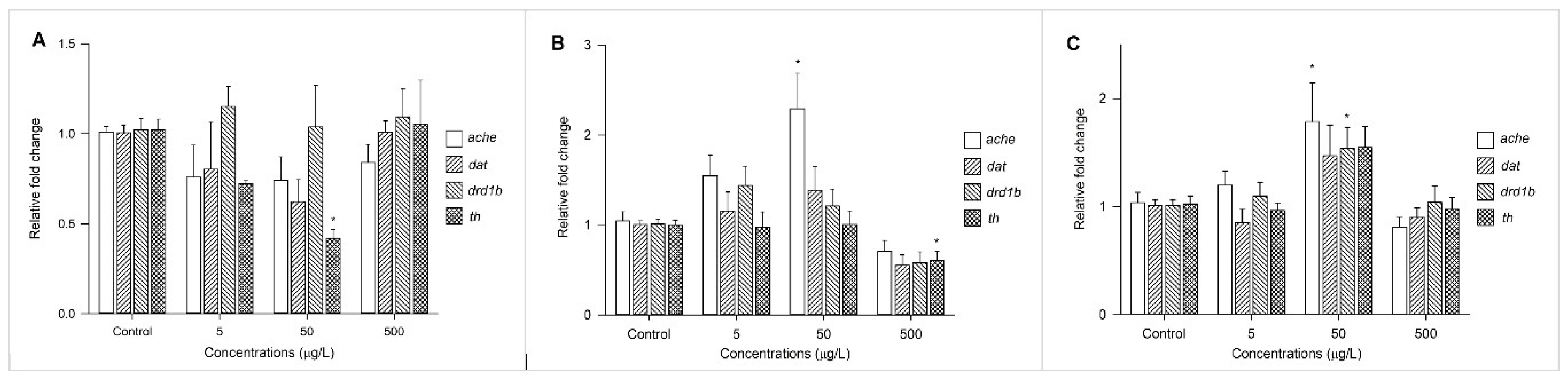
3.4. Changes in Expression Profiles of Genes Involved in Neurotransmission
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vieira, M.G.A.; da Silva, M.A.; dos Santos, L.O.; Beppu, M.M. Natural-based plasticizers and biopolymer films: A review. Eur. Polym. J. 2011, 47, 254–263. [Google Scholar] [CrossRef] [Green Version]
- Carlstedt, F.; Jönsson, B.A.G.; Bornehag, C.-G. PVC flooring is related to human uptake of phthalates in infants. Indoor Air 2013, 23, 32–39. [Google Scholar] [CrossRef]
- Duty, S.M.; Ackerman, R.M.; Calafat, A.M.; Hauser, R. Personal care product use predicts urinary concentrations of some phthalate monoesters. Environ. Health Perspect. 2005, 113, 1530–1535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwapniewski, R.; Kozaczka, S.; Hauser, R.; Silva, M.J.; Calafat, A.M.; Duty, S.M. Occupational exposure to dibutyl phthalate among manicurists. J. Occup. Environ. Med. 2008, 50, 705–711. [Google Scholar] [CrossRef]
- Meeker, J.D.; Sathyanarayana, S.; Swan, S.H. Phthalates and other additives in plastics: Human exposure and associated health outcomes. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2009, 364, 2097–2113. [Google Scholar] [CrossRef] [Green Version]
- Mancuso, M.; Savoca, S.; Bottari, T. First record of microplastics ingestion by European hake merluccius merluccius from the tyrrhenian sicilian coast (central mediterranean sea). J. Fish Biol. 2019, 94, 517–519. [Google Scholar] [CrossRef]
- Leonardo, T.; Teresa., A.; Sheela, S.; Adam, J.S.; Jan, B. Race/ethnicity–specific associations of urinary phthalates with childhood body mass in a nationally representative sample. Environ. Health Perspect. 2013, 121, 501–506. [Google Scholar] [CrossRef]
- Trasande, L.; Spanier, A.J.; Sathyanarayana, S.; Attina, T.M.; Blustein, J. Urinary phthalates and increased insulin resistance in adolescents. Pediatrics 2013, 132, e646–e655. [Google Scholar] [CrossRef] [Green Version]
- Bertelsen, R.J.; Calafat, A.M.; Hoppin, J.A. Urinary biomarkers for phthalates associated with asthma in Norwegian children. Environ. Health Perspect. 2013, 121, 251–256. [Google Scholar] [CrossRef] [Green Version]
- Parks, L.G.; Ostby, J.S.; Lambright, C.R.; Abbott, B.D.; Klinefelter, G.R.; Barlow, N.J.; Gray, L.E., Jr. The plasticizer diethylhexyl phthalate induces malformations by decreasing fetal testosterone synthesis during sexual differentiation in the male rat. Toxicol. Sci. 2000, 58, 339–349. [Google Scholar] [CrossRef]
- Yang, O.; Kim, H.L.; Weon, J.-I.; Seo, Y.R. Endocrine-disrupting chemicals: Review of toxicological mechanisms using molecular pathway analysis. J. Cancer Prev. 2015, 20, 12–24. [Google Scholar] [CrossRef] [PubMed]
- Engel, A.; Buhrke, T.; Imber, F.; Jessel, S.; Seidel, A.; Völkel, W.; Lampen, A. Agonistic and antagonistic effects of phthalates and their urinary metabolites on the steroid hormone receptors ERα, ERβ, and AR. Toxicol. Lett. 2017, 277, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Hannon, P.R.; Brannick, K.E.; Wang, W.; Gupta, R.K.; Flaws, J.A. Di(2-ethylhexyl) phthalate inhibits antral follicle growth, induces atresia, and inhibits steroid hormone production in cultured mouse antral follicles. Toxicol. Appl. Pharmacol. 2015, 284, 42–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meeker, J.D.; Calafat, A.M.; Hauser, R. Urinary metabolites of di(2-ethylhexyl) phthalate are associated with decreased steroid hormone levels in adult men. J. Androl. 2009, 30, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Gore, A.C. Neuroendocrine targets of endocrine disruptors. Hormones 2010, 9, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Miao, W.; Zhu, B.; Xiao, X.; Li, Y.; Dirbaba, N.B.; Zhou, B.; Wu, H. Effects of titanium dioxide nanoparticles on lead bioconcentration and toxicity on thyroid endocrine system and neuronal development in zebrafish larvae. Aquat. Toxicol. 2015, 161, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Carbone, S.; Ponzo, O.J.; Gobetto, N.; Samaniego, Y.A.; Reynoso, R.; Scacchi, P.; Moguilevsky, J.A.; Cutrera, R. Antiandrogenic effect of perinatal exposure to the endocrine disruptor di-(2-ethylhexyl) phthalate increases anxiety-like behavior in male rats during sexual maturation. Horm. Behav. 2013, 63, 692–699. [Google Scholar] [CrossRef]
- Quinnies, K.M.; Harris, E.P.; Snyder, R.W.; Sumner, S.S.; Rissman, E.F. Direct and transgenerational effects of low doses of perinatal di-(2-ethylhexyl) phthalate (DEHP) on social behaviors in mice. PLoS ONE 2017, 12, e0171977. [Google Scholar] [CrossRef]
- Xu, S.; Zhang, H.; Pao, P.-C.; Lee, A.; Wang, J.; Suen Chan, Y.; Manno, F.A.M., III; Wan Chan, S.; Han Cheng, S.; Chen, X. Exposure to phthalates impaired neurodevelopment through estrogenic effects and induced DNA damage in neurons. Aquat. Toxicol. 2020, 222, 105469. [Google Scholar] [CrossRef]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef]
- Kokel, D.; Bryan, J.; Laggner, C.; White, R.; Cheung, C.Y.J.; Mateus, R.; Healey, D.; Kim, S.; Werdich, A.A.; Haggarty, S.J.; et al. Rapid behavior-based identification of neuroactive small molecules in the zebrafish. Nat. Chem. Biol. 2010, 6, 231–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, C.M.; Lee, H.; Lee, B.; Ra, J.-S.; Kim, K.-T. Effects of the chorion on the developmental toxicity of organophosphate esters in zebrafish embryos. J. Hazard. Mater. 2021, 401, 123389. [Google Scholar] [CrossRef]
- Hawliczek, A.; Nota, B.; Cenijn, P.; Kamstra, J.; Pieterse, B.; Winter, R.; Winkens, K.; Hollert, H.; Segner, H.; Legler, J. Developmental toxicity and endocrine disrupting potency of 4-azapyrene, benzo[b]fluorene and retene in the zebrafish danio rerio. Reprod. Toxicol. 2012, 33, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Szewczyńska, M.; Pośniak, M.; Dobrzyńska, E. Determination of phthalates in particulate matter and gaseous phase emitted into the air of the working environment. Int. J. Environ. Sci. Technol. 2020, 17, 175–186. [Google Scholar] [CrossRef] [Green Version]
- Bekö, G.; Weschler, C.J.; Langer, S.; Callesen, M.; Toftum, J.; Clausen, G. Children’s phthalate intakes and resultant cumulative exposures estimated from urine compared with estimates from dust ingestion, inhalation and dermal absorption in their homes and daycare centers. PLoS ONE 2013, 8, e62442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paluselli, A.; Aminot, Y.; Galgani, F.; Net, S.; Sempéré, R. Occurrence of phthalate acid esters (PAEs) in the northwestern Mediterranean sea and the Rhone river. Prog. Oceanogr. 2018, 163, 221–231. [Google Scholar] [CrossRef] [Green Version]
- Council of the European Union. Directive 2010/63/EU of the European Parliament and of the Council of 22 September 2010 on the protection of animals used for scientific purposes. Off. J. Eur. Union 2010, 276, 33–79. [Google Scholar] [CrossRef]
- Braunbeck, T.; Boettcher, M.; Hollert, H.; Kosmehl, T.; Lammer, E.; Leist, E.; Rudolf, M.; Seitz, N. Towards an alternative for the acute fish LC(50) test in chemical assessment: The fish embryo toxicity test goes multi-species—An update. ALTEX 2005, 22, 87–102. [Google Scholar]
- Kim, K.T.; Zaikova, T.; Hutchison, J.E.; Tanguay, R.L. Gold nanoparticles disrupt zebrafish eye development and pigmentation. Toxicol. Sci. 2013, 133, 275–288. [Google Scholar] [CrossRef] [Green Version]
- Park, H.-C.; Kim, C.-H.; Bae, Y.-K.; Yeo, S.-Y.; Kim, S.-H.; Hong, S.-K.; Shin, J.; Yoo, K.-W.; Hibi, M.; Hirano, T.; et al. Analysis of upstream elements in the HuC promoter leads to the establishment of transgenic zebrafish with fluorescent neurons. Dev. Biol. 2000, 227, 279–293. [Google Scholar] [CrossRef] [Green Version]
- Fan, C.-Y.; Cowden, J.; Simmons, S.O.; Padilla, S.; Ramabhadran, R. Gene expression changes in developing zebrafish as potential markers for rapid developmental neurotoxicity screening. Neurotoxicol. Teratol. 2010, 32, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Bin, J.M.; Lyons, D.A. Imaging myelination in vivo using transparent animal models. Brain Plast. 2016, 2, 3–29. [Google Scholar] [CrossRef] [Green Version]
- McCurley, A.T.; Callard, G.V. Characterization of housekeeping genes in zebrafish: Male-female differences and effects of tissue type, developmental stage and chemical treatment. BMC Mol. Biol. 2008, 9, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper—Excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Pu, S.-Y.; Hamid, N.; Ren, Y.-W.; Pei, D.-S. Effects of phthalate acid esters on zebrafish larvae: Development and skeletal morphogenesis. Chemosphere 2020, 246, 125808. [Google Scholar] [CrossRef]
- Chen, X.; Xu, S.; Tan, T.; Lee, S.T.; Cheng, S.H.; Lee, F.W.F.; Xu, S.J.L.; Ho, K.C. Toxicity and estrogenic endocrine disrupting activity of phthalates and their mixtures. Int. J. Environ. Res. Public Health 2014, 11, 3156–3168. [Google Scholar] [CrossRef] [PubMed]
- Dach, K.; Yaghoobi, B.; Schmuck, M.R.; Carty, D.R.; Morales, K.M.; Lein, P.J. Teratological and behavioral screening of the national toxicology program 91-compound library in zebrafish (danio rerio). Toxicol. Sci. 2019, 167, 77–91. [Google Scholar] [CrossRef]
- Qian, L.; Liu, J.; Lin, Z.; Chen, X.; Yuan, L.; Shen, G.; Yang, W.; Wang, D.; Huang, Y.; Pang, S.; et al. Evaluation of the spinal effects of phthalates in a zebrafish embryo assay. Chemosphere 2020, 249, 126144. [Google Scholar] [CrossRef]
- Wibe, Å.E.; Billing, A.; Rosenqvist, G.; Jenssen, B.M. Butyl benzyl phthalate affects shoaling behavior and bottom-dwelling behavior in threespine stickleback. Environ. Res. 2002, 89, 180–187. [Google Scholar] [CrossRef]
- Oehlmann, J.; Schulte-Oehlmann, U.; Kloas, W.; Jagnytsch, O.; Lutz, I.; Kusk, K.O.; Wollenberger, L.; Santos, E.M.; Paull, G.C.; Van Look, K.J.W.; et al. A critical analysis of the biological impacts of plasticizers on wildlife. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2009, 364, 2047–2062. [Google Scholar] [CrossRef] [Green Version]
- Barse, A.V.; Chakrabarti, T.; Ghosh, T.K.; Pal, A.K.; Jadhao, S.B. Endocrine disruption and metabolic changes following exposure of Cyprinus carpio to diethyl phthalate. Pestic. Biochem. Physiol. 2007, 88, 36–42. [Google Scholar] [CrossRef]
- Ogungbemi, A.; Leuthold, D.; Scholz, S.; Küster, E. Hypo- or hyperactivity of zebrafish embryos provoked by neuroactive substances: A review on how experimental parameters impact the predictability of behavior changes. Environ. Sci. Eur. 2019, 31, 88. [Google Scholar] [CrossRef]
- Mwaffo, V.; Zhang, P.; Romero Cruz, S.; Porfiri, M. Zebrafish swimming in the flow: A particle image velocimetry study. PeerJ 2017, 5, e4041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herberholz, J.; Marquart, G.D. Decision making and behavioral choice during predator avoidance. Front. Neurosci. 2012, 6, 125. [Google Scholar] [CrossRef] [Green Version]
- Latash, M.L.; Huang, X. Neural control of movement stability: Lessons from studies of neurological patients. Neuroscience 2015, 301, 39–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haubensak, W.; Kunwar, P.S.; Cai, H.; Ciocchi, S.; Wall, N.R.; Ponnusamy, R.; Biag, J.; Dong, H.-W.; Deisseroth, K.; Callaway, E.M.; et al. Genetic dissection of an amygdala microcircuit that gates conditioned fear. Nature 2010, 468, 270–276. [Google Scholar] [CrossRef] [Green Version]
- Davis, M.; Whalen, P.J. The amygdala: Vigilance and emotion. Mol. Psychiatry 2001, 6, 13–34. [Google Scholar] [CrossRef] [Green Version]
- Roelofs, K. Freeze for action: Neurobiological mechanisms in animal and human freezing. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2017, 372, 20160206. [Google Scholar] [CrossRef] [Green Version]
- Shen, Q.; Truong, L.; Simonich, M.T.; Huang, C.; Tanguay, R.L.; Dong, Q. Rapid well-plate assays for motor and social behaviors in larval zebrafish. Behav. Brain Res. 2020, 391, 112625. [Google Scholar] [CrossRef]
- Prats, E.; Gómez-Canela, C.; Ben-Lulu, S.; Ziv, T.; Padrós, F.; Tornero, D.; Garcia-Reyero, N.; Tauler, R.; Admon, A.; Raldúa, D. Modelling acrylamide acute neurotoxicity in zebrafish larvae. Sci. Rep. 2017, 7, 13952. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, A.M.; Fero, K.; Arrenberg, A.B.; Bergeron, S.A.; Driever, W.; Burgess, H.A. Deep brain photoreceptors control light-seeking behavior in zebrafish larvae. Curr. Biol. 2012, 22, 2042–2047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emran, F.; Rihel, J.; Adolph, A.R.; Wong, K.Y.; Kraves, S.; Dowling, J.E. OFF ganglion cells cannot drive the optokinetic reflex in zebrafish. Proc. Natl. Acad. Sci. USA 2007, 104, 19126–19131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahaboob Basha, P.; Radha, M.J. Gestational di-n-butyl phthalate exposure induced developmental and teratogenic anomalies in rats: A multigenerational assessment. Environ. Sci. Pollut. Res. 2017, 24, 4537–4551. [Google Scholar] [CrossRef] [PubMed]
- Bissegger, S.; Pineda Castro, M.A.; Yargeau, V.; Langlois, V.S. Phthalates modulate steroid 5-reductase transcripts in the Western clawed frog embryo. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2018, 213, 39–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coffey, C.M.; Solleveld, P.A.; Fang, J.; Roberts, A.K.; Hong, S.-K.; Dawid, I.B.; Laverriere, C.E.; Glasgow, E. Novel oxytocin gene expression in the hindbrain is induced by alcohol exposure: Transgenic zebrafish enable visualization of sensitive neurons. PLoS ONE 2013, 8, e53991. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Huang, C.; Wang, X.; Chen, J.; Bai, C.; Chen, Y.; Chen, X.; Dong, Q.; Yang, D. BDE-47 disrupts axonal growth and motor behavior in developing zebrafish. Aquat. Toxicol. 2012, 120–121, 35–44. [Google Scholar] [CrossRef]
- Kwak, J.; Park, O.K.; Jung, Y.J.; Hwang, B.J.; Kwon, S.-H.; Kee, Y. Live image profiling of neural crest lineages in zebrafish transgenic lines. Mol. Cells 2013, 35, 255–260. [Google Scholar] [CrossRef] [Green Version]
- Jung, S.-H.; Kim, S.; Chung, A.-Y.; Kim, H.-T.; So, J.-H.; Ryu, J.; Park, H.-C.; Kim, C.-H. Visualization of myelination in GFP-transgenic zebrafish. Dev. Dyn. 2010, 239, 592–597. [Google Scholar] [CrossRef]
- Honoré, S.M.; Aybar, M.J.; Mayor, R. Sox10 is required for the early development of the prospective neural crest in Xenopus embryos. Dev. Biol. 2003, 260, 79–96. [Google Scholar] [CrossRef] [Green Version]
- Zada, D.; Tovin, A.; Lerer-Goldshtein, T.; Vatine, G.D.; Appelbaum, L. Altered behavioral performance and live imaging of circuit-specific neural deficiencies in a zebrafish model for psychomotor retardation. PLOS Genet. 2014, 10, e1004615. [Google Scholar] [CrossRef] [Green Version]
- Behra, M.; Cousin, X.; Bertrand, C.; Vonesch, J.-L.; Biellmann, D.; Chatonnet, A.; Strähle, U. Acetylcholinesterase is required for neuronal and muscular development in the zebrafish embryo. Nat. Neurosci. 2002, 5, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Goldman-Rakic, P.S.; Castner, S.A.; Svensson, T.H.; Siever, L.J.; Williams, G.V. Targeting the dopamine D1 receptor in schizophrenia: Insights for cognitive dysfunction. Psychopharmacology 2004, 174, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Mexas, L.M.; Florang, V.R.; Doorn, J.A. Inhibition and covalent modification of tyrosine hydroxylase by 3,4-dihydroxyphenylacetaldehyde, a toxic dopamine metabolite. Neurotoxicology 2011, 32, 471–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabrez, S.; Jabir, N.R.; Shakil, S.; Greig, N.H.; Alam, Q.; Abuzenadah, A.M.; Damanhouri, G.A.; Kamal, M.A. A synopsis on the role of tyrosine hydroxylase in Parkinson’s disease. CNS Neurol. Disord. Drug Targets 2012, 11, 395–409. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tran, C.M.; Do, T.N.; Kim, K.-T. Comparative Analysis of Neurotoxicity of Six Phthalates in Zebrafish Embryos. Toxics 2021, 9, 5. https://doi.org/10.3390/toxics9010005
Tran CM, Do TN, Kim K-T. Comparative Analysis of Neurotoxicity of Six Phthalates in Zebrafish Embryos. Toxics. 2021; 9(1):5. https://doi.org/10.3390/toxics9010005
Chicago/Turabian StyleTran, Cong Minh, Trinh Ngoc Do, and Ki-Tae Kim. 2021. "Comparative Analysis of Neurotoxicity of Six Phthalates in Zebrafish Embryos" Toxics 9, no. 1: 5. https://doi.org/10.3390/toxics9010005
APA StyleTran, C. M., Do, T. N., & Kim, K. -T. (2021). Comparative Analysis of Neurotoxicity of Six Phthalates in Zebrafish Embryos. Toxics, 9(1), 5. https://doi.org/10.3390/toxics9010005