
Microbial Transglutaminase Is a Very Frequently Used Food Additive and Is a Potential Inducer of Autoimmune/Neurodegenerative Diseases



Abstract
:1. Introduction
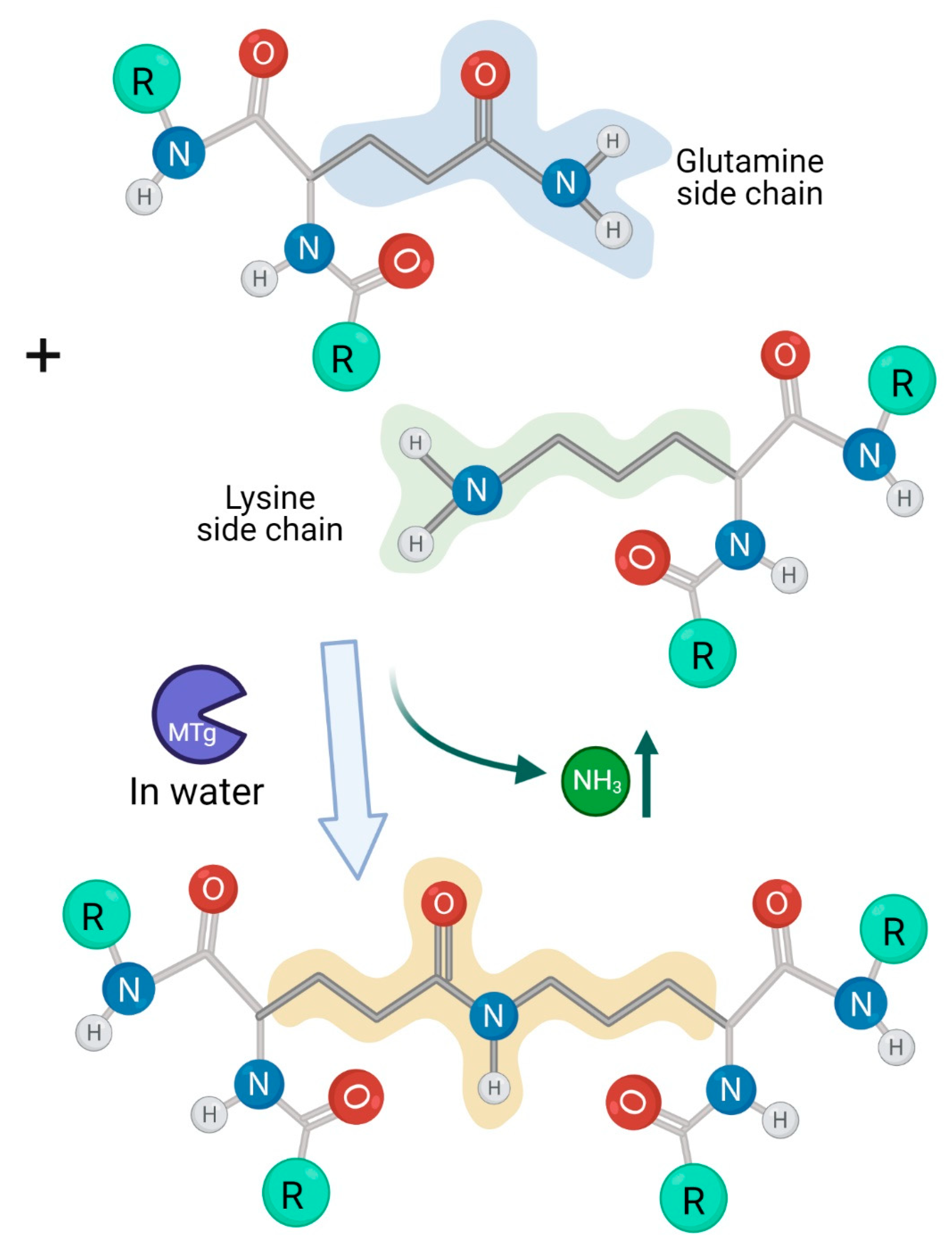
1.1. The Luminal Microbial Transglutaminase Sources in the Human Gut
1.2. Microbial Transglutaminase-Gliadin Complexes Are Immunogenic in Celiac Disease
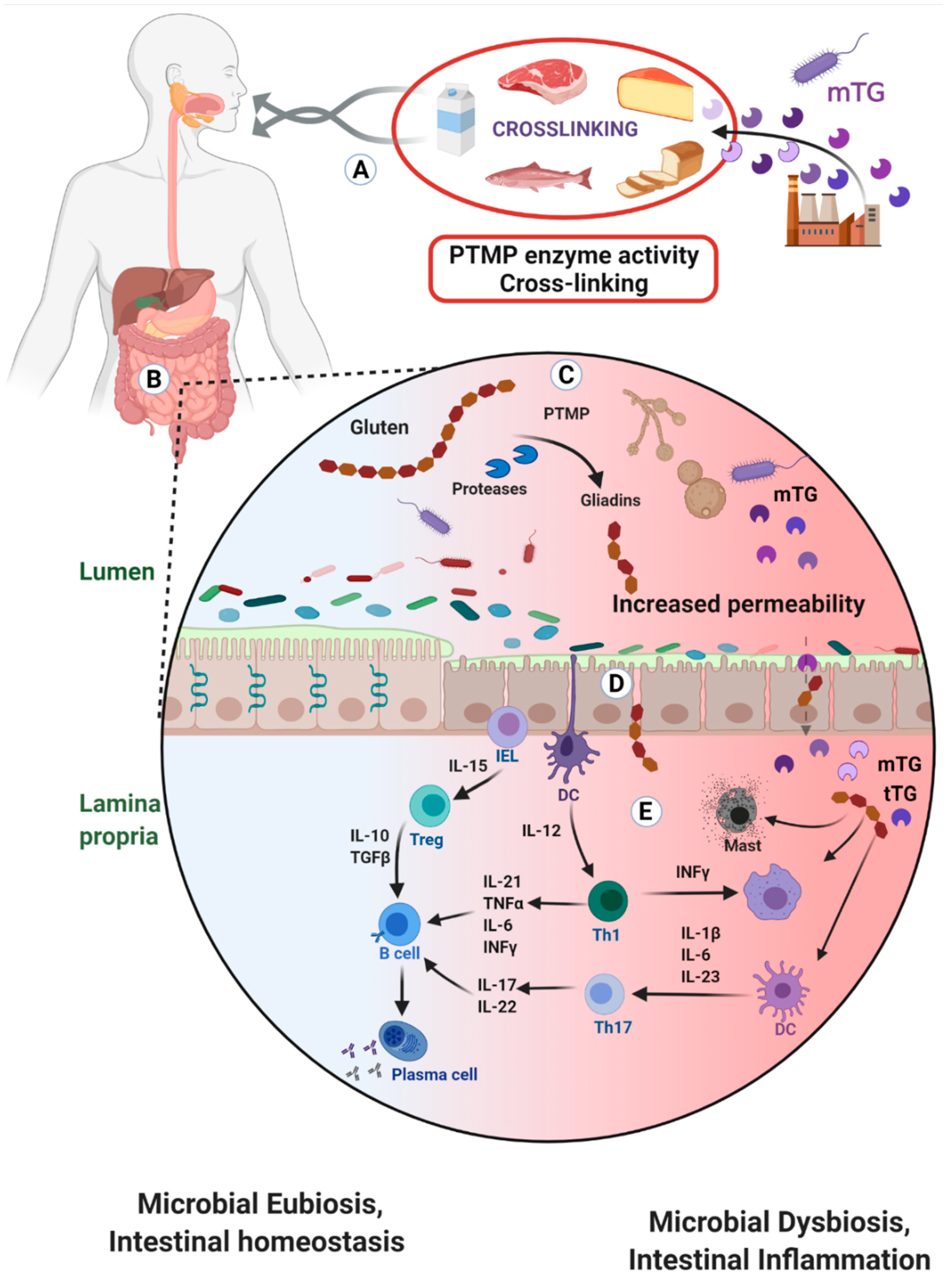
2. Microbial Transglutaminase Cross-Linked Complexes Are Pathogenic
2.1. Trans-Enterocytic Transport of Gliadin and mTG
2.2. Compromised Tight Junction Functional Integrity
- Pathogenic prokaryotes are powerful disruptors of human intestinal permeability [61,62]. Since mTG present a survival factor for the luminal microbes and since the mTG compromises some basic enteric physical and immune protective mechanisms, it might support luminal and mucosal pathobionts activities;
- Gliadins and gluten are known to open the tight junction gap by stimulating zonulin release [61]. As an integral part of the mTG-gliadin neo-complex, the gluten/gliadin part of the complex can drive gut permeability. It should be noticed that this mechanism is not only shared between the CD patient, but also by their closed relative and to some degree the broader normal population [63,64];
2.3. Enhances Enteric Epithelial Gliadins Uptake and Transportation
2.4. Suppression of Mechanical and Immunological Enteric Protective Barriers
2.5. Contributes to Luminal Microbiotic, Dysbiotic and Pathobiotic Proliferation
2.6. Potential mTG-Gliadin Complexes Uptake and Presentation by Mucosal Dendritic Cells
3. Is mTG Active in the Human Gut Lumen?
- Are the mTG-gliadins cross-linked complexes destroyed in the stomach? As mentioned above, those covalent iso-peptide bonds are extremely resistant to the luminal proteases, reducing agents and detergents;
- Microbial transglutaminase is temperature dependent and is active up to 60° Celsius. In reality, many food products are not boiled before consumption or during processing, and some populations prefer eating raw meat. Just as a reminder, analyzing supermarket shelves’ meat and meat products, many were found to contain transglutaminase [29]. Intriguingly, mTG gliadin docked complexes turn more immunogenic when heated to 90° Celsius [10,52]. It is logical to speculate that during denaturation, epitopes are exteriorized and are exposed to the immune system. Regarding mTG activity and temperature, the newly identified cold Atlantic cod TG opens a new area of thermostable mTG application for boiled/heated/cooked food product’s manufacturing [93];
- Microbial transglutaminase is active at pH-4.0 and above. However, gastric physiology and pathophysiology show that upon eating or post-prandially, gastric acidity is neutralized. Large pediatric, adult and elderly people are chronically consuming acid suppressor medications, infants and elderly have higher gastric pH and alkaline reflux is not rare. Notably, the stomach pH is differentially distributed and some areas are less acidic [10]. In summary, it is suggested that active mTG can execute its functions in the duodenum, small and large bowel. The cross-linked complexes are created ex-vivo, while processing the food, they are stomach passage resistant and are immunogenic.
4. Should mTG Usage Be Labeled and Declared on Food Products?
5. Should the Customers Be Warned for a Potential Health Risk of mTG Consumption?
6. Warnings for Use of Microbial Transglutaminase
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Yu, J.; Pian, Y.; Ge, J.; Guo, J.; Zheng, Y.; Jiang, H.; Hao, H.; Yuan, Y.; Jiang, Y.; Yang, M. Functional and structural characterization of the antiphagocytic properties of a novel transglutaminase from streptococcus suis. J. Biol. Chem. 2015, 290, 19081–19092. [Google Scholar]
- Xia, X.; Qin, W.; Zhu, H.; Wang, X.; Jiang, J.; Hu, J. How Streptococcus suis serotype 2 attempts to avoid attack by host immune defenses. J. Microbiol. Immunol. Infect. 2019, 52, 516–525. [Google Scholar] [CrossRef] [PubMed]
- Lerner, A.; Matthias, T. Microbial Transglutaminase is Beneficial to Food Industries but a Caveat to Public Health. Med One 2019, 4, e190001. [Google Scholar]
- Doti, N.; Caporale, A.; Monti, A.; Sandomenico, A.; Selis, F.; Ruvo, M. A recent update on the use of microbial transglutaminase for the generation of biotherapeutics. World J. Microbiol. Biotechnol. 2020, 36, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Miwa, N. Innovation in the food industry using microbial transglutaminase: Keys to success and future prospects. Anal. Biochem. 2020, 597, 113638. [Google Scholar] [CrossRef] [PubMed]
- Deweid, L.; Avrutina, O.; Kolmar, H. Microbial transglutaminase for biotechnological and biomedical engineering. Biol. Chem. 2019, 400, 257–274. [Google Scholar] [CrossRef]
- Steffen, W.; Ko, F.C.; Patel, J.; Lyamichev, V.; Albert, T.J.; Benz, J.; Rudolph, M.G.; Bergmann, F.; Streidl, T.; Kratzsch, P.; et al. Discovery of a microbial transglutaminase enabling highly site-specific labeling of proteins. J. Biol. Chem. 2017, 292, 15622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, S.K.; Lim, T.S. Bioengineering of microbial transglutaminase for biomedical applications. Appl. Microbiol. Biotechnol. 2019, 103, 2973–2984. [Google Scholar] [CrossRef]
- Lerner, A.; Matthias, T. Changes in intestinal tight junction permeability associated with industrial food additives explain the rising incidence of autoimmune disease. Autoimmun. Rev. 2015, 14, 479–489. [Google Scholar] [CrossRef] [Green Version]
- Lerner, A.; Matthias, T. Microbial transglutaminase should be considered as an environmental inducer of celiac disease. World J. Clin. Cases 2019, 7, 3912–3914. [Google Scholar] [CrossRef]
- Lerner, A.; Matthias, T. Microbial transglutaminase: A new potential player in celiac disease. Clin. Immunol. 2019, 199, 37–43. [Google Scholar]
- Lerner, A.; Matthias, T. Processed food additive microbial transglutaminase and its cross-linked gliadin complexes are potential public health concerns in celiac disease. Int. J. Mol. Sci. 2020, 21, 1127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matthias, T.; Lerner, A. Microbial Transglutaminase Is Immunogenic and Potentially Pathogenic in Pediatric Celiac Disease. Front. Pediatr. 2018, 6, 389. [Google Scholar]
- Lerner, A.; Matthias, T. Don’t forget the exogenous microbial transglutaminases: It is immunogenic and potentially pathogenic. AIMS Biophys. 2016, 3, 546–552. [Google Scholar] [CrossRef]
- Lerner, A.; Benzvi, C. “Let Food Be Thy Medicine”: Gluten and Potential Role in Neurodegeneration. Cells 2021, 10, 756. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.; de Vivo, G.D.; Ricotta, M.; Iannuzzi, M.; Gentile, V. Transglutaminases as possible therapeutic targets in neurodegenerative diseases. Recent Pat. CNS Drug Discov. 2010, 3, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Lerner, A.; Aminov, R.; Matthias, T. Dysbiosis may trigger autoimmune diseases via inappropriate post-translational modification of host proteins. Front. Microbiol. 2016, 7, 84. [Google Scholar] [CrossRef] [PubMed]
- Lerner, A.; Aminov, R.; Matthias, T. Transglutaminases in Dysbiosis as Potential Environmental Drivers of Autoimmunity. Front. Microbiol. 2017, 8, 66. [Google Scholar] [CrossRef] [Green Version]
- Kieliszek, M.; Misiewicz, A. Microbial transglutaminase and its application in the food industry. A review. Folia Microbiol. 2014, 59, 241–250. [Google Scholar] [CrossRef] [Green Version]
- Duarte, L.; Matte, C.R.; Bizarro, C.V.; Ayub, M.A.Z. Transglutaminases: Part I—origins, sources, and biotechnological characteristics. World J. Microbiol. Biotechnol. 2020, 36, 15. [Google Scholar] [CrossRef]
- Martins, I.M.; Matos, M.; Costa, R.; Lopes-da-Silva, F.; Pascoal, A.; Estevinho, L.M.; Choupina, A.B. Transglutaminases: Recent achievements and new sources. Appl. Microbiol. Biotechnol. 2014, 98, 6957–6964. [Google Scholar] [CrossRef]
- Singh, R.; Kumar, M.; Mittal, A.; Mehta, P.K. Microbial enzymes: Industrial progress in 21st century. 3 Biotech 2016, 6, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Fatima, S.W.; Khare, S.K. Current insight and futuristic vistas of microbial transglutaminase in nutraceutical industry. Microbiol. Res. 2018, 215, 7–14. [Google Scholar] [CrossRef]
- Tokay, F.G.; Yerlikaya, P. Shelf-Life Extension of Fish Fillets by Spraying with Microbial Transglutaminase. J. Aquat. Food Prod. Technol. 2017, 26, 940–948. [Google Scholar] [CrossRef]
- Santos, M.; Torne, J. Recent Patents on Transglutaminase Production and Applications: A Brief Review. Recent Pat. Biotechnol. 2009, 3, 166–174. [Google Scholar] [CrossRef]
- Duarte, L.; Matte, C.R.; Bizarro, C.V.; Ayub, M.A.Z. Review transglutaminases: Part II—Industrial applications in food, biotechnology, textiles and leather products. World J. Microbiol. Biotechnol. 2020, 36, 11. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, R.G.; Karki, P.; Langleite, I.E.; Bakksjø, R.J.; Eichacker, L.A.; Furnes, C. Characterisation of a novel cold-adapted calcium-activated transglutaminase: Implications for medicine and food processing. FEBS Open Bio 2020, 10, 495–506. [Google Scholar] [CrossRef] [PubMed]
- Lerner, A.; Matthias, T. Possible association between celiac disease and bacterial transglutaminase in food processing: A hypothesis. Nutr. Rev. 2015, 73, 544–552. [Google Scholar] [CrossRef]
- Kaufmann, A.; Köppel, R.; Widmer, M. Determination of microbial transglutaminase in meat and meat products. Food Addit. Contam. Part. A 2012, 29, 1364–1373. [Google Scholar] [CrossRef]
- Yew, S.E.; Lim, T.J.; Lew, L.C.; Bhat, R.; Mat-Easa, A.; Liong, M.T. Development of a probiotic delivery system from agrowastes, soy protein isolate, and microbial transglutaminase. J. Food Sci. 2011, 76, H108–H115. [Google Scholar] [CrossRef]
- Lerner, A.; Matthias, T. Candida albicans in celiac disease: A wolf in sheep’s clothing. Autoimmun. Rev. 2020, 19, 102621. [Google Scholar]
- Iversen, R.; Amundsen, S.F.; Kleppa, L.; du Pré, M.F.; Stamnaes, J.; Sollid, L.M. Evidence That Pathogenic Transglutaminase 2 in Celiac Disease Derives from Enterocytes. Gastroenterology 2020, 159, 788–790. [Google Scholar] [CrossRef]
- Lerner, A.; Ramesh, A.; Matthias, T. Serologic Diagnosis of Celiac Disease: New Biomarkers. Gastroenterol. Clin. N. Am. 2019, 48, 307–317. [Google Scholar] [CrossRef]
- Agardh, D.; Matthias, T.; Wusterhausen, P.; Neidhöfer, S.; Heller, A.; Lerner, A. Antibodies against neo-epitope of microbial and human transglutaminase complexes as biomarkers of childhood celiac disease. Clin. Exp. Immunol. 2019, 199, 294–302. [Google Scholar] [CrossRef]
- Matthias, T.; Jeremias, P.; Neidhöfer, S.; Lerner, A. The industrial food additive, microbial transglutaminase, mimics tissue transglutaminase and is immunogenic in celiac disease patients. Autoimmun. Rev. 2016, 15, 1111–1119. [Google Scholar] [CrossRef]
- Lerner, A.; Jeremias, P.; Neidhöfer, S.; Matthias, T. Antibodies against neo-epitope tTg complexed to gliadin are different and more reliable then anti-tTg for the diagnosis of pediatric celiac disease. J. Immunol. Methods 2016, 429, 15–20. [Google Scholar] [CrossRef] [Green Version]
- Lerner, A.; Jeremias, P.; Neidhöfer, S.; Matthias, T. Comparison of the Reliability of 17 Celiac Disease Associated Bio-Markers to Reflect Intestinal Damage. J. Clin. Cell. Immunol. 2017, 8, 486. [Google Scholar] [CrossRef] [Green Version]
- Rozenberg, O.; Lerner, A.; Pacht, A.; Grinberg, M.; Reginashvili, D.; Henig, C.; Barak, M. A novel algorithm for the diagnosis of celiac disease and a comprehensive review of celiac disease diagnostics. Clin. Rev. Allergy Immunol. 2011, 42, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Rozenberg, O.; Lerner, A.; Pacht, A.; Grinberg, M.; Reginashvili, D.; Henig, C.; Barak, M. A new algorithm for the diagnosis of celiac disease. Cell. Mol. Immunol. 2011, 8, 146–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berti, C.; Roncoroni, L.; Falini, M.L.; Caramanico, R.; Dolfini, E.; Bardella, M.T.; Elli, L.; Terrani, C.; Forlani, F. Celiac-related properties of chemically and enzymatically modified gluten proteins. J. Agric. Food Chem. 2007, 55, 2482–2488. [Google Scholar] [CrossRef]
- Cabrera-CháVez, F.; Rouzaud-Sández, O.; Sotelo-Cruz, N.; Calderón de la Barca, A.M. Bovine milk caseins and transglutaminase-treated cereal prolamins are differentially recognized by iga of celiac disease patients according to their age. J. Agric. Food Chem. 2009, 57, 3754–3759. [Google Scholar] [CrossRef] [PubMed]
- Elli, L.; Roncoroni, L.; Hils, M.; Pasternack, R.; Barisani, D.; Terrani, C.; Vaira, V.; Ferrero, S.; Bardella, M.T. Immunological effects of transglutaminase-treated gluten in coeliac disease. Hum. Immunol. 2012, 73, 992–997. [Google Scholar] [CrossRef] [PubMed]
- Cabrera-Chávez, F.; Rouzaud-Sández, O.; Sotelo-Cruz, N.; Calderón De La Barca, A.M. Transglutaminase treatment of wheat and maize prolamins of bread increases the serum IgA reactivity of celiac disease patients. J. Agric. Food Chem. 2008, 56, 1387–1391. [Google Scholar] [CrossRef] [PubMed]
- Falini, M.L.; Elli, L.; Caramanico, R.; Bardella, M.T.; Terrani, C.; Roncoroni, L.; Doneda, L.; Forlani, F. Immunoreactivity of antibodies against transglutaminase-deamidated gliadins in adult celiac disease. Dig. Dis. Sci. 2008, 53, 2697–2701. [Google Scholar] [CrossRef]
- Heredia-Sandoval, N.G.; Islas-Rubio, A.R.; Cabrera-Chávez, F.; Calderón De La Barca, A.M. Transamidation of gluten proteins during the bread-making process of wheat flour to produce breads with less immunoreactive gluten. Food Funct. 2014, 5, 1813–1818. [Google Scholar] [CrossRef]
- Dekking, E.H.A.; Van Veelen, P.A.; de Ru, A.; Kooy-Winkelaar, E.M.C.; Gröneveld, T.; Nieuwenhuizen, W.F.; Koning, F. Microbial transglutaminases generate T cell stimulatory epitopes involved in celiac disease. J. Cereal Sci. 2008, 47, 339–346. [Google Scholar] [CrossRef]
- Luongo, D.; Maurano, F.; Bergamo, P.; Rossi, M. Microbial transglutaminase: A biotechnological tool to manage gluten intolerance. Anal. Biochem. 2020, 592, 113584. [Google Scholar] [CrossRef]
- Luongo, D.; Bonavita, R.; Rossi, S.; Rotondi Aufiero, V.; Feliciello, N.R.; Maurano, F.; Gaetano Iaquinto, G.; Mazzarella, G.; Rossi, M. Tailoring the immune response to wheat gliadin by enzymatic transamidation. Cytokine 2019, 117, 23–29. [Google Scholar] [CrossRef]
- Zhou, L.; Kooy-Winkelaar, Y.M.C.; Cordfunke, R.A.; Dragan, I.; Thompson, A.; Drijfhout, J.W.; van Veelen, P.A.; Chen, H.; Koning, F. Abrogation of Immunogenic Properties of Gliadin Peptides through Transamidation by Microbial Transglutaminase Is Acyl-Acceptor Dependent. J. Agric. Food Chem. 2017, 65, 7542–7552. [Google Scholar] [CrossRef]
- Heil, A.; Ohsam, J.; Van Genugten, B.; Diez, O.; Yokoyama, K.; Kumazawa, Y.; Pasternack, R.; Hils, M. Microbial Transglutaminase Used in Bread Preparation at Standard Bakery Concentrations Does Not Increase Immunodetectable Amounts of Deamidated Gliadin. J. Agric. Food Chem. 2017, 65, 6982–6990. [Google Scholar] [CrossRef]
- Lerner, A.; Matthias, T. Food Industrial Microbial Transglutaminase in Celiac Disease: Treat or Trick. Int. J. Celiac Dis. 2015, 3, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Stricker, S.; De Laffolie, J.; Rudloff, S.; Komorowski, L.; Zimmer, K.P. Intracellular localization of microbial transglutaminase and its influence on the transport of gliadin in enterocytes. J. Pediatr. Gastroenterol. Nutr. 2019, 68, e43–e50. [Google Scholar] [CrossRef]
- Chen, L.; Ullah, N.; Li, C.; Hackman, R.M.; Li, Z.; Xu, X.; Guanghong Zhou, G.; Feng, X. Incorporated glucosamine adversely affects the emulsifying properties of whey protein isolate polymerized by transglutaminase. J. Dairy Sci. 2017, 100, 3413–3423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, X.; Ren, J.; Zhao, M.; Cui, C.; He, P. Emulsifying properties of the transglutaminase-treated crosslinked product between peanut protein and fish (Decapterus maruadsi) protein hydrolysates. J. Sci. Food Agric. 2010, 91, 578–585. [Google Scholar] [CrossRef]
- Li, F.; Lu, J.; Kong, X.; Hyeon, T.; Ling, D. Dynamic Nanoparticle Assemblies for Biomedical Applications. Adv. Mater. 2017, 29, 1605897. [Google Scholar] [CrossRef]
- Singh, T.; Shukla, S.; Kumar, P.; Wahla, V.; Bajpai, V.K. Application of nanotechnology in food science: Perception and overview. Front. Microbiol. 2017, 8, 1501. [Google Scholar] [CrossRef] [Green Version]
- Faust, J.J.; Masserano, B.M.; Mielke, A.H.; Abraham, A.; Capco, D.G. Engineered nanoparticles induced brush border disruption in a human model of the intestinal epithelium. Adv. Exp. Med. Biol. 2014, 811, 55–72. [Google Scholar] [PubMed]
- Lama, S.; Merlin-Zhang, O.; Yang, C. In vitro and in vivo models for evaluating the oral toxicity of nanomedicines. Nanomaterials 2020, 10, 2177. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.H.; Tang, M.Z.; Yu, X.T.; Xu, C.M.; Yang, H.M.; Tang, J.B. Site-specific, covalent immobilization of an engineered enterokinase onto magnetic nanoparticles through transglutaminase-catalyzed bioconjugation. Colloids Surf. B Biointerfaces 2019, 177, 506–511. [Google Scholar] [CrossRef]
- Ma, T.; Lu, J.; Zhu, J.; Li, X.; Gu, H.; Montalbán-López, M.; Wu, X.; Luo, S.; Zhao, Y.; Jiang, S.; et al. The Secretion of Streptomyces monbaraensis Transglutaminase from Lactococcus lactis and Immobilization on Porous Magnetic Nanoparticles. Front Microbiol. 2019, 10, 1675. [Google Scholar] [CrossRef] [Green Version]
- Fasano, A. Zonulin and its regulation of intestinal barrier function: The biological door to inflammation, autoimmunity, and cancer. Physiol. Rev. 2011, 91, 151–175. [Google Scholar] [CrossRef] [Green Version]
- Sanders, D.J.; Inniss, S.; Sebepos-Rogers, G.; Rahman, F.Z.; Smith, A.M. The role of the microbiome in gastrointestinal inflammation. Biosci. Rep. 2021, 41, BSR20203850. [Google Scholar] [CrossRef] [PubMed]
- Hollon, J.; Puppa, E.L.; Greenwald, B.; Goldberg, E.; Guerrerio, A.; Fasano, A. Effect of gliadin on permeability of intestinal biopsy explants from celiac disease patients and patients with Non-Celiac gluten sensitivity. Nutrients 2015, 7, 1565–1576. [Google Scholar] [CrossRef] [PubMed]
- Cardoso-Silva, D.; Delbue, D.; Itzlinger, A.; Moerkens, R.; Withoff, S.; Branchi, F.; Schumann, M. Intestinal barrier function in gluten-related disorders. Nutrients 2019, 11, 2325. [Google Scholar] [CrossRef] [Green Version]
- Kumar, M.D.; Singh, A.K.; Shama, H.; Deshwal, G.K. Histone cross-linking by transglutaminase. Biochem. Biophys. Res. Commun. 2002, 293, 1453–1457. [Google Scholar]
- Dieli-Crimi, R.; Cénit, M.C.; Núñez, C. The genetics of celiac disease: A comprehensive review of clinical implications. J. Autoimmun. 2015, 64, 26–41. [Google Scholar] [CrossRef]
- Perry, A.S.; Baird, A.M.; Gray, S.G. Epigenetic methodologies for the study of celiac disease. Methods Mol. Biol. 2015, 1326, 131–158. [Google Scholar]
- Miyoshi, Y.; Tanabe, S.; Suzuki, T. Cellular zinc is required for intestinal epithelial barrier maintenance via the regulation of claudin-3 and occludin expression. Am. J. Physiol. Liver Physiol. 2016, 311, G105–G116. [Google Scholar] [CrossRef] [Green Version]
- Lerner, A.; Neidhöfer, S.; Matthias, T. The Gut Microbiome Feelings of the Brain: A Perspective for Non-Microbiologists. Microorganisms 2017, 5, 66. [Google Scholar] [CrossRef]
- Boukhettala, N.; Claeyssens, S.; Bensifi, M.; Maurer, B.; Abed, J.; Lavoinne, A.; Déchelotte, P.; Coëffier, M. Effects of essential amino acids or glutamine deprivation on intestinal permeability and protein synthesis in HCT-8 cells: Involvement of GCN2 and mTOR pathways. Amino Acids 2012, 42, 375–383. [Google Scholar] [CrossRef]
- Obrenovich, M. Leaky Gut, Leaky Brain? Microorganisms 2018, 6, 107. [Google Scholar] [CrossRef] [Green Version]
- Lerner, A.; Makhoul, B.F.; Eliakim, R. Neurological Manifestations of Celiac Disease in Children and Adults Affiliations Celiac disease and environment View project Neurological Manifestations of Celiac Disease in Children and Adults. Eur. Neurol. J. 2012, 4, 15–20. [Google Scholar]
- Zelnik, N.; Pacht, A.; Obeid, R.; Lerner, A. Range of neurologic disorders in patients with celiac disease. Pediatrics 2004, 113, 1672–1676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vojdani, A.; Lerner, A.; Vojdani, E. Cross-Reactivity and Sequence Homology between Al-Pha-Synuclein and Food Products: A Step Further for Parkinson’s Disease Synucleinopathy. Cells 2021, 10, 1111. [Google Scholar] [CrossRef]
- Lerner, A.; Sobolevskaia, P.; Churilov, L.; Shoenfeld, Y. Alpha-enolase involvement in intestinal and extraintestinal manifestations of celiac disease. J. Transl. Autoimmun. 2021, 4, 100109. [Google Scholar] [CrossRef]
- Gatta, N.G.; Cammarota, G.; Iannaccone, M.; Gentile, V. Transglutaminase Activity as a Possible Molecular Mechanism in the Etiopathogenesis of Neurodegenerative Diseases. J. Biochem. Mol. Biol. Res. 2016, 2, 157–165. [Google Scholar] [CrossRef] [Green Version]
- Lebreton, C.; Ménard, S.; Abed, J.; Moura, I.C.; Coppo, R.; Dugave, C.; Monteiro, R.C.; Fricot, A.; Traore, M.G.; Martin Griffin, M. Interactions among secretory immunoglobulin A, CD71, and transglutaminase-2 affect permeability of intestinal epithelial cells to gliadin peptides. Gastroenterology 2012, 143, 698–707.e4. [Google Scholar] [CrossRef]
- Recktenwald, C.V.; Hansson, G.C. The reduction-insensitive bonds of the MUC2 mucin are isopeptide bonds. J. Biol. Chem. 2016, 291, 13580–13590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pian, Y.; Wang, P.; Liu, P.; Zheng, Y.; Zhu, L.; Wang, H.; Xu, B.; Yuan, Y.; Jiang, Y. Proteomics identification of novel fibrinogen-binding proteins of Streptococcus suis contributing to antiphagocytosis. Front. Cell. Infect. Microbiol. 2015, 5, 19. [Google Scholar] [CrossRef] [Green Version]
- Xu, B.; Zhang, P.; Li, W.; Liu, R.; Tang, J.; Fan, H. hsdS, belonging to the type i restriction-modification system, contributes to the streptococcus suis serotype 2 survival ability in phagocytes. Front. Microbiol. 2017, 8, 1524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, R.Y.; Chen, J.; Li, Y. Heterologous leaky production of transglutaminase in Lactococcus lactis significantly enhances the growth performance of the host. Appl. Environ. Microbiol. 2005, 71, 8481–8490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, R.Y.; Chen, J.; Li, Y. Influence of expression of transglutaminase on the growth of Lactococcus lactis. Wei Sheng Wu Xue Bao 2005, 45, 510–515. [Google Scholar]
- Zhang, N.; Zhang, S.; He, Y.; Chen, X.; Zhang, Y.; Dong, Z. Intein-mediated intracellular production of active microbial transglutaminase in Corynebacterium glutamicum. Enzym. Microb. Technol. 2020, 142, 109680. [Google Scholar] [CrossRef] [PubMed]
- Rickert, M.; Strop, P.; Lui, V.; Melton-Witt, J.; Farias, S.E.; Foletti, D.; Shelton, D.; Jaume Pons, J.; Rajpal, A. Production of soluble and active microbial transglutaminase in Escherichia coli for site-specific antibody drug conjugation. Protein Sci. 2016, 25, 442–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lerner, A.; Matthias, T.; Aminov, R. Potential effects of horizontal gene exchange in the human gut. Front. Immunol. 2017, 8, 1630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mann, E.R.; Li, X. Intestinal antigen-presenting cells in mucosal immune homeostasis: Crosstalk between dendritic cells, macrophages and B-cells. World J Gastroenterol. 2014, 20, 9653–9664. [Google Scholar] [CrossRef] [PubMed]
- Stagg, A.J. Intestinal Dendritic Cells in Health and Gut Inflammation. Front. Immunol. 2018, 9, 2883. [Google Scholar] [CrossRef] [Green Version]
- Bekiaris, V.; Persson, E.K.; Agace, W.W. Intestinal dendritic cells in the regulation of mucosal immunity. Immunol. Rev. 2014, 260, 86–101. [Google Scholar] [CrossRef]
- Chrobok, N.L.; Sestito, C.; Wilhelmus, M.M.M.; Drukarch, B.; van Dam, A.M. Is monocyte- and macrophage-derived tissue transglutaminase involved in inflammatory processes? Amino Acids 2017, 49, 441–452. [Google Scholar] [CrossRef] [Green Version]
- Ráki, M.; Schjetne, K.W.; Stamnaes, J.; Molberg Jahnsen, F.L.; Issekutz, T.B.; Bogen, B.; Sollid, L.M. Surface expression of transglutaminase 2 by dendritic cells and its potential role for uptake and presentation of gluten peptides to T cells. Scand. J. Immunol. 2007, 65, 213–220. [Google Scholar] [CrossRef]
- Hodrea, J.; Demény, M.Á.; Majai, G.; Sarang, Z.; Korponay-Szabó, I.R.; Fésüs, L. Transglutaminase 2 is expressed and active on the surface of human monocyte-derived dendritic cells and macrophages. Immunol. Lett. 2010, 130, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Chander, A.M.; Yadav, H.; Jain, S.; Bhadada, S.K.; Dhawan, D.K. Cross-talk between gluten, intestinal microbiota and intestinal mucosa in celiac disease: Recent advances and basis of autoimmunity. Front. Microbiol. 2018, 9, 2597. [Google Scholar] [CrossRef] [PubMed]
- Lerner, A.; Ramesh, A.; Matthias, T. The temperature and pH repertoire of the transglutaminase family is expanding. FEBS Open Bio 2020, 10, 492–494. [Google Scholar] [CrossRef]
- Deckers, M.; Vanneste, K.; Winand, R.; Keersmaecker, S.C.J.D.; Denayer, S.; Heyndrickx, M.; Deforce, D.; Fraiture, M.A.; Roosens, N.H.C. Strategy for the identification of micro-organisms producing food and feed products: Bacteria producing food enzymes as study case. Food Chem. 2020, 305, 125431. [Google Scholar] [CrossRef] [Green Version]
- Fraiture, M.A.; Deckers, M.; Papazova, N.; Roosens, N.H.C. Are antimicrobial resistance genes key targets to detect genetically modified microorganisms in fermentation products? Int. J. Food Microbiol. 2020, 331, 108749. [Google Scholar] [CrossRef]
- Cheng, G.; Ning, J.; Ahmed, S.; Huang, J.; Ullah, R.; An, B.; Hao, H.; Dai, M.; Huang, L.; Wang, X.; et al. Selection and dissemination of antimicrobial resistance in Agri-food production. Antimicrob. Resist. Infect. Control. 2019, 8, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Paul, R.H.; Frestedt, J.; Magurany, K. GRAS from the ground up: Review of the Interim Pilot Program for GRAS notification. Food Chem Toxicol. 2017, 105, 140–150. [Google Scholar] [CrossRef]
- Burdock, G.A.; Carabin, I.G. Generally recognized as safe (GRAS): History and description. Toxicol. Lett. 2004, 150, 3–18. [Google Scholar] [CrossRef]
- Hallagan, J.B.; Hall, R.L.; Drake, J. The GRAS provision—The FEMA GRAS program and the safety and regulation of flavors in the United States. Food Chem. Toxicol. 2020, 138, 111236. [Google Scholar] [CrossRef] [PubMed]
- Faustman, C.; Aaron, D.; Negowetti, N.; Leib, E.B. Ten years post-GAO assessment, FDA remains uninformed of potentially harmful GRAS substances in foods. Crit. Rev. Food Sci. Nutr. 2021, 61, 1260–1268. [Google Scholar] [CrossRef]
- Sewalt, V.; LaMarta, J.; Shanahan, D.; Gregg, L.; Carrillo, R. Letter to the editor regarding “GRAS from the ground up: Review of the Interim Pilot Program for GRAS notification” by Hanlon et al., 2017. Food Chem. Toxicol. 2017, 107, 520–521. [Google Scholar] [CrossRef]
- Kruger, C. The relevance of international assessments to GRAS determinations. Regul. Toxicol. Pharmacol. 2016, 79, S119–S123. [Google Scholar] [CrossRef] [PubMed]
- Neltner, T.G.; Alger, H.M.; O’Reilly, J.T.; Krimsky, S.; Bero, L.A.; Maffini, M.V. Conflicts of interest in approvals of additives to food: Determined to be generally recognized as safe: Out of balance. JAMA Intern. Med. 2013, 173, 2032–2036. [Google Scholar] [CrossRef] [PubMed]
- Roberts, A.; Haighton, L.A. A hard look at FDA’s review of GRAS notices. Regul. Toxicol. Pharmacol. 2016, 79, S124–S128. [Google Scholar] [CrossRef]
- Williams, G.M.; Kobets, T.; Iatropoulos, M.J.; Duan, J.D.; Brunnemann, K.D. GRAS determination scientific procedures and possible alternatives. Regul. Toxicol. Pharmacol. 2016, 79, S105–S111. [Google Scholar] [CrossRef]
- Trasande, L.; Shaffer, R.M.; Sathyanarayana, S. Council on Environmental Health. Food Additives and Child Health. Pediatrics 2018, 142, e20181408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sander, I.; Keller, C.; Czibor, C.; Meurer, U.; Merget, R.; Raulf, M. Unusual allergen in a butcher with respiratory symptoms. Allergol. Sel. 2020, 4, 105–109. [Google Scholar] [CrossRef] [PubMed]
- De Palma, G.; Apostoli, P.; Mistrello, G.; Zanotta, S.; Bertorelli, G. Microbial transglutaminase: A new and emerging occupational allergen. Ann. Allergy Asthma Immunol. 2014, 112, 553–554. [Google Scholar] [CrossRef]
- Zhao, L.; Li, X.; Zhao, J.; Ma, S.; Ma, X.; Fan, D.; Zhu, C.; Liu, Y. A novel smart injectable hydrogel prepared by microbial transglutaminase and human-like collagen: Its characterization and biocompatibility. Mater. Sci. Eng. C 2016, 68, 317–326. [Google Scholar] [CrossRef]
- De Oliveira, G.L.V.; Cardoso CR de, B.; Taneja, V.; Fasano, A. Editorial: Intestinal Dysbiosis in Inflammatory Diseases. Front. Immunol. 2021, 12, 727485. [Google Scholar] [CrossRef]
- Malandain, H. Transglutaminases: A meeting point for wheat allergy, celiac disease, and food safety. Allerg. Immunol. 2005, 37, 397–403. [Google Scholar]
- Skovbjerg, H.; Norén, O.; Anthonsen, D.; Moller, J.; Sjöström, H. Gliadin is a good substrate of several transglutaminases: Possible implication in the pathogenesis of coeliac disease. Scand. J. Gastroenterol. 2002, 37, 812–817. [Google Scholar] [CrossRef] [PubMed]
- Gerrard, J.A.; Sutton, K.H. Addition of transglutaminase to cereal products may generate the epitope responsible for coeliac disease. Trends Food Sci. Technol. 2005, 16, 510–512. [Google Scholar] [CrossRef]
- Gerrard, J.A.; Cottam, J.R. Protein Cross-linking in Food—Structure, Applications, Implications for Health and Food Safety. In Food Biochemistry and Food Processing; Wiley: Hoboken, NJ, USA, 2012; pp. 207–222. [Google Scholar]
- Kumar, M.D.; Singh, A.K.; Sharma, H.; Deshwal, G.K. Promising Scope of Transglutaminase as Processing Aid in Food Industries. Food Sci. Rep. 2020, 1, 53–55. [Google Scholar]
- SR 817.022.108—Verordnung des EDI vom 23. November 2005 Über Lebensmittel Tierischer Herkunft [Internet]. Available online: https://www.fedlex.admin.ch/eli/cc/2005/801/de (accessed on 23 August 2021).
- Darloman, I. Microbial Transglutaminase: A Review on Current Concerning Aspects [Internet]. [Barcelona]: Universitat Autònoma de Barcelona. 2018. Available online: https://ddd.uab.cat/pub/trerecpro/2018/hdl_2072_365412/TFM_idarloman.pdf (accessed on 23 August 2021).
- Sutay Kocabaş, D.; Grumet, R. Evolving regulatory policies regarding food enzymes produced by recombinant microorganisms. GM Crop. Food 2019, 10, 191–207. [Google Scholar] [CrossRef] [PubMed]
- Akbari, M.; Razavi, S.H.; Kieliszek, M. Recent advances in microbial transglutaminase biosynthesis and its application in the food industry. Trends Food Sci. Technol. 2021, 110, 458–469. [Google Scholar] [CrossRef]
- Ziberna, F.; Sblattero, D.; Lega, S.; Stefani, C.; Dal Ferro, M.; Marano, F.; Gaita, B.; De Leo, L.; Vatta, S.; Berti, L.; et al. A novel quantitative ELISA as accurate and reproducible tool to detect epidermal transglutaminase antibodies in patients with Dermatitis Herpetiformis. J. Eur. Acad. Dermatol. Venereol. 2021, 35, e78–e80. [Google Scholar] [CrossRef] [PubMed]
- Amirdivani, S.; Khorshidian, N.; Fidelis, M.; Granato, D.; Koushki, M.R.; Mohammadi, M.; Khoshtinat, K.; Mortazavian, A.M. Effects of transglutaminase on health properties of food products. Curr. Opin. Food Sci. 2018, 22, 74–80. [Google Scholar] [CrossRef]
- Vojdani, A.; Vojdani, E. Amyloid-Beta 1-42 Cross-Reactive Antibody Prevalent in Human Sera May Contribute to Intraneuronal Deposition of A-Beta-P-42. Int. J. Alzheimers Dis. 2018, 2018, 1672568. [Google Scholar] [CrossRef] [Green Version]
- Karpuj, M.V.; Becher, M.W.; Steinman, L. Evidence for a role for transglutaminase in Huntington’s disease and the potential therapeutic implications. Neurochem. Int. 2002, 40, 31–36. [Google Scholar] [CrossRef]
- Osman, D.; Umar, S.; Muhammad, H.; Nikfekr, E.; Rostami, K.; Ishaq, S. Neurological manifestation of coeliac disease with particular emphasis on gluten ataxia and immunological injury: A review article. Gastroenterol. Hepatol. Bed Bench. 2021, 14, 1. [Google Scholar] [PubMed]
- Vojdani, E.; Vojdani, A. Reaction of antibodies to Campylobacter jejuni and cytolethal distending toxin B with tissues and food antigens. World J. Gastroenterol. 2019, 25, 1050–1066. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Human Disease | References |
|---|---|
| Celiac disease | [3,9,10,11,12,13,17,18,28,33,34,35,37,38,39,41,42,43,44,46,51,52,112,113] |
| Dermatitis herpetiformis | [120] |
| Neurodegenerative diseases | [14,15,76,121] |
| Parkinson’s disease | [74] |
| Alzheimer’s disease | [121,122] |
| Huntington disease | [121,123] |
| Gluten ataxia | [124] |
| Allergic diseases | [107,108] |
| Campylobacter jejuni associated diseases | [125] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lerner, A.; Benzvi, C. Microbial Transglutaminase Is a Very Frequently Used Food Additive and Is a Potential Inducer of Autoimmune/Neurodegenerative Diseases. Toxics 2021, 9, 233. https://doi.org/10.3390/toxics9100233
Lerner A, Benzvi C. Microbial Transglutaminase Is a Very Frequently Used Food Additive and Is a Potential Inducer of Autoimmune/Neurodegenerative Diseases. Toxics. 2021; 9(10):233. https://doi.org/10.3390/toxics9100233
Chicago/Turabian StyleLerner, Aaron, and Carina Benzvi. 2021. "Microbial Transglutaminase Is a Very Frequently Used Food Additive and Is a Potential Inducer of Autoimmune/Neurodegenerative Diseases" Toxics 9, no. 10: 233. https://doi.org/10.3390/toxics9100233
APA StyleLerner, A., & Benzvi, C. (2021). Microbial Transglutaminase Is a Very Frequently Used Food Additive and Is a Potential Inducer of Autoimmune/Neurodegenerative Diseases. Toxics, 9(10), 233. https://doi.org/10.3390/toxics9100233