
High-Throughput Screening of Psychotropic Compounds: Impacts on Swimming Behaviours in Artemia franciscana



Abstract
:1. Introduction
2. Materials and Methods
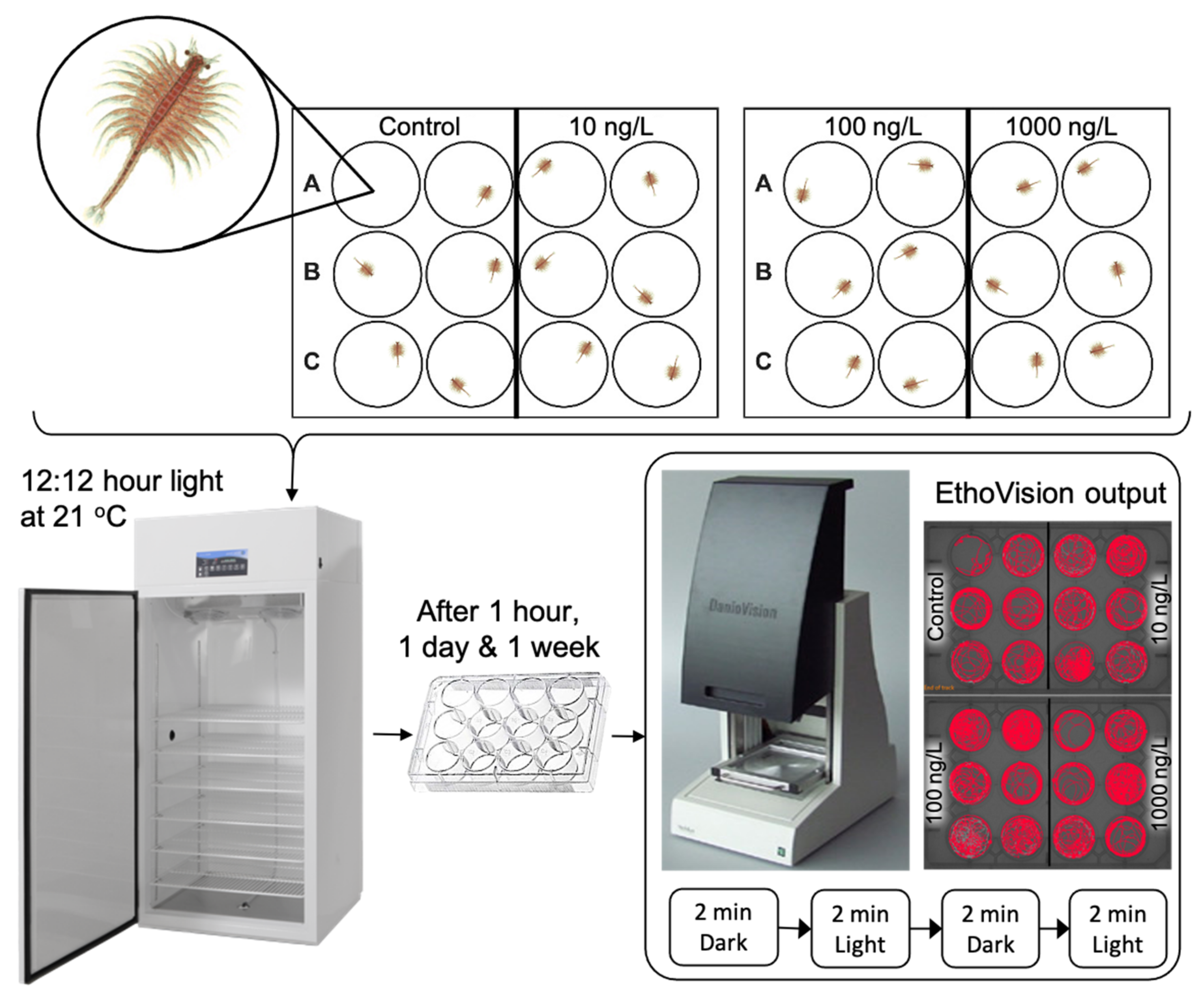
2.1. Animal Culture and Husbandry
2.2. Measuring Behaviour
2.3. Baseline Behaviours
2.4. Velocity
Phototaxis
2.5. Psychotropic Exposures
Preparation of Solutions
2.6. Exposures and Behavioural Analysis
2.7. Statistics
3. Results
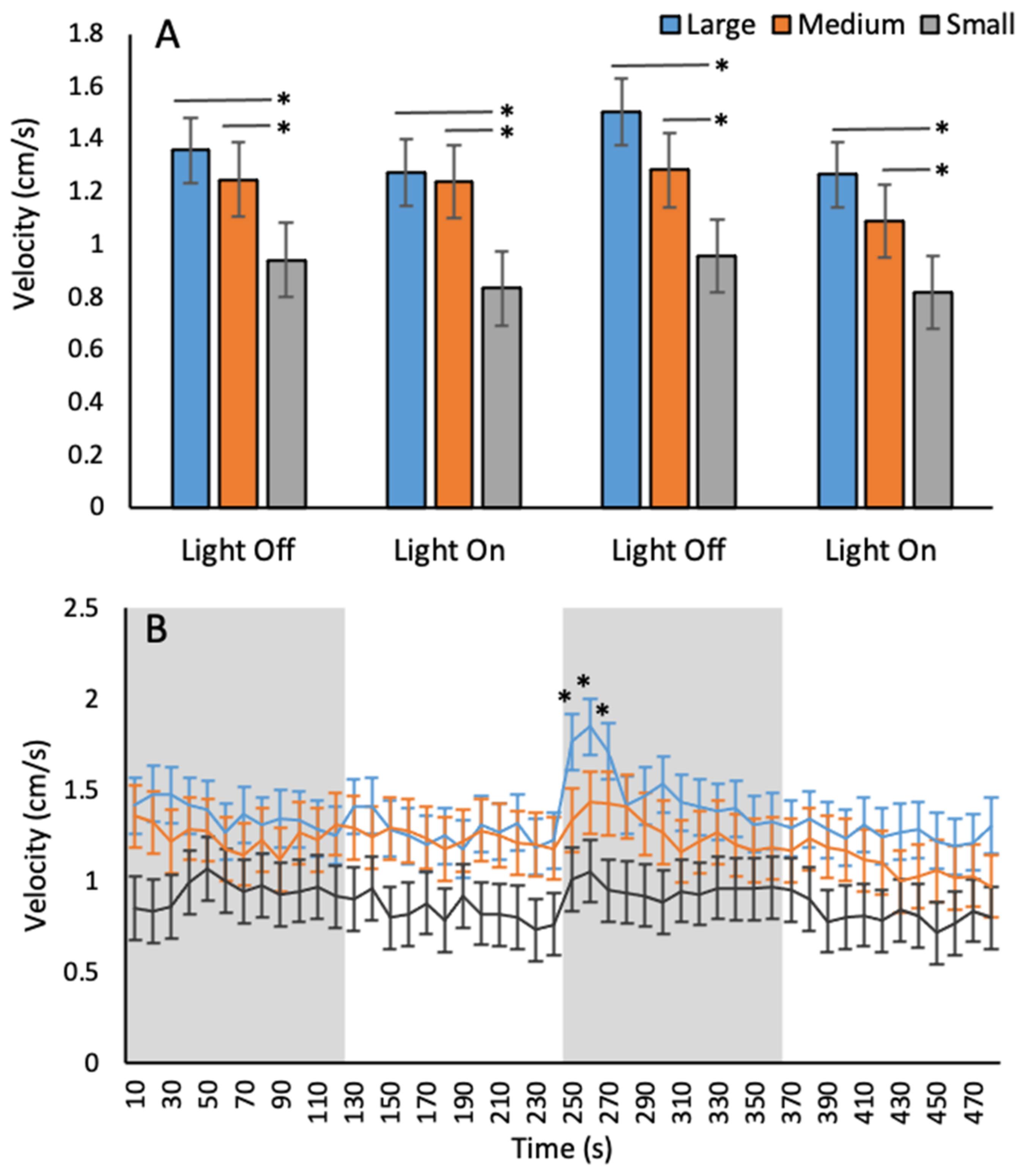
3.1. Baseline Behaviours
Velocity
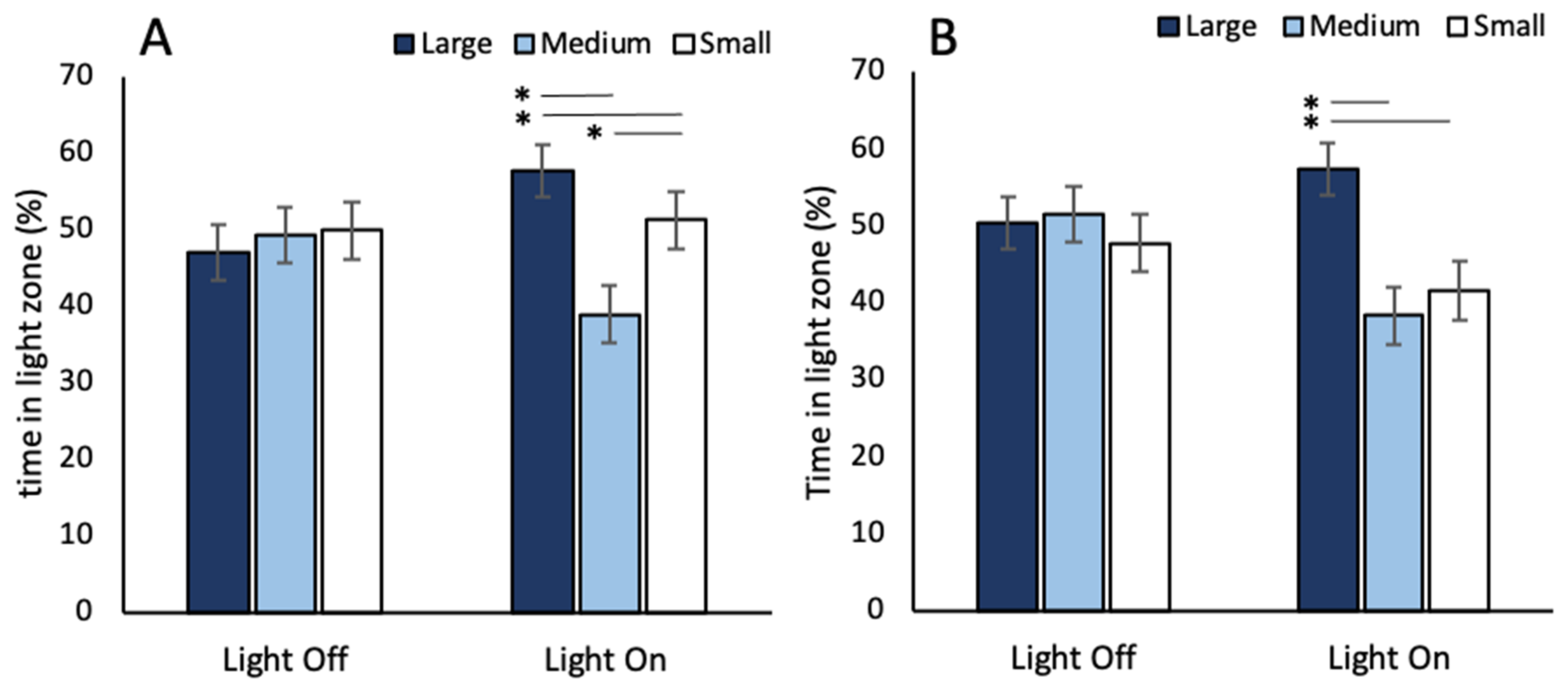
3.2. Phototaxis
3.3. Psychotropic Exposures
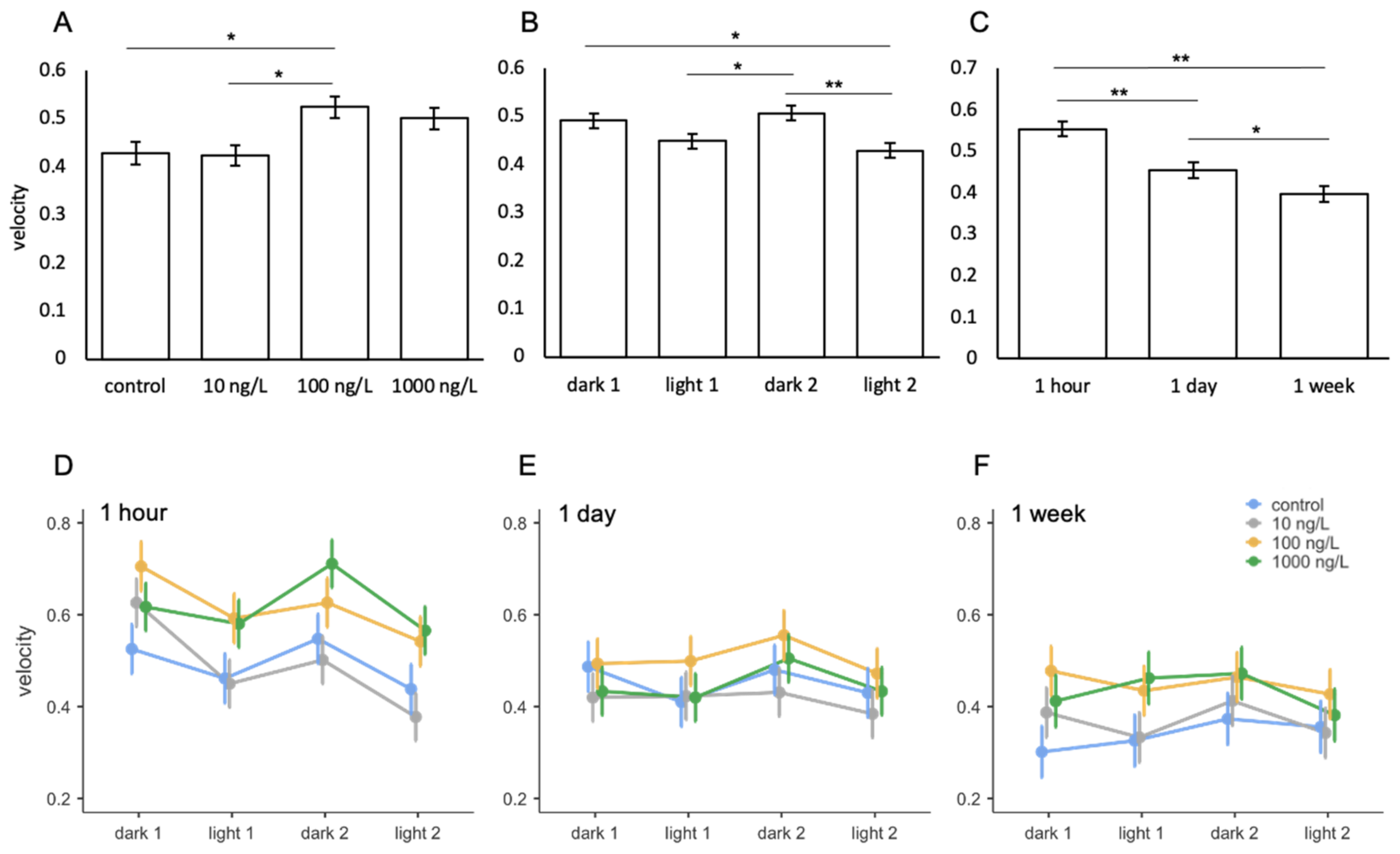
Fluoxetine
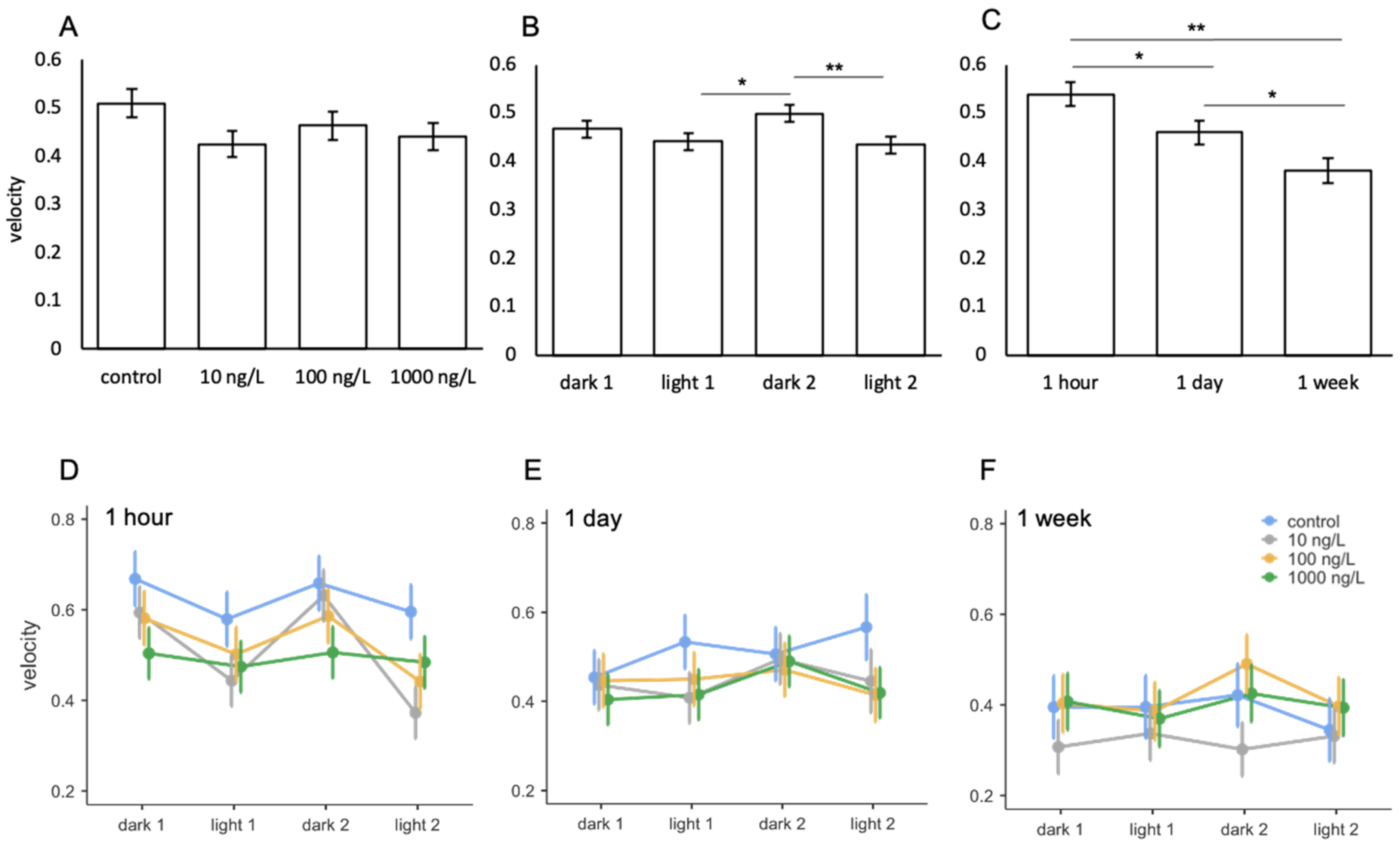
3.4. Oxazepam
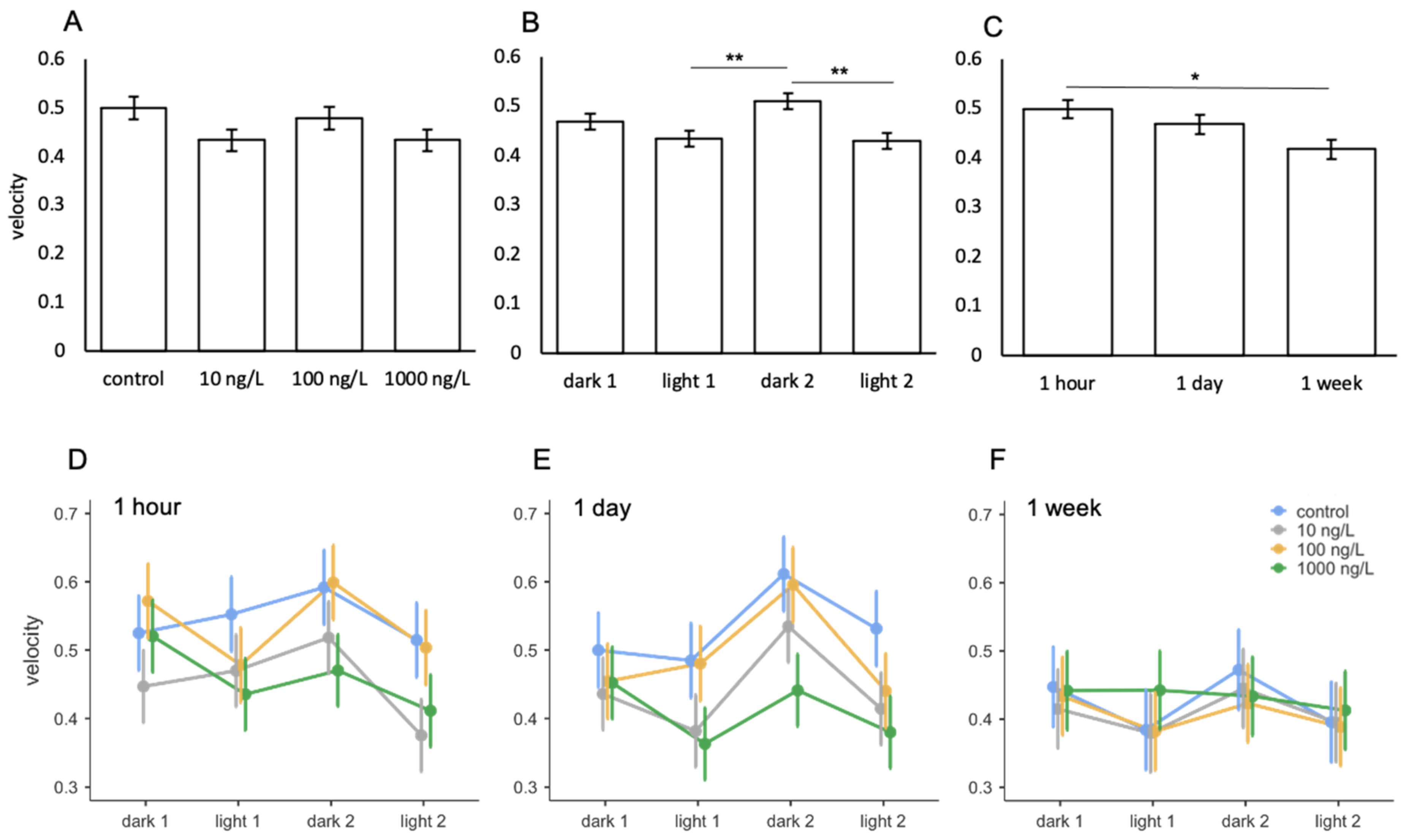
3.5. Amitriptyline
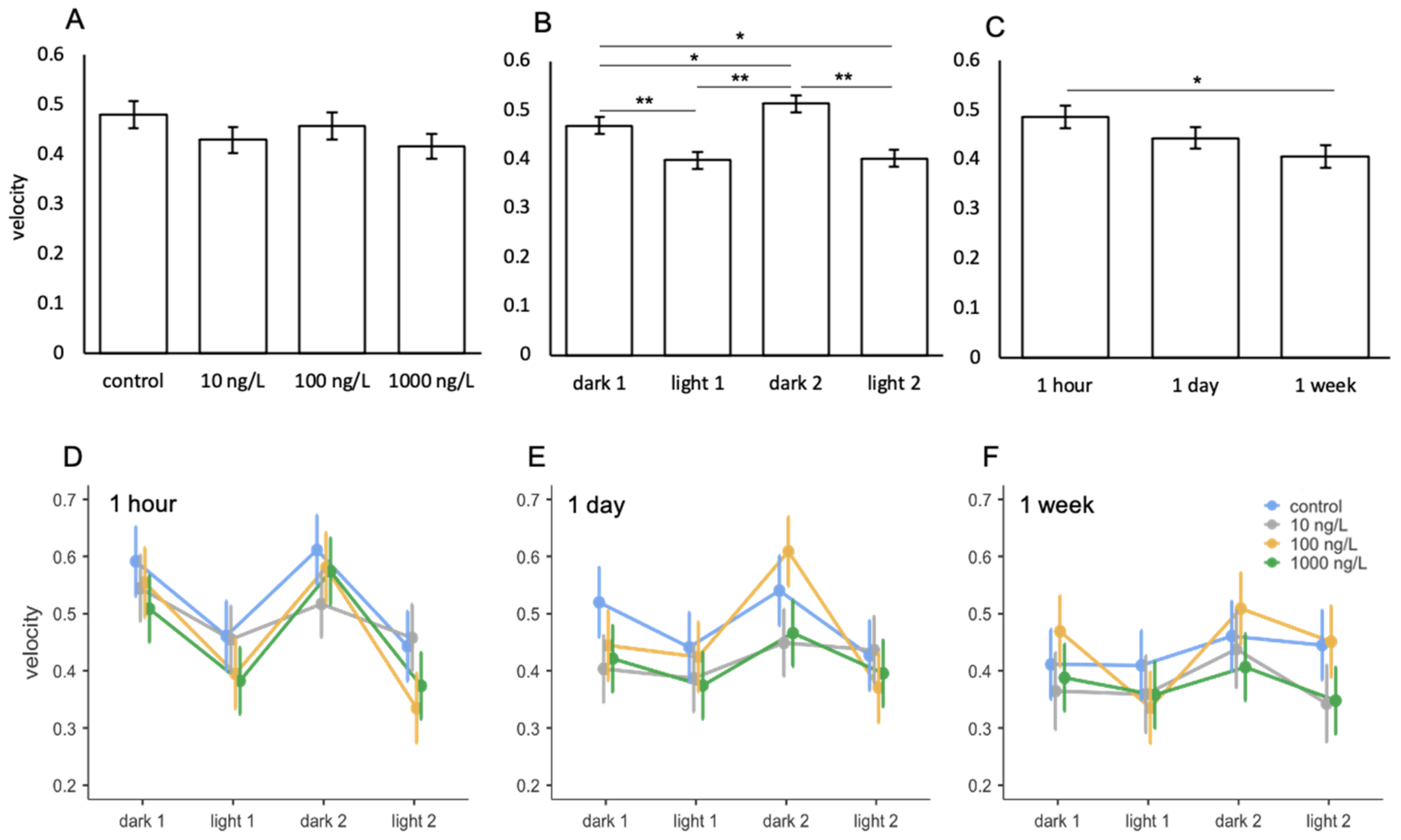
3.6. Venlafaxine
4. Discussion
4.1. Baseline Behaviours
4.2. Psychotropic Exposures
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Simão, F.C.P.; Martínez-Jerónimo, F.; Blasco, V.; Moreno, F.; Porta, J.M.; Pestana, J.L.T.; Soares, A.M.V.M.; Raldúa, D.; Barata, C. Using a new high-throughput video-tracking platform to assess behavioural changes in Daphnia magna exposed to neuro-active drugs. Sci. Total Environ. 2019, 662, 160–167. [Google Scholar] [CrossRef] [Green Version]
- Melvin, S.D.; Wilson, S.P. The utility of behavioral studies for aquatic toxicology testing: A meta-analysis. Chemosphere 2013, 93, 2217–2223. [Google Scholar] [CrossRef]
- Parker, M.O. Adult vertebrate behavioural aquatic toxicology: Reliability and validity. Aquat. Toxicol. 2016, 170, 323–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pyle, G.G.; Ford, A.T. Behaviour revised: Contaminant effects on aquatic animal behaviour. Aquat. Toxicol. 2016, 182, 226–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wlodkowic, D.; Khoshmanesh, K.; Akagi, J.; Williams, D.E.; Cooper, J.M. Wormometry-on-a-chip: Innovative technologies for In Situ Analysis of Small Multicellular Organisms. Cytom. Part. A 2011, 79, 799–813. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Persoone, G.; Nugegoda, D.; Wlodkowic, D. Enabling sub-lethal behavioral ecotoxicity biotests using microfluidic Lab-on-a-Chip technology. Sens. Actuators B Chem. 2016, 226, 289–298. [Google Scholar] [CrossRef]
- Cartlidge, R.; Nugegoda, D.; Wlodkowic, D. Millifluidic Lab-on-a-Chip technology for automated toxicity tests using the marine amphipod Allorchestes compressa. Sens. Actuators 2017, 239, 660–670. [Google Scholar] [CrossRef]
- Richendrfer, H.; Pelkowski, S.D.; Colwill, R.M.; Creton, R. On the edge: Pharmacological evidence for anxiety-related behavior in zebrafish larvae. Behav. Brain Res. 2012, 228, 99–106. [Google Scholar] [CrossRef] [Green Version]
- Schnörr, S.J.; Steenbergen, P.J.; Richardson, M.K.; Champagne, D.L. Measuring thigmotaxis in larval zebrafish. Behav. Brain Res. 2012, 228, 367–374. [Google Scholar] [CrossRef] [PubMed]
- Kohler, S.A.; Parker, M.O.; Ford, A.T. Shape and size of the arenas affect amphipod behaviours: Implications for ecotoxicology. PeerJ 2018, 6, e5271. [Google Scholar] [CrossRef]
- Kohler, S.A.; Parker, M.O.; Ford, A.T. Species-specific behaviours in amphipods highlight the need for understanding baseline behaviours in ecotoxicology. Aquat. Toxicol. 2018, 202, 173–180. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Yu, J. A Well-Established Method for the Rapid Assessment of Toxicity Using Artemia spp. Model. In Assessment and Management of Radioactive and Electronic Wastes; IntechOpen: London, UK, 2019; pp. 1–15. [Google Scholar] [CrossRef] [Green Version]
- Persoone, G.; Wells, P. Artemia in aquatic toxicology: A review. Artemia Res. Appl. 1987, 1, 259–275. [Google Scholar]
- Faimali, M.; Garaventa, F.; Piazza, V.; Greco, G.; Corrà, C.; Magillo, F.; Pittore, M.; Giacco, E.; Gallus, L.; Falugi, C.; et al. Swimming speed alteration of larvae of Balanus amphitrite as a behavioural end-point for laboratory toxicological bioassays. Mar. Biol. 2006, 149, 87–96. [Google Scholar] [CrossRef]
- Garaventa, F.; Gambardella, C.; Di Fino, A.; Pittore, M.; Faimali, M. Swimming speed alteration of Artemia sp. and Brachionus plicatilis as a sub-lethal behavioural end-point for ecotoxicological surveys. Ecotoxicology 2010, 19, 512–519. [Google Scholar] [CrossRef] [PubMed]
- Manfra, L.; Savorelli, F.; Di Lorenzo, B.; Libralato, G.; Comin, S.; Conti, D.; Floris, B.; Francese, M.; Gallo, M.L.; Gartner, I.; et al. Intercalibration of ecotoxicity testing protocols with Artemia franciscana. Ecol. Indic. 2015, 57, 41–47. [Google Scholar] [CrossRef]
- Andrés-Costa, M.J.; Proctor, K.; Sabatini, M.T.; Gee, A.P.; Lewis, S.E.; Pico, Y.; Kasprzyk-Hordern, B. Enantioselective transformation of fluoxetine in water and its ecotoxicological relevance. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef]
- Calisto, V.; Esteves, V.I. Psychiatric pharmaceuticals in the environment. Chemosphere 2009, 77, 1257–1274. [Google Scholar] [CrossRef] [PubMed]
- Kolpin, D.W.; Furlong, E.; Meyer, M.; Thurman, M.E.; Zaugg, S.; Barber, L.; Buxton, H. Pharmaceuticals, Hormones, and Other Organic Wastewater Contaminants in U.S. Streams, 1999–2000: A National Reconnaissance. Environ. Sci. Technol. 2002, 36, 1202–1211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meador, J.P.; Yeh, A.; Young, G.; Gallagher, E.P. Contaminants of emerging concern in a large temperate estuary. Environ. Pollut. 2016, 213, 254–267. [Google Scholar] [CrossRef] [Green Version]
- Barry, M.J. ‘Effects of fluoxetine on the swimming and behavioural responses of the Arabian killifish’. Ecotoxicology 2013, 22, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Bossus, M.C.; Guler, Y.Z.; Short, S.J.; Morrison, E.R.; Ford, A.T. Behavioural and transcriptional changes in the amphipod Echinogammarus marinus exposed to two antidepressants, fluoxetine and sertraline. Aquat. Toxicol. 2014, 151, 46–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campos, B.; Piña, B.; Fernández-Sanjuán, M.; Lacorte, S.; Barata, C. Enhanced offspring production in Daphnia magna clones exposed to serotonin reuptake inhibitors and 4-nonylphenol. Stage- and food-dependent effects. Aquat. Toxicol. 2012, 109, 100–110. [Google Scholar] [CrossRef] [PubMed]
- Campos, B.; Garcia-Reyero, N.; Rivetti, C.; Escalon, L.; Habib, T.; Tauler, R.; Tsakovski, S.; Pina, B.; Barata, C. Identification of Metabolic Pathways in Daphnia magna Explaining Hormetic Effects of Selective Serotonin Reuptake Inhibitors and 4-Nonylphenol Using Transcriptomic and Phenotypic Responses. Environ. Sci. Technol. 2013, 47, 9434–9443. [Google Scholar] [CrossRef] [PubMed]
- Campos, B.; Rivetti, C.; Kress, T.; Barata, C.; Dircksen, H. Depressing antidepressant: Fluoxetine affects serotonin neurons causing adverse reproductive responses in Daphnia magna. Environ. Sci. Technol. 2016. [Google Scholar] [CrossRef]
- Lazzara, R.; Blázquez, M.; Porte, C.; Barata, C. Low environmental levels of fluoxetine induce spawning and changes in endogenous estradiol levels in the zebra mussel Dreissena polymorpha. Aquat. Toxicol. 2012, 106, 123–130. [Google Scholar] [CrossRef] [Green Version]
- Perreault, H.; Semsar, K.; Godwin, J. Fluoxetine treatment decreases territorial aggression in a coral reef fish. Physiol. Behav. 2003, 79, 719–724. [Google Scholar] [CrossRef]
- Dzieweczynski, T.L.; Hebert, O.L. Fluoxetine alters behavioral consistency of aggression and courtship in male Siamese fighting fish, Betta splendens. Physiol. Behav. 2012, 107, 92–97. [Google Scholar] [CrossRef]
- Huber, R.; Delago, A. Serotonin alters decisions to withdraw in fighting crayfish, Astacus astacus: The motivational concept revisited. J. Comp. Physiol. A Sens. NeuralBehav. Physiol. 1998, 182, 573–583. [Google Scholar] [CrossRef]
- De Castro-Català, N.; Muñoz, I.; Riera, J.L.; Ford, A.T. Evidence of low dose effects of the antidepressant fluoxetine and the fungicide prochloraz on the behavior of the keystone freshwater invertebrate Gammarus pulex. Environ. Pollut. 2017, 231, 406–414. [Google Scholar] [CrossRef] [Green Version]
- Hazelton, P.D.; Cope, W.G.; Mosher, S.; Pandolfo, T.J.; Belden, J.B.; Barnhart, M.C.; Bringolf, R.B. Fluoxetine alters adult freshwater mussel behavior and larval metamorphosis. Sci. Total Environ. 2013, 445, 94–100. [Google Scholar] [CrossRef]
- Weinberger, J.; Klaper, R. Environmental concentrations of the selective serotonin reuptake inhibitor fluoxetine impact specific behaviors involved in reproduction, feeding and predator avoidance in the fish Pimephales promelas (fathead minnow). Aquat. Toxicol. 2014, 151, 77–83. [Google Scholar] [CrossRef] [Green Version]
- Barbosa, A.; Alves, F.L.; Pereira, A.D.S.F.; Ide, L.M.; Hoffmann, A. Behavioral characterization of the alarm reaction and anxiolytic-like effect of acute treatment with fluoxetine in piaucu fish. Physiol. Behav. 2012, 105, 784–790. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.M.; Saaristo, M.; Bertram, M.G.; Lewis, P.J.; Coggan, T.L.; Clarke, B.O.; Wong, B.B.M. The psychoactive pollutant fluoxetine compromises antipredator behaviour in fish. Environ. Pollut. 2017, 222, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Egan, R.J.; Bergner, C.L.; Hart, P.C.; Cachat, J.M.; Canavello, P.R.; Elegante, M.F.; Elkhayat, S.I.; Bartels, B.K.; Tien, A.K.; Tien, D.H.; et al. Understanding behavioral and physiological phenotypes of stress and anxiety in zebrafish. Behav. Brain Res. 2009, 205, 38–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamilton, T.J.; Kwan, G.T.; Gallup, J.; Tresguerres, M. Acute fluoxetine exposure alters crab anxiety-like behaviour, but not aggressiveness. Sci. Rep. 2016, 6, 19850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, R.Y.; Oxendine, S.E.; Godwin, J. Behavioral and neurogenomic transcriptome changes in wild-derived zebrafish with fluoxetine treatment. BMC Genom. 2013, 14, 348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guler, Y.; Ford, A.T. Anti-depressants make amphipods see the light. Aquat. Toxicol. 2010, 99, 397–404. [Google Scholar] [CrossRef]
- Tierney, A.J.; Hanzlik, K.N.; Hathaway, R.M.; Powers, C.; Roy, M. Effects of fluoxetine on growth and behavior in the crayfish Orconectes rusticus. Mar. Freshw. Behav. Physiol. 2016, 49, 133–145. [Google Scholar] [CrossRef]
- Kosjek, T.; Perko, S.; Zupanc, M.; ZanoŠki Hren, M.; Landeka Dragičević, T.; Žigon, D.; Kompare, B.; Heath, E. Environmental occurrence, fate and transformation of benzodiazepines in water treatment. Water Res. 2012, 46, 355–368. [Google Scholar] [CrossRef]
- Brodin, T.; Nordling, J.; Lagesson, A.; Klaminder, J.; Hellström, G.; Christensen, B.; Fick, J. Environmental relevant levels of a benzodiazepine (oxazepam) alters important behavioral traits in a common planktivorous fish, (Rutilus rutilus). J. Toxicol. Environ. Health Part. A Curr. Issues 2017, 80, 963–970. [Google Scholar] [CrossRef]
- Chiffre, A.; Clérandeau, C.; Dwoinikoff, C.; Le Bihanic, F.; Budzinski, H.; Geret, F.; Cachot, J. Psychotropic drugs in mixture alter swimming behaviour of Japanese medaka (Oryzias latipes) larvae above environmental concentrations. Environ. Sci. Pollut. Res. 2016, 23, 4964–4977. [Google Scholar] [CrossRef] [PubMed]
- Kubec, J.; Hossain, M.; Grabicová, K.; Randák, T.; Kouba, A.; Grabic, R.; Roje, S.; Buřič, M. Oxazepam Alters the Behavior of Crayfish at Diluted Concentrations, Venlafaxine Does Not. Water 2019, 11, 196. [Google Scholar] [CrossRef] [Green Version]
- Brodin, T.; Fick, J.; Jonsson, M.; Klaminder, J. Dilute concentrations of a psychiatric drug alter behavior of fish from natural populations. Science 2013, 339, 814–815. [Google Scholar] [CrossRef] [PubMed]
- Klaminder, J.; Hellström, G.; Fahlman, J.; Jonsson, M.; Fick, J.; Lagesson, A.; Bergman, E.; Brodin, T. Drug-Induced Behavioral Changes: Using Laboratory Observations to Predict Field Observations. Front. Environ. Sci. 2016, 4, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Saaristo, M.; Lagesson, A.; Bertram, M.G.; Fick, J.; Klaminder, J.; Johnstone, C.P.; Wong, B.B.M.; Brodin, T. Behavioural effects of psychoactive pharmaceutical exposure on European perch (Perca fluviatilis) in a multi-stressor environment. Sci. Total Environ. 2019, 655, 1311–1320. [Google Scholar] [CrossRef] [PubMed]
- Hellström, G.; Klaminder, J.; Finn, F.; Persson, L.; Alanärä, A.; Jonsson, M.; Fick, J.; Brodin, T. GABAergic anxiolytic drug in water increases migration behaviour in salmon. Nat. Commun. 2016, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasprzyk-Hordern, B.; Dinsdale, R.M.; Guwy, A.J. The removal of pharmaceuticals, personal care products, endocrine disruptors and illicit drugs during wastewater treatment and its impact on the quality of receiving waters. Water Res. 2009, 43, 363–380. [Google Scholar] [CrossRef]
- Lajeunesse, A.; Smyth, S.A.; Barclay, K.; Sauvé, S.; Gagnon, C. Distribution of antidepressant residues in wastewater and biosolids following different treatment processes by municipal wastewater treatment plants in Canada. Water Res. 2012, 46, 5600–5612. [Google Scholar] [CrossRef]
- Minguez, L.; Farcy, E.; Ballandonne, C.; Lepailleur, A.; Serpentini, A.; Lebel, J.M.; Bureau, R.; Halm-Lemeille, M.P. Acute toxicity of 8 antidepressants: What are their modes of action? Chemosphere 2014, 108, 314–319. [Google Scholar] [CrossRef]
- Petrie, B.; Barden, R.; Kasprzyk-Hordern, B. A review on emerging contaminants in wastewaters and the environment: Current knowledge, understudied areas and recommendations for future monitoring. Water Res. 2014, 72, 3–27. [Google Scholar] [CrossRef]
- Subramanian, P.; Subbaraj, R. Effect of imipramine and amitriptyline on circadian locomotor activity rhythm in the field mouse Mus booduga. Proc. Anim. Sci. 1990, 99, 503–507. [Google Scholar] [CrossRef]
- Shakunthala, V.; Najafabadi, Z.G.; Sathisha, B. Effect of Amitriptyline hydrochloride an antidepressant on courtship and reproducibility of D. melanogaster. Int. J. Currrent Microbiol. Appl. Sci. 2014, 3, 649–656. [Google Scholar]
- Egan, J.; Earley, C.J.; Leonard, B.E. The effect of amitriptyline and mianserine (Org. GB94) on food motivated behaviour of rats trained in a runway: Possible correlation with biogenic amine concentration in the limbic system. Psychopharmacology 1979, 61, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Enginar, N.; Hatipoǧlu, I.; Firtina, M. Evaluation of the acute effects of amitriptyline and fluoxetine on anxiety using grooming analysis algorithm in rats. Pharmacol. Biochem. Behav. 2008, 89, 450–455. [Google Scholar] [CrossRef]
- Parra, A.; Vinader-Caerols, C.; Ferrer-Añó, A.; Urquiza, A.; Monleón, S. The effect of amitriptyline on inhibitory avoidance in mice is dose-dependent. Psicothema 2009, 21, 528–530. [Google Scholar]
- Fong, P.P.; DiPenta, K.E.; Jonik, S.M.; Ward, C.D. Short-term exposure to tricyclic antidepressants delays righting time in marine and freshwater snails with evidence for low-dose stimulation of righting speed by imipramine. Environ. Sci. Pollut. Res. 2019. [Google Scholar] [CrossRef]
- Fong, P.P.; Molnar, N. Antidepressants cause foot detachment from substrate in five species of marine snail. Mar. Environ. Res. 2013, 84, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Fong, P.P.; Bury, T.B.; Dworkin-Brodsky, A.D.; Jasion, C.M.; Kell, R.C. The antidepressants venlafaxine (“Effexor”) and fluoxetine (“Prozac”) produce different effects on locomotion in two species of marine snail, the oyster drill (Urosalpinx cinerea) and the starsnail (Lithopoma americanum). Mar. Environ. Res. 2015, 103, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Bisesi, J.H.; Bridges, W.; Klaine, S.J. Reprint of: Effects of the antidepressant venlafaxine on fish brain serotonin and predation behavior. Aquat. Toxicol. 2014, 151, 88–96. [Google Scholar] [CrossRef]
- De Oliveira, R.A.; Cunha, G.M.A.; Karla, K.D.; De Bruin, G.S.; Dos Santos-Filho, E.A.; Viana, G.S.B.; De Bruin, V.M.S. The effect of venlafaxine on behaviour, body weight and striatal monoamine levels on sleep-deprived female rats. Pharmacol. Biochem. Behav. 2004, 79, 499–506. [Google Scholar] [CrossRef]
- McGrath, C.; Norman, T.R. The effect of venlafaxine treatment on the behavioural and neurochemical changes in the olfactory bulbectomised rat. Psychopharmacology 1998, 136, 394–401. [Google Scholar] [CrossRef] [PubMed]
- Nowakowska, E.; Kus, K.; Chodera, A. Comparison of Behavioural Effects of Venlafaxine and Imipramine in Rats. Arzneimittelforschung 2012, 53, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.H.; Gu, J.Y.; Zhang, X.L.; Dong, J.; Yang, J.; Zhang, Y.L.; Ning, Q.F.; Shan, X.W.; Li, Y. Venlafaxine ameliorates the depression-like behaviors and hippocampal S100B expression in a rat depression model. Behav. Brain Funct. 2016, 12, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mesquita, S.R.; Guilhermino, L.; Guimarães, L. Biochemical and locomotor responses of Carcinus maenas exposed to the serotonin reuptake inhibitor fluoxetine. Chemosphere 2011, 85, 967–976. [Google Scholar] [CrossRef] [PubMed]
- Bradley, D.J.; Forward, R.B. Phototaxis of adult brine shrimp, Artemia salina. Can. J. Zool. 1984, 62, 2357–2359. [Google Scholar] [CrossRef]
- Dojmi Di Delupis, G.; Rotondo, V. Phototaxis in aquatic invertebrates: Possible use in ecotoxicity test. Ecotoxicol. Environ. Saf. 1988, 16, 189–193. [Google Scholar] [CrossRef]
- Sumpter, J.P.; Donnachie, R.L.; Johnson, A.C. The apparently very variable potency of the anti-depressant fluoxetine. Aquat. Toxicol. 2014, 151, 57–60. [Google Scholar] [CrossRef] [PubMed]
- Melvin, S.D.; Petit, M.A.; Duvignacq, M.C.; Sumpter, J.P. Towards improved behavioural testing in aquatic toxicology: Acclimation and observation times are important factors when designing behavioural tests with fish. Chemosphere 2017, 180, 430–436. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Davis, R.L.; Roman, G. Exploratory activity in Drosophila requires the kurtz nonvisual arrestin. Genetics 2007, 175, 1197–1212. [Google Scholar] [CrossRef] [Green Version]
- Montgomery, K.C. The relation between exploratory behaviour and spontaneous alternation in the white rat. J. Comp. Physiol. Psychol. 1951, 44, 582–589. [Google Scholar] [CrossRef]
- Oldham, J.; Morlock, H. The effects of open-field size on activity in the Mongolian gerbil. Psychon. Sci. 1970, 20, 290. [Google Scholar] [CrossRef]
- Forward, R.B.; Rittschof, D. Brine shrimp larval photoresponses involved in diel vertical migration: Activation by fish mucus and modified amino sugars. Limnol. Oceanogr. 1999, 44, 1904–1916. [Google Scholar] [CrossRef]
- Pearre, S. Problems of detection and interpretation of vertical migration. J. Plankton Res. 1979, 1, 29–44. [Google Scholar] [CrossRef]
- Basnet, R.; Zizioli, D.; Taweedet, S.; Finazzi, D.; Memo, M. Zebrafish Larvae as a Behavioral Model in Neuropharmacology. Biomedicines 2019, 7, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Lin, J.; Zhang, Y.; Liu, X.; Chen, X.Q.; Xu, M.Q.; He, L.; Li, S.; Guo, N. Differential behavioral responses of zebrafish larvae to yohimbine treatment. Psychopharmacology 2015, 232, 197–208. [Google Scholar] [CrossRef]
- Van Den Bos, R.; Mes, W.; Galligani, P.; Heil, A.; Zethof, J.; Flik, G.; Gorissen, M. Further characterisation of differences between TL and AB Zebrafish (Danio rerio): Gene expression, physiology and behaviour at day 5 of the larval stage. PLoS ONE 2017, 12, e0175420. [Google Scholar] [CrossRef]
- Nielsen, M.E.; Roslev, P. Behavioral responses and starvation survival of Daphnia magna exposed to fluoxetine and propranolol. Chemosphere 2018, 211, 978–985. [Google Scholar] [CrossRef]
- Mulloney, B.; Acevedo, L.D.; Bradbury, A.G. Modulation of the crayfish swimmeret rhythm by octopamine and the neuropeptide proctolin. J. Neurophysiol. 1987, 58, 584–597. [Google Scholar] [CrossRef]
- Bauknecht, P.; Jékely, G. Ancient coexistence of norepinephrine, tyramine, and octopamine signaling in bilaterians. BMC Biol. 2017, 15, 6. [Google Scholar] [CrossRef] [Green Version]
- Rivetti, C.; Campos, B.; Barata, C. Low environmental levels of neuro-active pharmaceuticals alter phototactic behaviour and reproduction in Daphnia magna. Aquat. Toxicol. 2016, 170, 289–296. [Google Scholar] [CrossRef]
- Rivetti, C.; Campos, B.; Piña, B.; Raldúa, D.; Kato, Y.; Watanabe, H.; Barata, C. Tryptophan hydroxylase (TRH) loss of function mutations induce growth and behavioral defects in Daphnia magna. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourin, M.; Hascoët, M. The mouse light-dark box test. Eur. J. Pharmacol. 2003, 463, 55–65. [Google Scholar] [CrossRef]
- Magurran, A.E.; Girling, S.L. Predator model recognition and response habituation in shoaling minnows. Anim. Behav. 1986, 34, 510–518. [Google Scholar] [CrossRef]
- Wong, K.; Elegante, M.; Bartels, B.; Elkhayat, S.; Tien, D.; Roy, S.; Goodspeed, J.; Suciu, C.; Tan, J.; Grimes, C.; et al. Analyzing habituation responses to novelty in zebrafish (Danio rerio). Behav. Brain Res. 2010, 208, 450–457. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Class | Environment | Concentration | Source | Behaviour Impacts | Source |
|---|---|---|---|---|---|---|
| fluoxetine hydrochloride | Selective Serotonin Reuptake Inhibitor (SSRI) | effluents | 0.001–5 µg/L | [17,18,19,20] | activity | [21,22] |
| surface waters | 0.012–0.02 µg/L | reproduction | [23,24,25,26,27] | |||
| marine env | 0.012 µg/L | aggression | [27,28,29] | |||
| feeding | [30,31,32] | |||||
| predator avoidance | [21,32,33,34] | |||||
| stress/anxiety | [35,36,37] | |||||
| taxis | [38,39] | |||||
| oxazepam | Benzodiazepine (BZD) | effluents | 0.25–0.73 ug/L | [18,20,40] | activity | [41,42,43] |
| surface waters | 0.02–0.58 ug/L | feeding | [44] | |||
| boldness | [41,45,46] | |||||
| social behaviour | [44] | |||||
| migration | [47] | |||||
| amitriptyline hydrochloride | Tricyclic Antidepressant (TCA) | effluents | <2–357 ng/L | [48,49,50,51] | activity | [52] |
| surface waters | <0.5–72 ng/L | reproduction | [53] | |||
| bio-solids | 263–632 ng/g | feeding | [54] | |||
| stress/anxiety | [55] | |||||
| memory & learning | [56] | |||||
| orientation | [57] | |||||
| venlafaxine hydrochloride | Selective Serotonin and Norepinephrine Reuptake Inhibitor (SNRI) | effluents | 600–1454 ng/L | [49,50] | activity | [58,59] |
| surface waters | 187 ng/L | feeding | [60] | |||
| bio-solids | 289–499 ng/g | stress/anxiety | [61,62,63,64] |
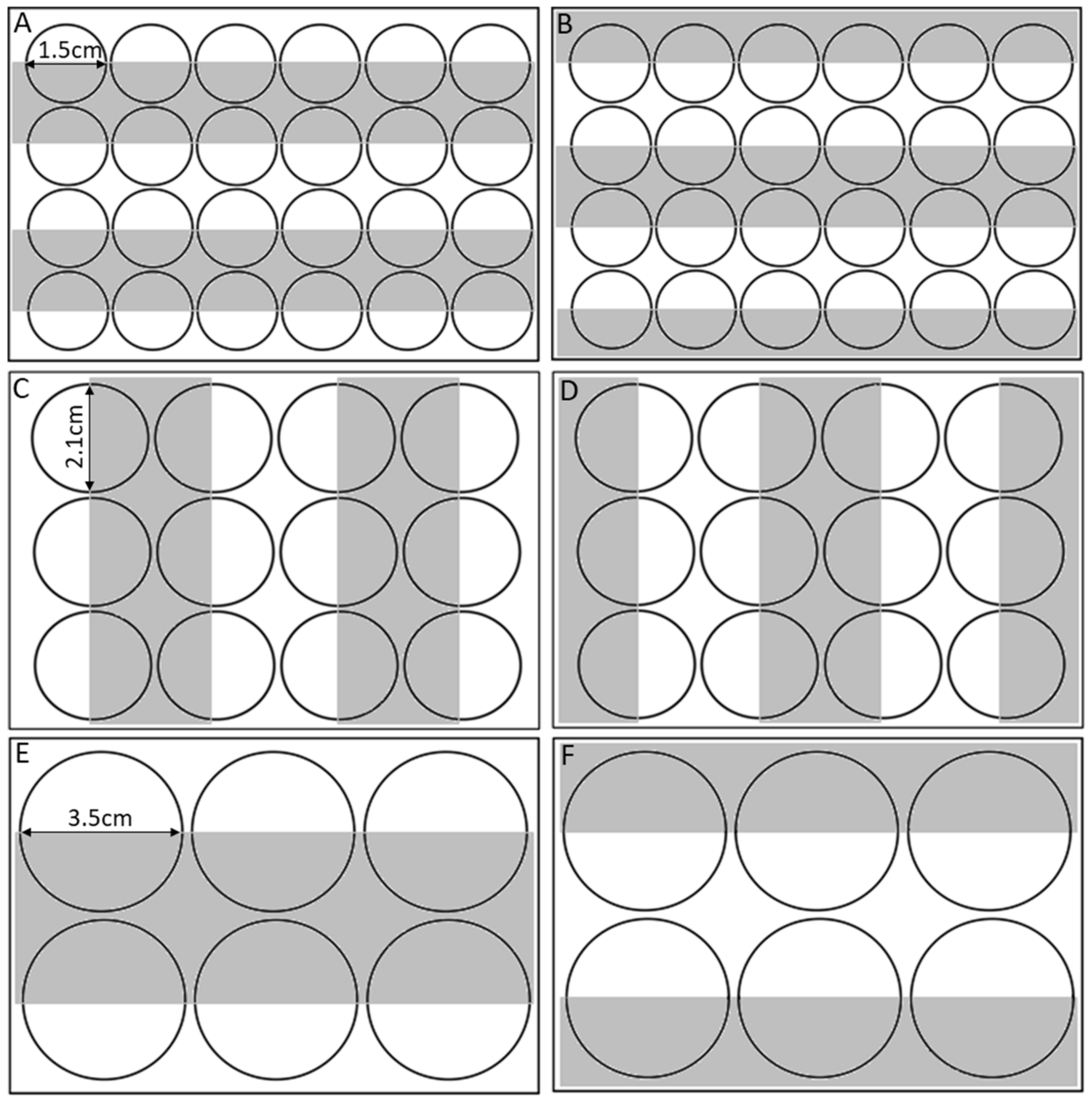
| Arena | Diameter | Area | Volume AFSW | Replicates |
|---|---|---|---|---|
| Small | 1.5 cm | 3 cm3 | 1.5 mL | 24 |
| Medium | 2.1 cm | 6 cm3 | 3 mL | 24 |
| Large | 3.5 cm | 16.4 cm3 | 8 mL | 30 |
| Arena Size | Acrylic Plate Used | Light Intensity | Replicates |
|---|---|---|---|
| Small | 1 | 100% | 24 |
| 5% | 24 | ||
| 2 | 100% | 24 | |
| 5% | 24 | ||
| Medium | 1 | 100% | 24 |
| 5% | 24 | ||
| 2 | 100% | 24 | |
| 5% | 24 | ||
| Large | 1 | 100% | 30 |
| 5% | 30 | ||
| 2 | 100% | 30 | |
| 5% | 30 |
| Comparison | 2-min | 10-s | ||||||
|---|---|---|---|---|---|---|---|---|
| N-df | D-df | F | p | N-df | D-df | F | p | |
| arena size | 2 | 75 | 16.3 | <0.001 | 2 | 74 | 16.2 | <0.001 |
| time | 3 | 225 | 13.6 | <0.001 | 47 | 3522 | 7.0 | <0.001 |
| arena size * time | 6 | 225 | 1.5 | 0.180 | 94 | 3522 | 1.7 | <0.001 |
| ICC | 0.679 | 0.475 | ||||||
| R2 | 0.271 | 0.217 | ||||||
| Comparison | Num df | Den df | F | p |
|---|---|---|---|---|
| arena size | 2 | 344.64 | 13.37 | <0.001 |
| light intensity | 1 | 302.92 | 1.02 | 0.313 |
| light phase | 1 | 337.59 | 4.35 | 0.038 |
| arena size * light intensity | 2 | 303.77 | 3.72 | 0.025 |
| arena size * light phase | 2 | 305.09 | 54.75 | <0.001 |
| light intensity * light phase | 1 | 303.25 | 8.07 | 0.005 |
| arena size * light intensity * light phase | 2 | 307.19 | 0.73 | 0.483 |
| ICC = 0.364 | ||||
| R2 = 0.198 |
| Comparison | F | Num df | Den df | p |
|---|---|---|---|---|
| treatment | 5.328 | 3 | 111 | 0.002 |
| light phase | 8.221 | 3 | 392 | <0.001 |
| exposure period | 17.400 | 2 | 111 | <0.001 |
| treatment * light phase | 0.757 | 9 | 392 | 0.657 |
| treatment * exposure period | 0.663 | 6 | 111 | 0.679 |
| light phase * exposure period | 1.728 | 6 | 392 | 0.113 |
| treatment * light phase * exposure period | 0.520 | 18 | 392 | 0.949 |
| ICC = 0.315 | ||||
| R2 = 0.212 |
| Comparison | F | Num df | Den df | p |
|---|---|---|---|---|
| treatment | 1.602 | 3 | 114 | 0.193 |
| light phase | 6.490 | 3 | 367 | <0.001 |
| exposure period | 9.926 | 2 | 114 | <0.001 |
| treatment * light phase | 0.563 | 9 | 367 | 0.827 |
| treatment * exposure period | 0.534 | 6 | 114 | 0.782 |
| light phase * exposure period | 2.783 | 6 | 367 | 0.012 |
| treatment * light phase * exposure period | 1.109 | 18 | 367 | 0.341 |
| ICC = 0.557 | ||||
| R2 = 0.174 |
| Comparison | F | Num df | Den df | p |
|---|---|---|---|---|
| treatment | 2.204 | 3 | 112 | 0.092 |
| light phase | 8.415 | 3 | 386 | <0.001 |
| exposure period | 4.258 | 2 | 112 | 0.017 |
| treatment * light phase | 0.537 | 9 | 386 | 0.847 |
| treatment * exposure period | 0.703 | 6 | 112 | 0.648 |
| light phase * exposure period | 0.861 | 6 | 386 | 0.524 |
| treatment * light phase * exposure period | 0.393 | 18 | 386 | 0.989 |
| ICC = 0.318 | ||||
| R2 = 0.116 |
| Comparison | F | Num df | Den df | p |
|---|---|---|---|---|
| treatment | 1.180 | 3 | 116 | 0.321 |
| light phase | 18.062 | 3 | 394 | <0.001 |
| exposure period | 3.091 | 2 | 116 | 0.049 |
| treatment * light phase | 0.873 | 9 | 394 | 0.550 |
| treatment * exposure period | 0.200 | 6 | 116 | 0.976 |
| light phase * exposure period | 2.095 | 6 | 394 | 0.053 |
| treatment * light phase * exposure period | 0.581 | 18 | 394 | 0.913 |
| ICC = 0.410 | ||||
| R2 = 0.121 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kohler, S.A.; Parker, M.O.; Ford, A.T. High-Throughput Screening of Psychotropic Compounds: Impacts on Swimming Behaviours in Artemia franciscana. Toxics 2021, 9, 64. https://doi.org/10.3390/toxics9030064
Kohler SA, Parker MO, Ford AT. High-Throughput Screening of Psychotropic Compounds: Impacts on Swimming Behaviours in Artemia franciscana. Toxics. 2021; 9(3):64. https://doi.org/10.3390/toxics9030064
Chicago/Turabian StyleKohler, Shanelle A., Matthew O. Parker, and Alex T. Ford. 2021. "High-Throughput Screening of Psychotropic Compounds: Impacts on Swimming Behaviours in Artemia franciscana" Toxics 9, no. 3: 64. https://doi.org/10.3390/toxics9030064
APA StyleKohler, S. A., Parker, M. O., & Ford, A. T. (2021). High-Throughput Screening of Psychotropic Compounds: Impacts on Swimming Behaviours in Artemia franciscana. Toxics, 9(3), 64. https://doi.org/10.3390/toxics9030064