
Salinity Alters Toxicity of Commonly Used Pesticides in a Model Euryhaline Fish Species (Menidia beryllina)



Abstract
:
1. Introduction
2. Materials and Methods
2.1. Exposure Chemicals
2.2. Organism Husbandry
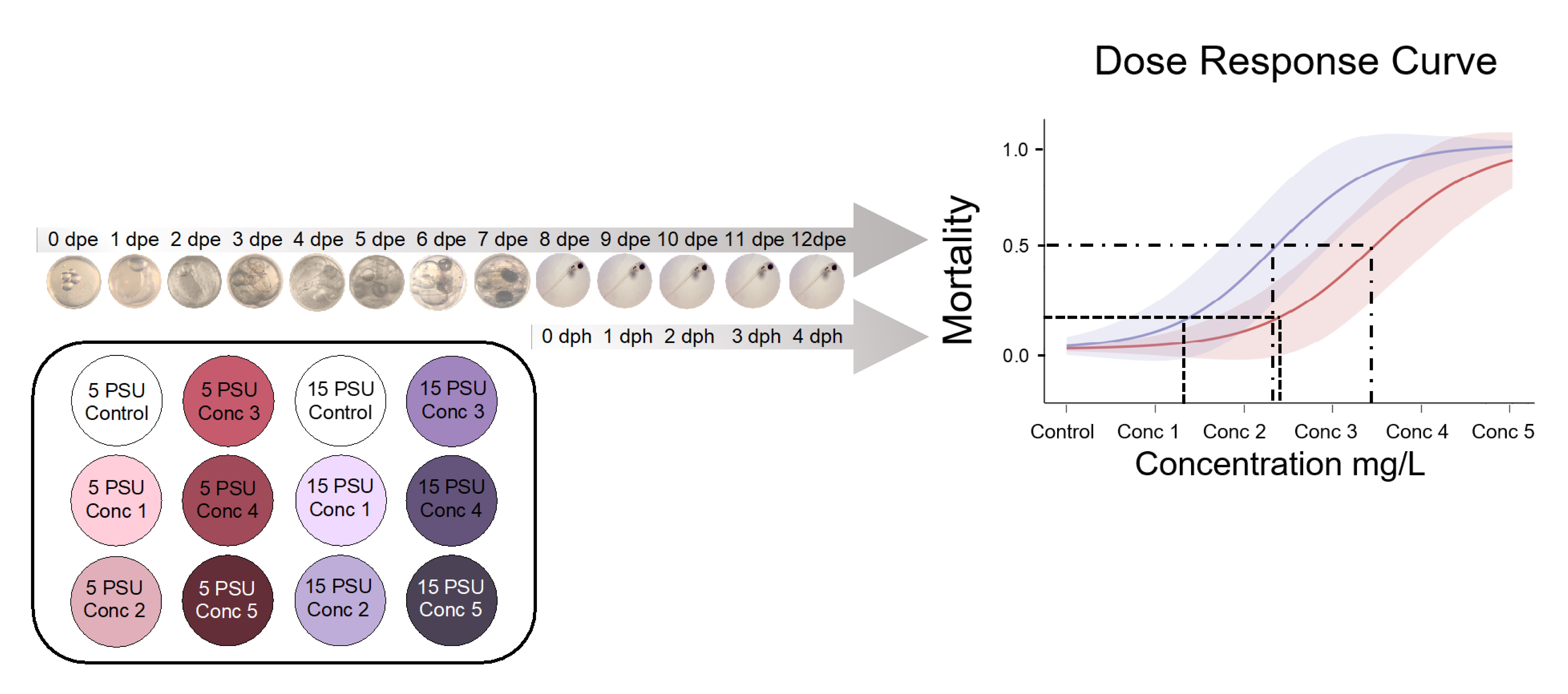
2.3. Bioassays
2.4. Analytical Chemistry
2.4.1. Extraction and Sample Preparation
2.4.2. High Performance Liquid Chromatographic (HPLC) Analysis
2.4.3. Gas Chromatographic Analysis
2.5. Statistical Analysis
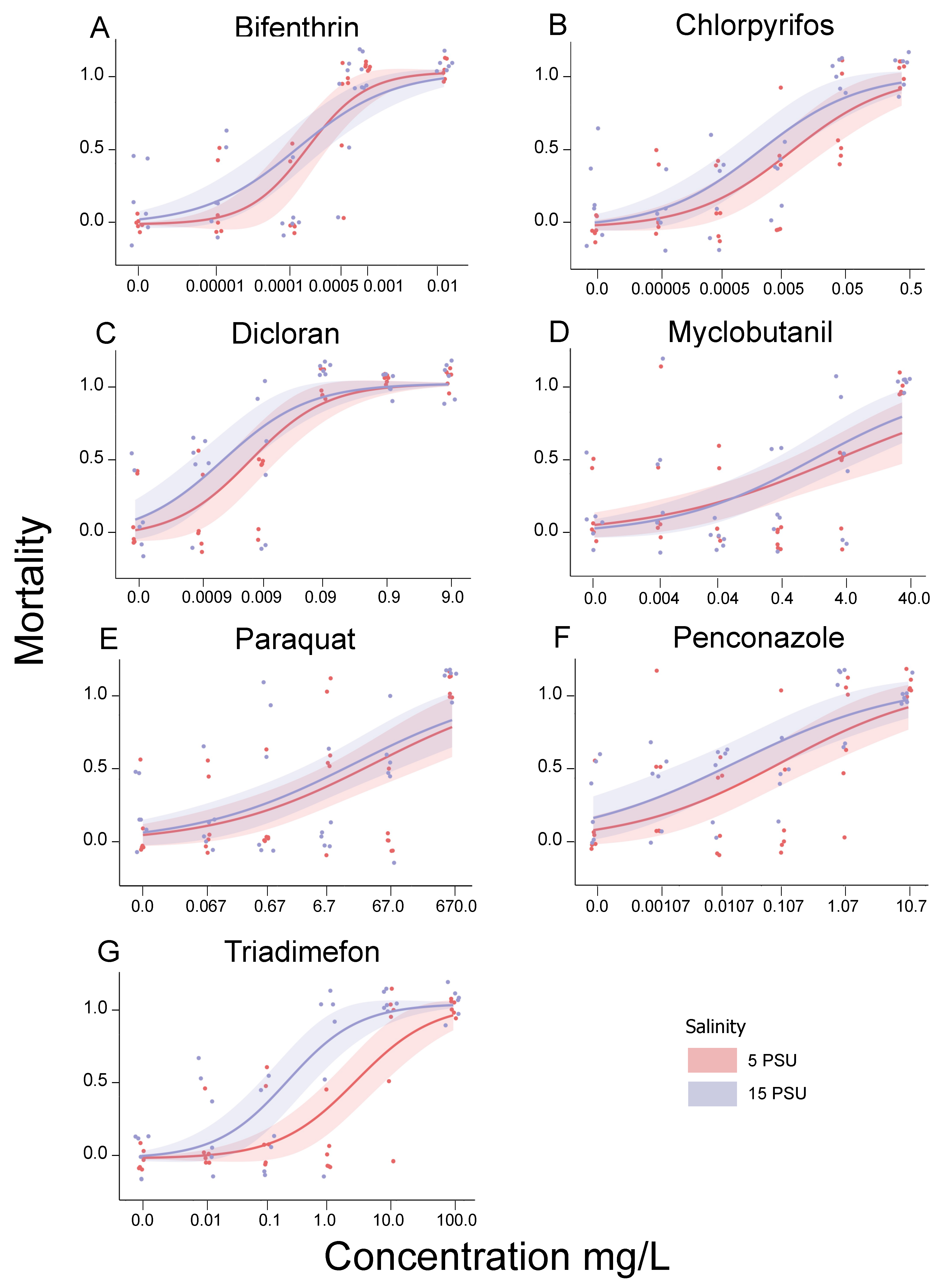
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Beck, M.W.; Heck, K.L.; Able, K.W.; Childers, D.L.; Eggleston, D.B.; Gillanders, B.M.; Halpern, B.S.; Hays, C.G.; Hoshino, K.; Minello, T.J. The role of nearshore ecosystems as fish and shellfish nurseries. Issues Ecol. 2003, 11, 1–12. [Google Scholar]
- Delorenzo, M.E. Impacts of climate change on the ecotoxicology of chemical contaminants in estuarine organisms. Curr. Zool. 2015, 61, 641–652. [Google Scholar] [CrossRef] [Green Version]
- Pawar, A.P.; Sanaye, S.V.; Shyama, S.; Sreepada, R.A.; Dake, A.S. Effects of salinity and temperature on the acute toxicity of the pesticides, dimethoate and chlorpyrifos in post-larvae and juveniles of the whiteleg shrimp. Aquac. Rep. 2020, 16. [Google Scholar] [CrossRef]
- Field, C.B. Climate Change 2014–Impacts, Adaptation and Vulnerability: Regional Aspects; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Hong, B.; Shen, J. Responses of estuarine salinity and transport processes to potential future sea-level rise in the Chesapeake Bay. Estuar. Coast. Shelf Sci. 2012, 104–105, 33–45. [Google Scholar] [CrossRef]
- Kaushal, S.S.; Pace, M.L.; Groffman, P.M.; Band, L.E.; Belt, K.T.; Meyer, P.M.; Welty, C. Land use and climate variability amplify contaminant pulses. EosTrans. Am. Geophys. Union 2010, 91, 221–222. [Google Scholar] [CrossRef] [Green Version]
- Kimbro, D.L.; White, J.W.; Tillotson, H.; Cox, N.; Christopher, M.; Stokes-Cawley, O.; Yuan, S.; Pusack, T.J.; Stallings, C.D. Local and regional stressors interact to drive a salinization-induced outbreak of predators on oyster reefs. Ecosphere 2017, 8, e01992. [Google Scholar] [CrossRef]
- Cook, B.I.; Mankin, J.S.; Anchukaitis, K.J. Climate change and drought: From past to future. Curr. Clim. Chang. Rep. 2018, 4, 164–179. [Google Scholar] [CrossRef]
- Hobbs, J.A.; Lewis, L.S.; Willmes, M.; Denney, C.; Bush, E. Complex life histories discovered in a critically endangered fish. Sci. Rep. 2019, 9, 16772. [Google Scholar] [CrossRef]
- Segarra, A.; Mauduit, F.; Amer, N.R.; Biefel, F.; Hladik, M.L.; Connon, R.E.; Brander, S.M. Salinity Changes the Dynamics of Pyrethroid Toxicity in Terms of Behavioral Effects on Newly Hatched Delta Smelt Larvae. Toxics 2021, 9, 40. [Google Scholar] [CrossRef]
- DeCourten, B.M.; Connon, R.E.; Brander, S.M. Direct and indirect parental exposure to endocrine disruptors and elevated temperature influences gene expression across generations in a euryhaline model fish. PeerJ 2019, 7, e6156. [Google Scholar] [CrossRef]
- DeCourten, B.; Romney, A.; Brander, S. The Heat Is On: Complexities of Aquatic Endocrine Disruption in a Changing Global Climate. In Evaluating Water Quality to Prevent Future Disasters; Separation Science and Technology; Academic Press: Cambridge, MA, USA, 2019; pp. 13–49. [Google Scholar]
- Hladik, M.L.; Kolpin, D.W.; Kuivila, K.M. Widespread occurrence of neonicotinoid insecticides in streams in a high corn and soybean producing region, USA. Environ. Pollut. 2014, 193, 189–196. [Google Scholar] [CrossRef] [PubMed]
- McKnight, U.S.; Rasmussen, J.J.; Kronvang, B.; Binning, P.J.; Bjerg, P.L. Sources, occurrence and predicted aquatic impact of legacy and contemporary pesticides in streams. Environ. Pollut. 2015, 200, 64–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuivila, K.; Hladik, M. Understanding the occurrence and transport of current-use pesticides in the San Francisco estuary watershed. San Fr. Estuary Watershed Sci. 2008, 6, 6. [Google Scholar] [CrossRef] [Green Version]
- Hapke, W.B.; Morace, J.L.; Nilsen, E.B.; Alvarez, D.A.; Masterson, K. Year-round monitoring of contaminants in Neal and Rogers Creeks, Hood River Basin, Oregon, 2011–2012, and assessment of risks to salmonids. PLoS ONE 2016, 11, e0158175. [Google Scholar] [CrossRef]
- Thompson, T.J.; Briggs, M.A.; Phillips, P.J.; Blazer, V.S.; Smalling, K.L.; Kolpin, D.W.; Wagner, T. Groundwater discharges as a source of phytoestrogens and other agriculturally derived contaminants to streams. Sci. Total Environ. 2021, 755, 142873. [Google Scholar] [CrossRef] [PubMed]
- Saranjampour, P.; Vebrosky, E.N.; Armbrust, K.L. Salinity impacts on water solubility and n-octanol/water partition coefficients of selected pesticides and oil constituents. Environ. Toxicol. Chem. 2017, 36, 2274–2280. [Google Scholar] [CrossRef]
- Activity Coefficient and Solubility in Water. In Environmental Organic Chemistry; Wiley Online Library: Hoboken, NJ, USA, 2002; pp. 133–180.
- Jeon, J.; Kannan, K.; Lim, H.K.; Moon, H.B.; Ra, J.S.; Kim, S.D. Bioaccumulation of perfluorochemicals in pacific oyster under different salinity gradients. Environ. Sci. Technol. 2010, 44, 2695–2701. [Google Scholar] [CrossRef]
- Moreira, L.B.; Diamante, G.; Giroux, M.; Coffin, S.; Xu, E.G.; Moledo de Souza Abessa, D.; Schlenk, D. Impacts of Salinity and Temperature on the Thyroidogenic Effects of the Biocide Diuron in Menidia beryllina. Environ. Sci. Technol. 2018, 52, 3146–3155. [Google Scholar] [CrossRef]
- Fortin, M.G.; Couillard, C.M.; Pellerin, J.; Lebeuf, M. Effects of salinity on sublethal toxicity of atrazine to mummichog (Fundulus heteroclitus) larvae. Mar. Environ. Res. 2008, 65, 158–170. [Google Scholar] [CrossRef]
- Blewett, T.; MacLatchy, D.L.; Wood, C.M. The effects of temperature and salinity on 17-alpha-ethynylestradiol uptake and its relationship to oxygen consumption in the model euryhaline teleost (Fundulus heteroclitus). Aquat. Toxicol. 2013, 127, 61–71. [Google Scholar] [CrossRef]
- Hasenbein, S.; Poynton, H.; Connon, R.E. Contaminant exposure effects in a changing climate: How multiple stressors can multiply exposure effects in the amphipod Hyalella azteca. Ecotoxicology 2018, 27, 845–859. [Google Scholar] [CrossRef] [PubMed]
- Brander, S.M.; Jeffries, K.M.; Cole, B.J.; DeCourten, B.M.; White, J.W.; Hasenbein, S.; Fangue, N.A.; Connon, R.E. Transcriptomic changes underlie altered egg protein production and reduced fecundity in an estuarine model fish exposed to bifenthrin. Aquat. Toxicol. 2016, 174, 247–260. [Google Scholar] [CrossRef] [PubMed]
- Topal, A.; Şişecioğlu, M.; Atamanalp, M.; Işık, A.; Yılmaz, B. The in vitro and in vivo effects of chlorpyrifos on acetylcholinesterase activity of rainbow trout brain. J. Appl. Anim. Res. 2016, 44, 243–247. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; McConnell, L.; Torrents, A. Hydrolysis of chlorpyrifos in natural waters of the Chesapeake Bay. Chemosphere 2001, 44, 1315–1323. [Google Scholar] [CrossRef]
- Berni, I.; Menouni, A.; El, I.G.; Duca, R.-C.; Kestemont, M.-P.; Godderis, L.; El, S.J. Understanding farmers’ safety behavior regarding pesticide use in Morocco. Sustain. Prod. Consum. 2021, 25, 471–483. [Google Scholar] [CrossRef]
- Marrs, T.C.; Ballantyne, B. Pesticide Toxicology and International Regulation; Wiley Online Library: Hoboken, NJ, USA, 2004; Volume 1. [Google Scholar]
- Agency, U. Reregistration Eligibility Decision for DCNA (Dicloran); 738/F-05/003; EPA: Washington, DC, USA, 2006. [Google Scholar]
- Zarn, J.A.; Bruschweiler, B.J.; Schlatter, J.R. Azole fungicides affect mammalian steroidogenesis by inhibiting sterol 14 alpha-demethylase and aromatase. Environ. Health Perspect 2003, 111, 255–261. [Google Scholar] [CrossRef]
- Lin, C.H.; Chou, P.H.; Chen, P.J. Two azole fungicides (carcinogenic triadimefon and non-carcinogenic myclobutanil) exhibit different hepatic cytochrome P450 activities in medaka fish. J. Hazard. Mater. 2014, 277, 150–158. [Google Scholar] [CrossRef]
- Ismail, B.; Sameni, M.; Halimah, M. Evaluation of herbicide pollution in the kerian ricefields of Perak, Malaysia. World Appl. Sci. J. 2011, 15, 5–13. [Google Scholar]
- Rashidipour, M.; Maleki, A.; Kordi, S.; Birjandi, M.; Pajouhi, N.; Mohammadi, E.; Heydari, R.; Rezaee, R.; Rasoulian, B.; Davari, B. Pectin/chitosan/tripolyphosphate nanoparticles: Efficient carriers for reducing soil sorption, cytotoxicity, and mutagenicity of paraquat and enhancing its herbicide activity. J. Agric. Food Chem. 2019, 67, 5736–5745. [Google Scholar] [CrossRef]
- Pateiro-Moure, M.; Nóvoa-Muñoz, J.; Arias-Estévez, M.; López-Periago, E.; Martínez-Carballo, E.; Simal-Gándara, J. Quaternary herbicides retention by the amendment of acid soils with a bentonite-based waste from wineries. J. Hazard. Mater. 2009, 164, 769–775. [Google Scholar] [CrossRef]
- Liu, H.-C.; Chu, T.-Y.; Chen, L.-L.; Gui, W.-J.; Zhu, G.-N. The cardiovascular toxicity of triadimefon in early life stage of zebrafish and potential implications to human health. Environ. Pollut. 2017, 231, 1093–1103. [Google Scholar] [CrossRef] [PubMed]
- Mercadante, R.; Polledri, E.; Rubino, F.M.; Mandic-Rajcevic, S.; Vaiani, A.; Colosio, C.; Moretto, A.; Fustinoni, S. Assessment of penconazole exposure in winegrowers using urinary biomarkers. Environ. Res. 2019, 168, 54–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brander, S.M.; Connon, R.E.; He, G.; Hobbs, J.A.; Smalling, K.L.; Teh, S.J.; White, J.W.; Werner, I.; Denison, M.S.; Cherr, G.N. From ‘omics to otoliths: Responses of an estuarine fish to endocrine disrupting compounds across biological scales. PLoS ONE 2013, 8, e74251. [Google Scholar] [CrossRef]
- Middaugh, D.P.; Hemmer, M.J. Reproductive ecology of the inland silverside, Menidia beryllina,(Pisces: Atherinidae) from Blackwater Bay, Florida. Copeia 1992, 1992, 1446535. [Google Scholar] [CrossRef]
- Yang, L.; Cheng, Q.; Lin, L.; Wang, X.; Chen, B.; Luan, T.; Tam, N.F. Partitions and vertical profiles of 9 endocrine disrupting chemicals in an estuarine environment: Effect of tide, particle size and salinity. Environ. Pollut. 2016, 211, 58–66. [Google Scholar] [CrossRef]
- Tomlin, C. The pesticides manual: A world compendium. Br. Crop. Prot. Counc. 2006, 14, 351. [Google Scholar]
- Ritz, C.; Baty, F.; Streibig, J.C.; Gerhard, D. Dose-response analysis using R. PLoS ONE 2015, 10, e0146021. [Google Scholar] [CrossRef] [Green Version]
- Nagelkerke, N.J. A note on a general definition of the coefficient of determination. Biometrika 1991, 78, 691–692. [Google Scholar] [CrossRef]
- Metropolis, N.; Ulam, S. The monte carlo method. J. Am. Stat. Assoc. 1949, 44, 335–341. [Google Scholar] [CrossRef]
- Hui, T.J.; Ariffin, M.M.; Tahir, N.M. Hydrolysis of chlorpyrifos in aqueous solutions at different temperatures and pH. Malays. J. Anal. Sci. 2010, 14, 50–54. [Google Scholar]
- Shukla, P.; Gopalani, M.; Ramteke, D.S.; Wate, S.R. Influence of salinity on PAH Uptake from water soluble fraction of crude oil in Tilapia mossambica. Bull. Environ. Contam. Toxicol. 2007, 79, 601–605. [Google Scholar] [CrossRef] [PubMed]
- Derby, A.P.; Fuller, N.W.; Hartz, K.E.H.; Segarra, A.; Connon, R.E.; Brander, S.M.; Lydy, M.J. Trophic transfer, bioaccumulation and transcriptomic effects of permethrin in inland silversides, Menidia beryllina, under future climate scenarios. Environ. Pollut. 2021, 275, 116545. [Google Scholar] [CrossRef] [PubMed]
- Lavado, R.; Rimoldi, J.M.; Schlenk, D. Mechanisms of fenthion activation in rainbow trout (Oncorhynchus mykiss) acclimated to hypersaline environments. Toxicol. Appl. Pharmacol. 2009, 235, 143–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riar, N.; Crago, J.; Jiang, W.; Maryoung, L.A.; Gan, J.; Schlenk, D. Effects of salinity acclimation on the endocrine disruption and acute toxicity of bifenthrin in freshwater and euryhaline strains of Oncorhynchus mykiss. Environ. Toxicol. Chem. 2013, 32, 2779–2785. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Vebrosky, E.N.; Richards, M.L.; Armbrust, K.L. Evaluation of dicloran phototoxicity using primary cardiomyocyte culture from Crassostrea virginica. Sci. Total Environ. 2018, 628–629, 1–10. [Google Scholar] [CrossRef]
- Badroo, I.A.; Nandurkar, H.P.; Khanday, A.H. Toxicological impacts of herbicide paraquat dichloride on histological profile (gills, liver, and kidney) of freshwater fish Channa punctatus (Bloch). Environ. Sci. Pollut. Res. 2020, 27, 39054–39067. [Google Scholar] [CrossRef]
- Pang, S.; Guo, M.; Zhang, X.; Yu, L.; Zhang, Z.; Huang, L.; Gao, J.; Li, X. Myclobutanil developmental toxicity, bioconcentration and sex specific response in cholesterol in zebrafish (Denio rerio). Chemosphere 2020, 242, 125209. [Google Scholar] [CrossRef]
- Ma, J.; Li, Y.; Li, W.; Li, X. Hepatotoxicity of paraquat on common carp (Cyprinus carpio L.). Sci. Total Environ. 2018, 616, 889–898. [Google Scholar] [CrossRef]
- Shafer, T.J.; Meyer, D.A.; Crofton, K.M. Developmental neurotoxicity of pyrethroid insecticides: Critical review and future research needs. Environ. Health Perspect. 2005, 113, 123–136. [Google Scholar] [CrossRef]
- Kavitha, P.; Rao, J.V. Toxic effects of chlorpyrifos on antioxidant enzymes and target enzyme acetylcholinesterase interaction in mosquito fish, Gambusia affinis. Environ. Toxicol. Pharmacol. 2008, 26, 192–198. [Google Scholar] [CrossRef]
- Husak, V.V.; Mosiichuk, N.M.; Storey, J.M.; Storey, K.B.; Lushchak, V.I. Acute exposure to the penconazole-containing fungicide Topas partially augments antioxidant potential in goldfish tissues. Comp. Biochem. Physiol. C Toxicol. Pharm. 2017, 193, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Henry, M.J.; Sisler, H.D. Effects of sterol biosynthesis-inhibiting (SBI) fungicides on cytochrome P-450 oxygenations in fungi. Pestic. Biochem. Physiol. 1984, 22, 262–275. [Google Scholar] [CrossRef]
- Fuentes, J.; Eddy, F. Drinking in marine, euryhaline and freshwater teleost fish. In Ionic Regulation in Animals: A Tribute to Professor WTW Potts; Springer: Berlin/Heidelberg, Germany, 1997; pp. 135–149. [Google Scholar]
- Hladik, M.L.; Kuivila, K.M. Assessing the occurrence and distribution of pyrethroids in water and suspended sediments. J. Agric. Food Chem. 2009, 57, 9079–9085. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Chen, B.; Qiu, X.; Chen, M.; Ma, Z.; Yu, X. Distribution and risk assessment of 82 pesticides in Jiulong River and estuary in South China. Chemosphere 2016, 144, 1177–1192. [Google Scholar] [CrossRef]
- Weston, D.P.; Chen, D.; Lydy, M.J. Stormwater-related transport of the insecticides bifenthrin, fipronil, imidacloprid, and chlorpyrifos into a tidal wetland, San Francisco Bay, California. Sci. Total Environ. 2015, 527, 18–25. [Google Scholar] [CrossRef]
- Rico, A.; Dafouz, R.; Vighi, M.; Rodríguez-Gil, J.L.; Daam, M.A. Use of postregistration monitoring data to evaluate the ecotoxicological risks of pesticides to surface waters: A case study with chlorpyrifos in the Iberian Peninsula. Environ. Toxicol. Chem. 2021, 40, 500–512. [Google Scholar] [CrossRef]
- Tan, H.; Zhang, H.; Wu, C.; Wang, C.; Li, Q. Pesticides in surface waters of tropical river basins draining areas with rice–vegetable rotations in Hainan, China: Occurrence, relation to environmental factors, and risk assessment. Environ. Pollut. 2021, 283, 117100. [Google Scholar] [CrossRef] [PubMed]
- Smalling, K.L.; Kuivila, K.M.; Orlando, J.L.; Phillips, B.M.; Anderson, B.S.; Siegler, K.; Hunt, J.W.; Hamilton, M. Environmental fate of fungicides and other current-use pesticides in a central California estuary. Mar. Pollut. Bull. 2013, 73, 144–153. [Google Scholar] [CrossRef]
- Liu, Z.; Qi, P.; Wang, J.; Wang, Z.; Di, S.; Xu, H.; Zhao, H.; Wang, Q.; Wang, X.; Wang, X. Development, validation, comparison, and implementation of a highly efficient and effective method using magnetic solid-phase extraction with hydrophilic-lipophilic-balanced materials for LC-MS/MS analysis of pesticides in seawater. Sci. Total Environ. 2020, 708, 135221. [Google Scholar] [CrossRef]
- Tsai, W.-T. Status of herbicide use, regulatory management and case study of paraquat in Taiwan. Environ. Dev. Sustain. 2020, 22, 2673–2683. [Google Scholar] [CrossRef]
- Rodrigues, E.T.; Alpendurada, M.F.; Guimaraes, A.; Avo, R.; Ferreira, B.; Pardal, M.A. The environmental condition of an estuarine ecosystem disturbed by pesticides. Environ. Sci. Pollut. Res. Int. 2019, 26, 24075–24087. [Google Scholar] [CrossRef] [PubMed]
- Glinski, D.A.; Purucker, S.T.; Van Meter, R.J.; Black, M.C.; Henderson, W.M. Analysis of pesticides in surface water, stemflow, and throughfall in an agricultural area in South Georgia, USA. Chemosphere 2018, 209, 496–507. [Google Scholar] [CrossRef]
- Vebrosky, E.N.; Saranjampour, P.; Crosby, D.G.; Armbrust, K.L. Photodegradation of Dicloran in Freshwater and Seawater. J. Agric. Food Chem. 2018, 66, 2654–2659. [Google Scholar] [CrossRef] [PubMed]
- Noicharoen, D.; Parkpian, P.; Shipin, O.V.; Polprasert, C.; Delaune, R.D.; Kongchum, M. Effect of salinity on adsorption and desorption of paraquat in Pak Phanang river sediment, Thailand. J. Environ. Sci. Health Part A 2012, 47, 1897–1908. [Google Scholar] [CrossRef] [PubMed]
- Tsai, W.T.; Lai, C.W.; Hsien, K.J. The effects of pH and salinity on kinetics of paraquat sorption onto activated clay. Colloids Surf. A Physicochem. Eng. Asp. 2003, 224, 99–105. [Google Scholar] [CrossRef]
- Fytizas, R. Toxicity of paraquat to three marine organisms. Bull. Environ. Contam. Toxicol. 1980, 25, 283–288. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Yang, G.; Cao, X.; Lin, Z. Photochemical degradation of triadimefon in seawater. J. Ocean. Univ. China 2008, 7, 171–176. [Google Scholar] [CrossRef]
- Zhang, W.; Lu, Y.; Huang, L.; Cheng, C.; Di, S.; Chen, L.; Zhou, Z.; Diao, J. Comparison of triadimefon and its metabolite on acute toxicity and chronic effects during the early development of Rana nigromaculata tadpoles. Ecotoxicol. Environ. Saf. 2018, 156, 247–254. [Google Scholar] [CrossRef]
- Cloern, J.E.; Jassby, A.D.; Schraga, T.S.; Nejad, E.; Martin, C. Ecosystem variability along the estuarine salinity gradient: Examples from long-term study of San Francisco Bay. Limnol. Oceanogr. 2017, 62, S272–S291. [Google Scholar] [CrossRef] [Green Version]
- National Research Council. Toxicity Testing in the 21st Century: A Vision and a Strategy; The National Academies Press: Washington, DC, USA, 2007. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Chemical | Conc. 1 mg/L | Conc. 2 mg/L | Conc. 3 mg/L | Conc. 4 mg/L | Conc. 5 mg/L | log KOW * | Water Solubility mg/L * |
|---|---|---|---|---|---|---|---|
| Bifenthrin | 0.00001 | 0.0001 | 0.0005 | 0.001 | 0.01 | 6.6 | 0.1 |
| Chlorpyrifos | 0.00005 | 0.0005 | 0.005 | 0.05 | 0.5 | 4.7 | 1.4 |
| Dicloran | 0.0009 | 0.009 | 0.09 | 0.9 | 9.0 | 2.8 | 6.3 |
| Myclobutanil | 0.004 | 0.04 | 0.4 | 4 | 40 | 2.8 | 132 |
| Paraquat | 0.067 | 0.67 | 6.7 | 67 | 670 | −4.5 | 6.20 × 105 |
| Penconazole | 0.00107 | 0.0107 | 0.107 | 1.07 | 10.7 | 3.7 | 73 |
| Triadimefon | 0.01 | 0.1 | 1 | 10 | 100 | 3.2 | 64 |
| Chemical | Nominal Concentration | Sample 1 | Sample 2 | Sample 3 |
|---|---|---|---|---|
| Bifenthrin | 0.0005 | 0.00030 ± 0.00000985 | 0.00030 ± 0.00000465 | 0.00027 ± 0.00000312 |
| Chlorpyrifos | 0.005 | 0.00282 ± 0.000400 | NA | |
| Dicloran | 0.09 | 0.0830 ± 0.0000676 | 0.079 ± 0.00014 | |
| Myclobutanil | 0.4 | 0.493 ± 0.00550 | 0.49 ± 0.0021 | |
| Paraquat | 6.7 | 6.61 ± 0.0150 | 7.19 ± 0.38 | |
| Penconazole | 0.107 | 0.0966 ± 0.00130 | 0.112 ± 0.0012 | |
| Triadimefon | 1.0 | 0.966 ± 0.00235 | 0.896 ± 0.00156 |
| Chemical | 5 PSU LC10 | 15 PSU LC10 | 5 PSU LC50 | 15 PSU LC50 | FW LC50 * |
|---|---|---|---|---|---|
| Bifenthrin | 0.0000227 | 0.00000533 | 0.000160 | 0.000120 | 0.00015–0.0147 |
| Chlorpyrifos | 0.000137 | 0.0000393 | 0.00803 | 0.00240 | <0.001–0.301 |
| Dicloran | 0.000288 | 0.0000704 | 0.00617 | 0.00224 | 0.56–1.6 |
| Myclobutanil | 0.00186 | 0.00430 | 3.76 | 1.55 | 4.2–5.27 |
| Paraquat | 0.0547 | 0.0245 | 37.50 | 18.14 | 15.0–38.68 |
| Penconazole | 0.000235 | 0.0000244 | 0.107 | 0.0202 | NA |
| Triadimefon | 0.118 | 0.0123 | 2.74 a | 0.218 a | 4.1–15.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hutton, S.J.; St. Romain, S.J.; Pedersen, E.I.; Siddiqui, S.; Chappell, P.E.; White, J.W.; Armbrust, K.L.; Brander, S.M. Salinity Alters Toxicity of Commonly Used Pesticides in a Model Euryhaline Fish Species (Menidia beryllina). Toxics 2021, 9, 114. https://doi.org/10.3390/toxics9050114
Hutton SJ, St. Romain SJ, Pedersen EI, Siddiqui S, Chappell PE, White JW, Armbrust KL, Brander SM. Salinity Alters Toxicity of Commonly Used Pesticides in a Model Euryhaline Fish Species (Menidia beryllina). Toxics. 2021; 9(5):114. https://doi.org/10.3390/toxics9050114
Chicago/Turabian StyleHutton, Sara J., Scott J. St. Romain, Emily I. Pedersen, Samreen Siddiqui, Patrick E. Chappell, J. Wilson White, Kevin L. Armbrust, and Susanne M. Brander. 2021. "Salinity Alters Toxicity of Commonly Used Pesticides in a Model Euryhaline Fish Species (Menidia beryllina)" Toxics 9, no. 5: 114. https://doi.org/10.3390/toxics9050114
APA StyleHutton, S. J., St. Romain, S. J., Pedersen, E. I., Siddiqui, S., Chappell, P. E., White, J. W., Armbrust, K. L., & Brander, S. M. (2021). Salinity Alters Toxicity of Commonly Used Pesticides in a Model Euryhaline Fish Species (Menidia beryllina). Toxics, 9(5), 114. https://doi.org/10.3390/toxics9050114