
Comparative Cytotoxicity Study of PM2.5 and TSP Collected from Urban Areas

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
3. Results
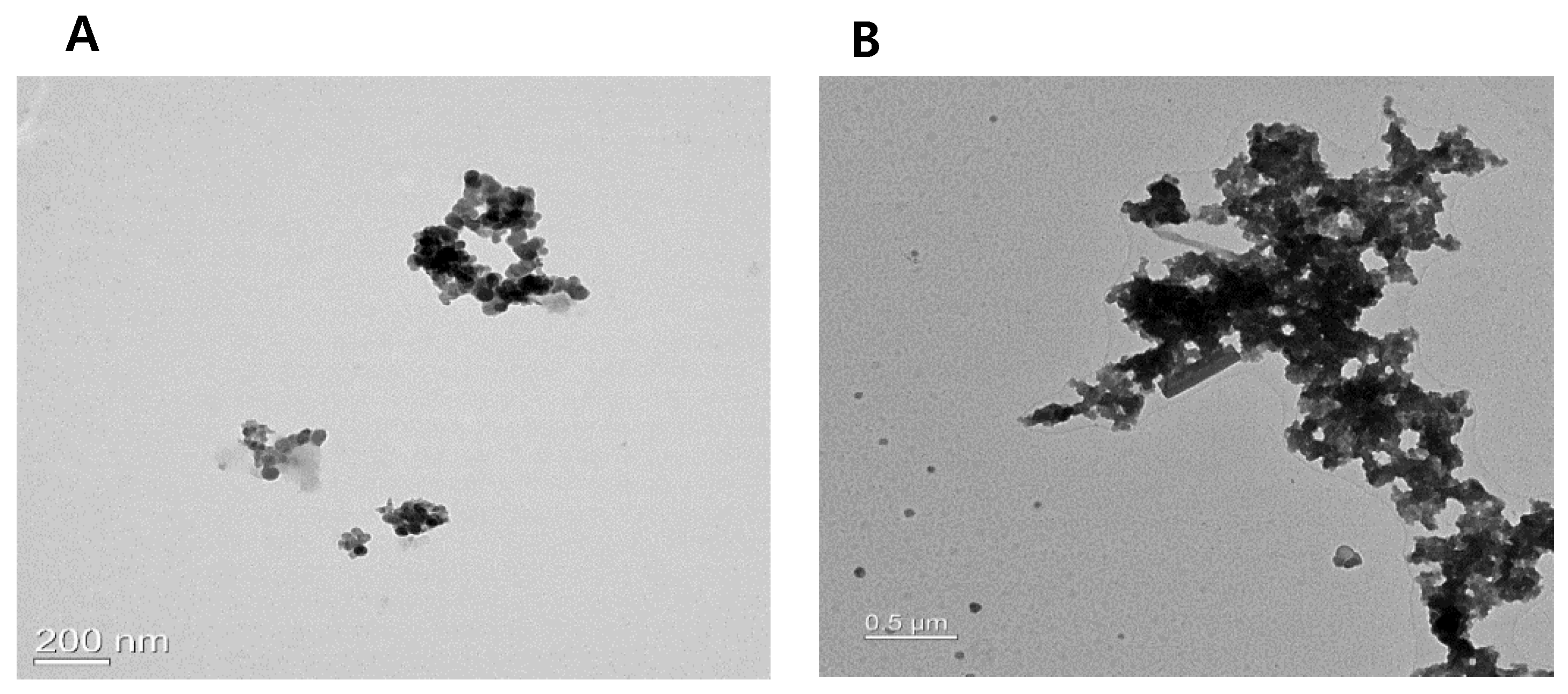
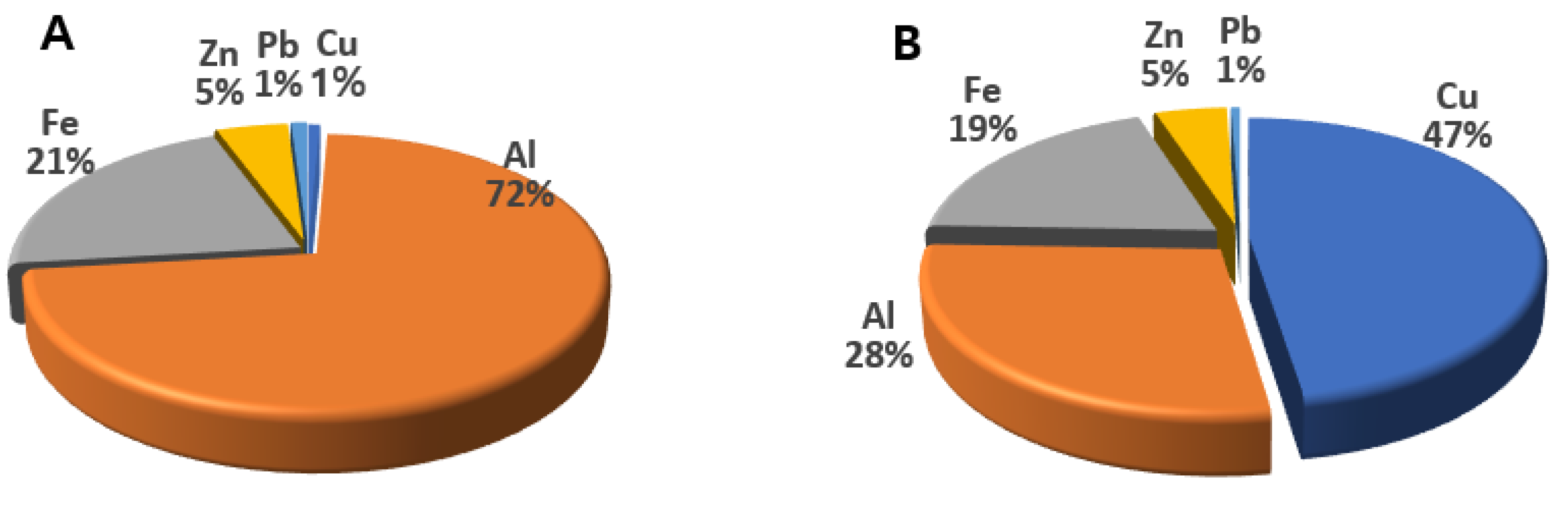
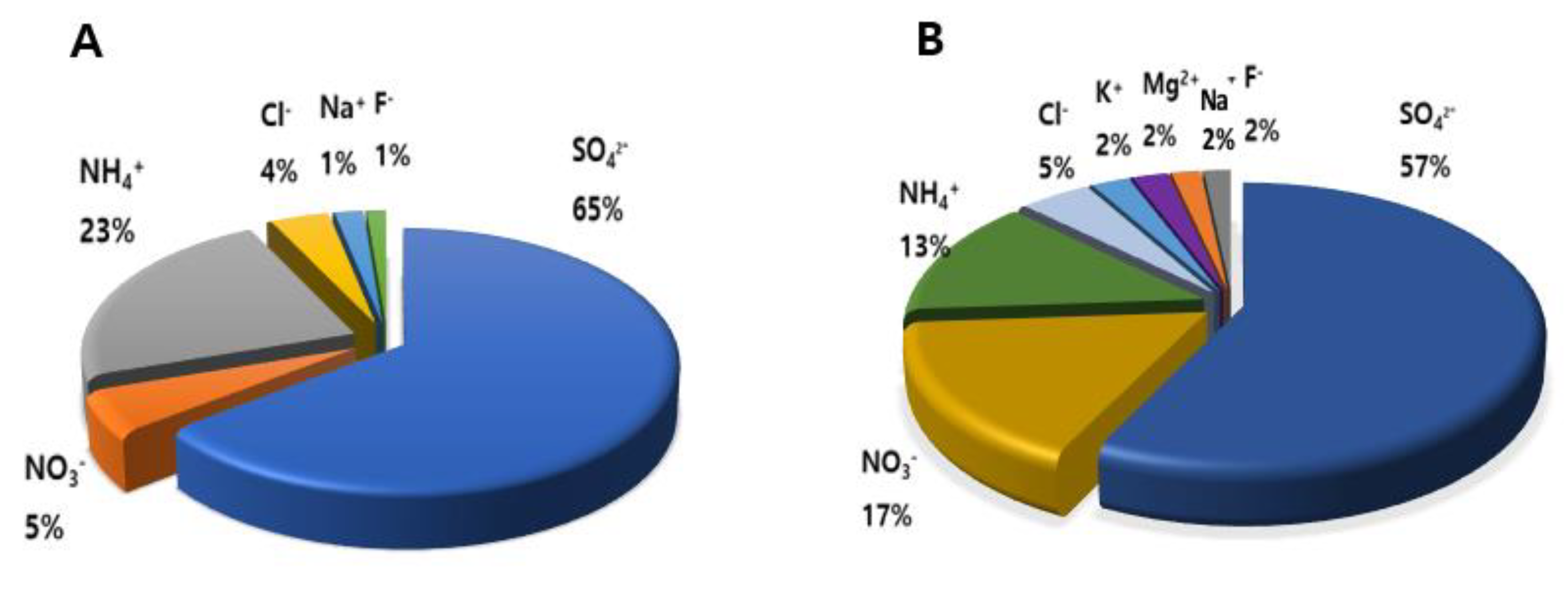
3.1. Analysis of PM2.5 and TSP
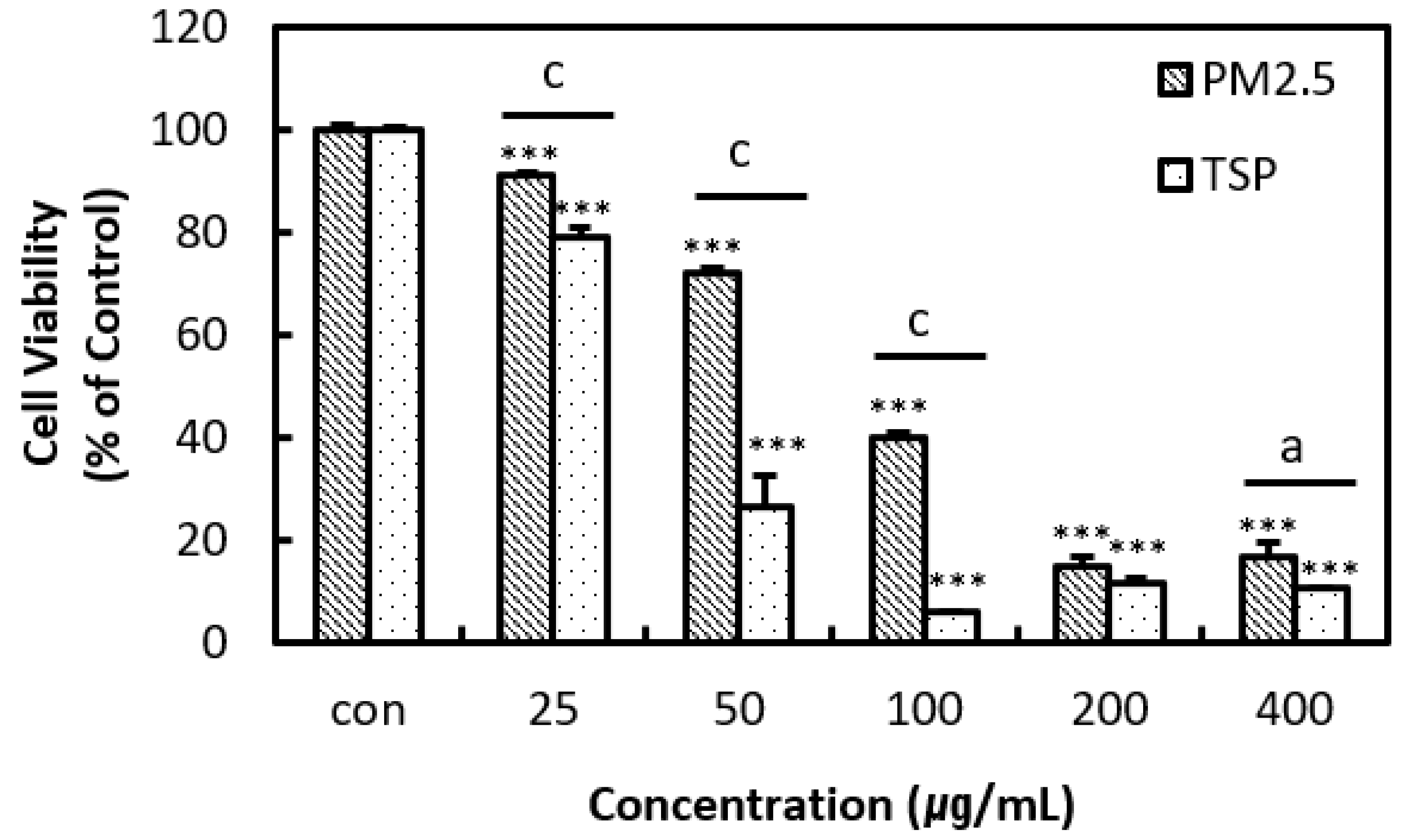
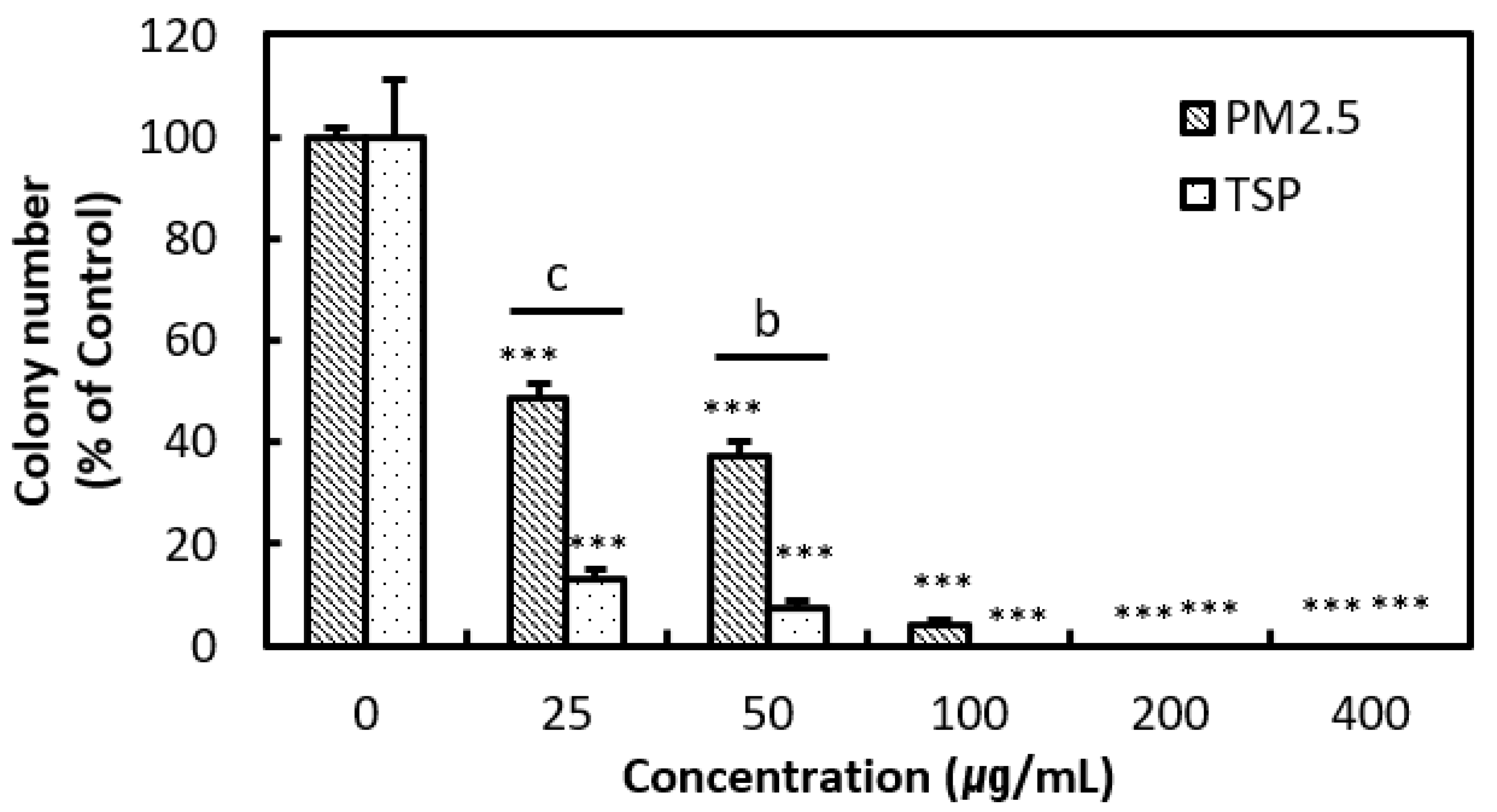
3.2. In Vitro Assay
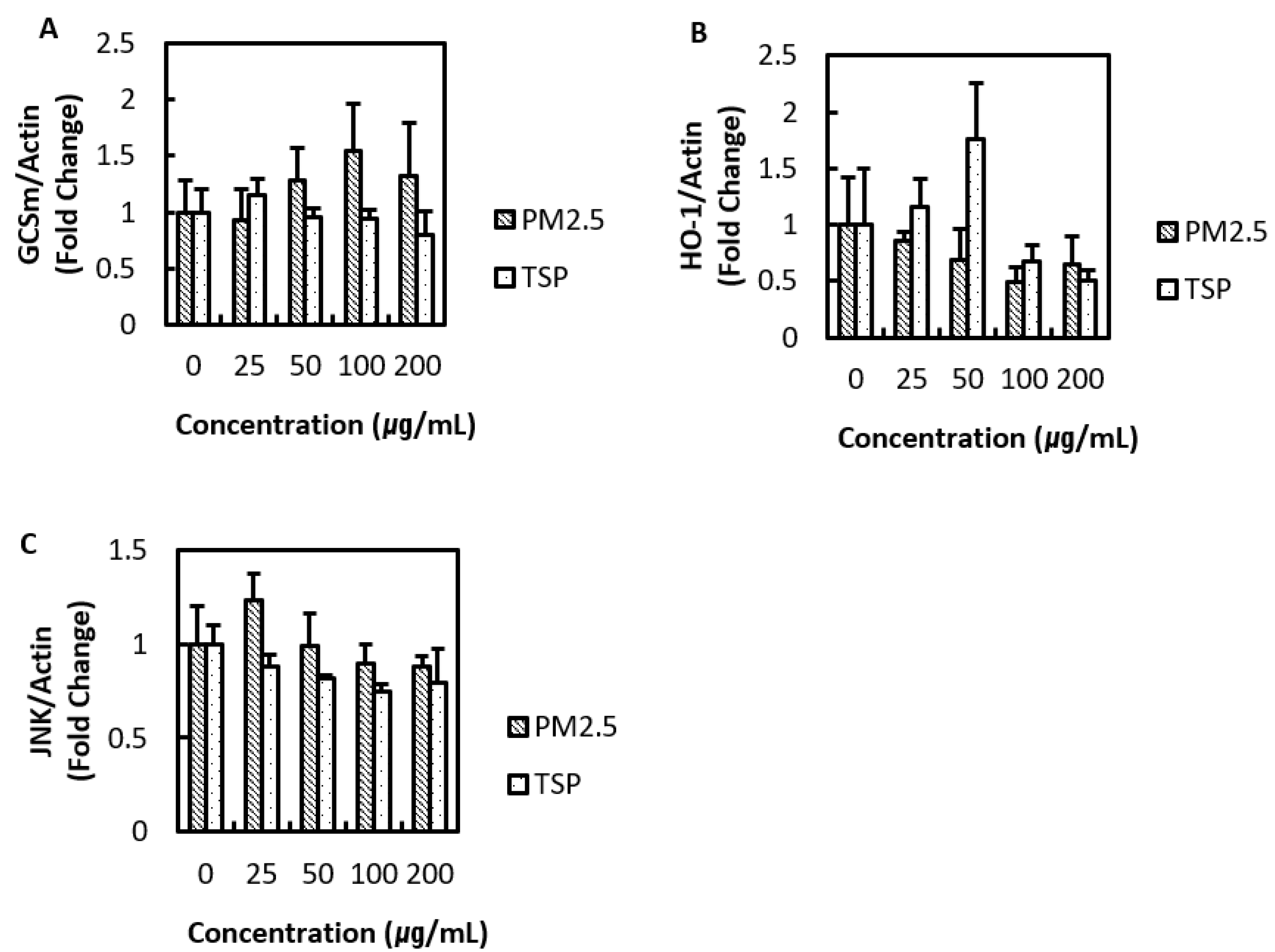
3.3. Oxidative Stress-Related Proteins
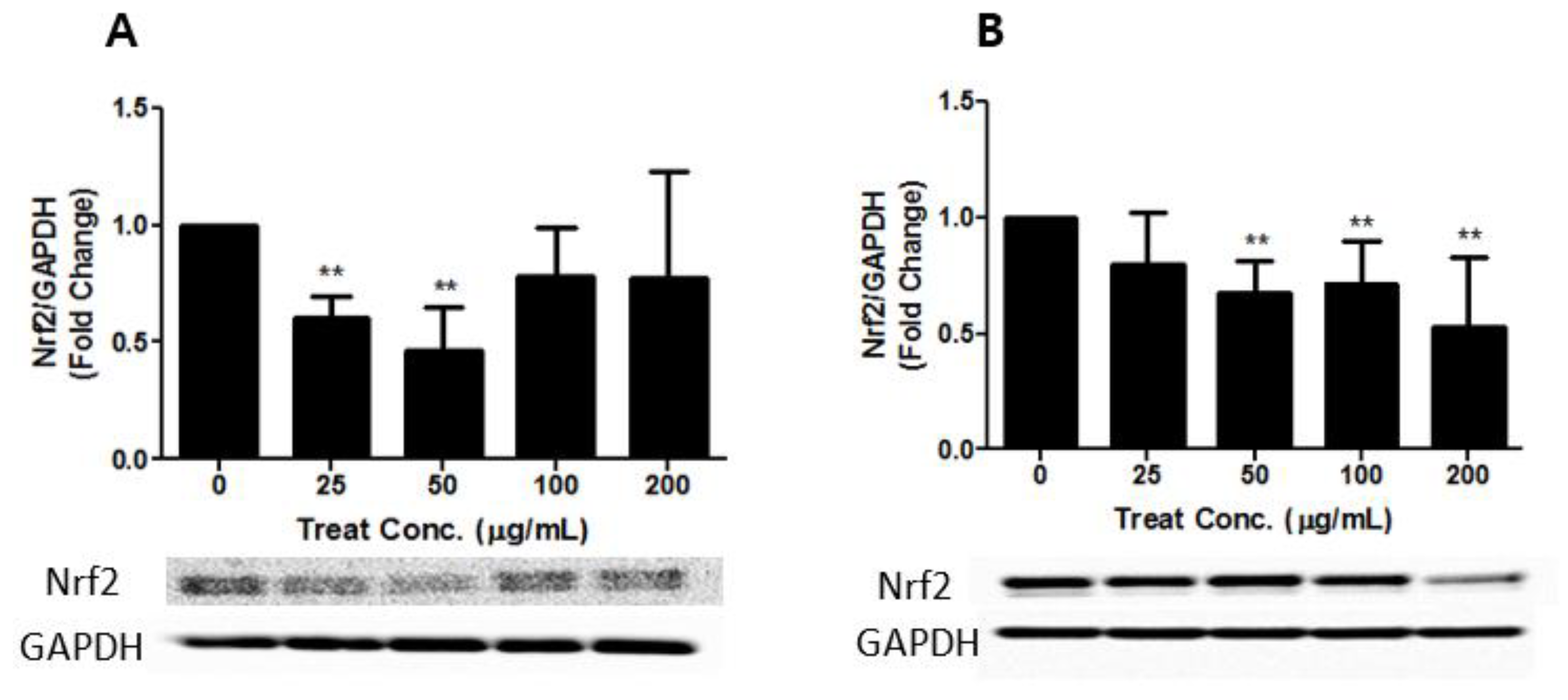
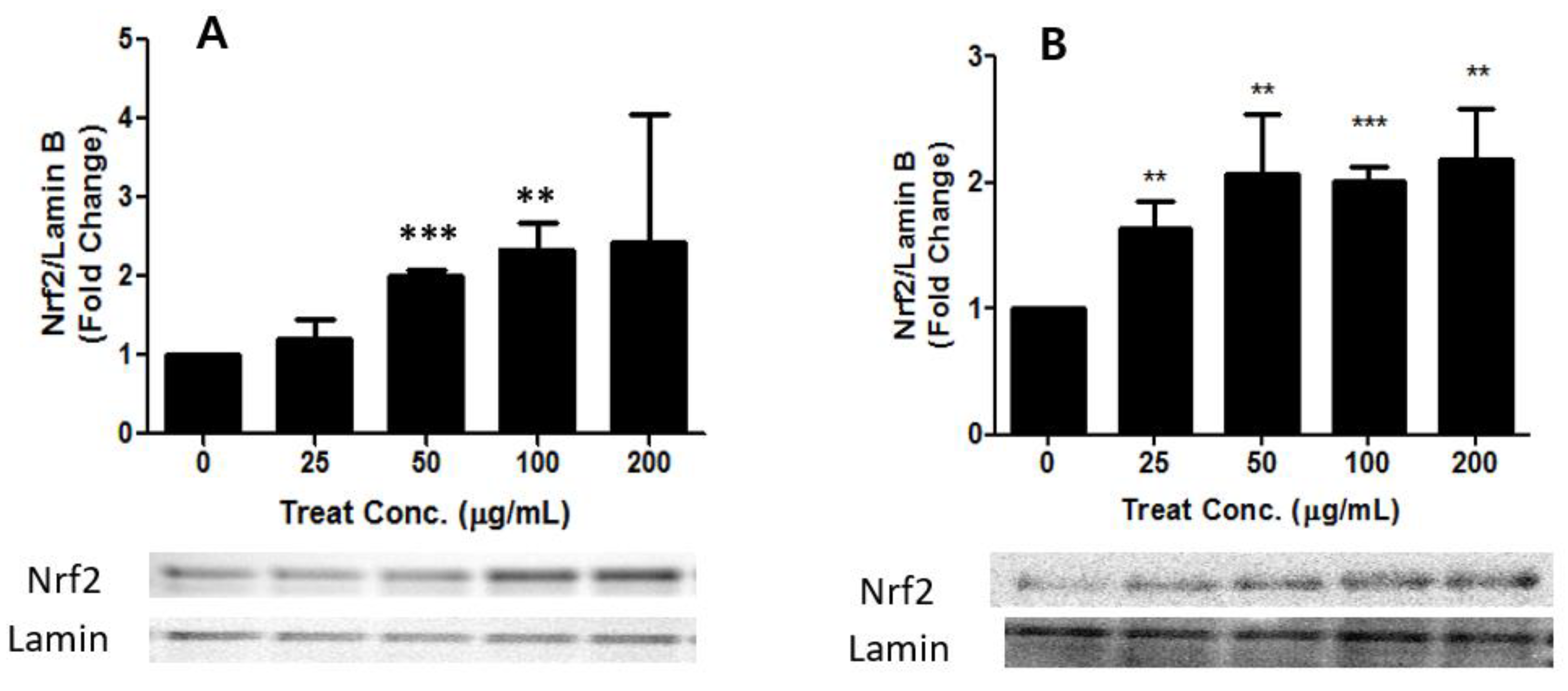
3.4. Effects of PM2.5 and TSP on the Keap1-Nrf2 Signaling Pathway
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Li, N.; Hao, M.; Phalen, R.F.; Hinds, W.C.; Nel, A.E. Particulate air pollutants and asthma; A paradigm for the role of oxidative stress in PM induced adverse health effects. Clin. Immunol. 2003, 109, 250–265. [Google Scholar] [CrossRef]
- Mills, N.L.; Donaldson, K.; Hadoke, P.W.; Boon, N.A.; MacNee, W.; Cassee, F.; Sandstrom, T.; Blomberg, A.; Newby, D.E. Adverse cardiovascular effects of air pollution. Nat. Rev. Cardiol. 2009, 6, 36. [Google Scholar] [CrossRef]
- Qing, H.; Wang, X.; Zhang, N.; Zheng, K.; Du, K.; Zheng, M.; Li, Y.; Chang, Y.; Zhang, L.; Bachert, C. The Effect of Fine Particulate Matter on the Inflammatory Responses in Human Upper Airway Mucosa. Am. J. Respir. Crit. Care Med. 2019, 200, 1315–1318. [Google Scholar] [CrossRef]
- Riediker, M.; Zink, D.; Kreyling, W.; Oberdorster, G.; Elder, A.; Graham, U.; Lynch, I.; Duschl, A.; Ichihara, G.; Ichihara, S.; et al. Particle toxicology and health—Where are we? Part. Fibre Toxicol. 2019, 16, 19. [Google Scholar] [CrossRef] [PubMed]
- Li, M.-H.; Fan, L.-C.; Mao, B.; Yang, J.-W.; Choi, A.M.K.; Cao, W.-J.; Xu, J.-F. Short-term exposure to ambient fine particulate matter increases hospitalizations and mortality in COPD: A systematic review and meta-analysis. Chest 2016, 149, 447–458. [Google Scholar] [CrossRef]
- Valavanidis, A.; Fiotakis, K.; Vlachogianni, T. Airborne particulate matter and human health: Toxicological assessment and importance of size and composition of particles for oxidative damage and carcinogenic mechanisms. J. Environ. Sci. Health Part C 2008, 26, 339–362. [Google Scholar] [CrossRef] [PubMed]
- Saikia, J.; Narzry, B.; Roy, S.; Bordoloi, M.; Saikia, P.; Saikia, B.K. Nanominerals, fullerene aggregates, and hazardous elements in coal and coal combustion-generated aerosols: An environmental and toxicological assessment. Chemosphere 2016, 164, 84–91. [Google Scholar] [CrossRef]
- Islam, N.; Dihingia, A.; Manna, P.; Das, T.; Kalita, J.; Dekaboruah, H.P.; Saikia, B.K. Environmental and toxicological assessment of nanodiamond-like materials derived from carbonaceous aerosols. Sci. Total Environ. 2019, 679, 209–220. [Google Scholar] [CrossRef]
- Xiao, X.; Wang, R.; Cao, L.; Shen, Z.-X.; Cao, Y.-X. The Role of MAPK Pathways in Airborne Fine Particulate Matter-Induced Upregulation of Endothelin Receptors in Rat Basilar Arteries. Toxicol. Sci. 2016, 149, 213–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rico, D.; Gonzalez, A.M.; Diaz, S.; Lucas, P.; Gutierrez, J.-C. Heavy metals generate reactive oxygen species in terrestrial and aquatic ciliated protozoa. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2009, 149, 90–96. [Google Scholar] [CrossRef]
- Shahid, M.; Pourrut, B.; Dumat, C.; Nadeem, M.; Aslam, M.; Pinelli, E. Heavy-metal-induced reactive oxygen species: Phytotoxicity and physicochemical changes in plants. In Reviews of Environmental Contamination and Toxicology; Springer: Berlin/Heidelberg, Germany, 2014; Volume 232, pp. 1–44. [Google Scholar]
- Lee, J.-C.; Son, Y.-O.; Pratheeshkumar, P.; Shi, X. Oxidative stress and metal carcinogenesis. Free Radic. Biol. Med. 2012, 53, 742–757. [Google Scholar] [CrossRef]
- Risom, L.; Møller, P.; Loft, S. Oxidative stress-induced DNA damage by particulate air pollution. Mutat. Res./Fundam. Mol. Mech. Mutagenesis 2005, 592, 119–137. [Google Scholar] [CrossRef]
- Cho, A.K.; Sioutas, C.; Miguel, A.H.; Kumagai, Y.; Schmitz, D.A.; Singh, M.; Fernandez, A.E.; Froines, J.R. Redox activity of airborne particulate matter at different sites in the Los Angeles Basin. Environ. Res. 2005, 99, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Valko, M.; Leibfritz, D.; Moncol, J.D.; Moncol, J.; Cronin, M.T.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef] [PubMed]
- Leetham, M.; DeWitt, B.; Buck, B.; Goossens, D.; Teng, Y.; Pollard, J.; McLaurin, B.; Gerads, R.; Keil, D. Oxidative stress and lung pathology following geogenic dust exposure. J. Appl. Toxicol. 2016, 36, 1276–1283. [Google Scholar] [CrossRef] [PubMed]
- Xia, T.; Kovochich, M.; Nel, A. The role of reactive oxygen species and oxidative stress in mediating particulate matter injury. Clin. Occup. Environ. Med. 2006, 5, 817–836. [Google Scholar]
- Li, Y.-J.; Kawada, T.; Azuma, A. Nrf2 is a protective factor against oxidative stresses induced by diesel exhaust particle in allergic asthma. Oxidative Med. Cell Longev. 2013, 2013, 323607. [Google Scholar] [CrossRef] [Green Version]
- Shim, I.; Seo, G.-B.; Oh, E.; Lee, M.; Kwon, J.-T.; Sul, D.; Lee, B.-W.; Yoon, B.-I.; Kim, P.; Choi, K.; et al. Inhalation exposure to chloramine T induces DNA damage and inflammation in lung of Sprague-Dawley rats. J. Toxicol. Sci. 2013, 38, 937–946. [Google Scholar] [CrossRef] [Green Version]
- Jung, Y.S.; Kim, S.J.; Kwon, D.Y.; Jun, D.S.; Kim, Y.C. Significance of alterations in the metabolomics of sulfur-containing amino acids during liver regeneration. Biochimie 2013, 95, 1605–1610. [Google Scholar] [CrossRef]
- Shim, I.; Kim, H.; Yang, S.; Choi, M.; Seo, G.; Lee, B.-W.; Yoon, B.-I.; Kim, P.; Choi, K. Inhalation of talc induces infiltration of macrophages and upregulation of manganese superoxide dismutase in rats. Int. J. Toxicol. 2015, 34, 491–499. [Google Scholar] [CrossRef]
- Nguyen, T.; Nioi, P.; Pickett, C.B. The Nrf2-antioxidant response element signaling pathway and its activation by oxidative stress. J. Biol. Chem. 2009, 284, 13291–13295. [Google Scholar] [CrossRef] [Green Version]
- Natsch, A. The Nrf2-Keap1-ARE toxicity pathway as a cellular sensor for skin sensitizers—functional relevance and a hypothesis on innate reactions to skin sensitizers. Toxicol. Sci. 2010, 113, 284–292. [Google Scholar] [CrossRef] [Green Version]
- Rychlik, K.A.; Secrest, J.R.; Lau, C.; Pulczinski, J.; Zamora, M.L.; Leal, J.; Langley, R.; Myatt, L.G.; Raju, M.; Chang, R.C.; et al. In utero ultrafine particulate matter exposure causes offspring pulmonary immunosuppression. Proc. Natl. Acad. Sci. USA 2019, 116, 3443–3448. [Google Scholar] [CrossRef] [Green Version]
- Deng, X.; Rui, W.; Zhang, F.; Ding, W. PM 2.5 induces Nrf2-mediated defense mechanisms against oxidative stress by activating PIK3/AKT signaling pathway in human lung alveolar epithelial A549 cells. Cell Biol. Toxicol. 2013, 29, 143–157. [Google Scholar] [CrossRef]
- Simmons, S.O.; Fan, C.-Y.; Yeoman, K.; Wakefield, J.; Ramabhadran, R. NRF2 oxidative stress induced by heavy metals is cell type dependent. Curr. Chem. Genom. 2011, 5, 1. [Google Scholar] [CrossRef] [Green Version]
- Schopfer, F.J.; Cipollina, C.; Freeman, B.A. Formation and signaling actions of electrophilic lipids. Chem. Rev. 2011, 111, 5997–6021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toyama, T.; Sumi, D.; Shinkai, Y.; Yasutake, A.; Taguchi, K.; Tong, K.I.; Yamamoto, M.; Kumagai, Y. Cytoprotective role of Nrf2/Keap1 system in methylmercury toxicity. Biochem. Biophys. Res. Commun. 2007, 363, 645–650. [Google Scholar] [CrossRef] [PubMed]
- Uruno, A.; Matsumaru, D.; Ryoke, R.; Saito, R.; Kadoguchi, S.; Saigusa, D.; Saito, T.; Saido, T.C.; Kawashima, R.; Yamamoto, M. Nrf2 Suppresses Oxidative Stress and Inflammation in App Knock-in Alzheimer’s Disease Model Mice. Mol. Cell. Biol. 2020, 40, 6. [Google Scholar] [CrossRef] [PubMed]
- Sawa, T.; Zaki, M.H.; Okamoto, T.; Akuta, T.; Tokutomi, Y.; Kim-Mitsuyama, S.; Ihara, H.; Kobayashi, A.; Yamamoto, M.; Fujii, S.; et al. Protein S-guanylation by the biological signal 8-nitroguanosine 3′,5′-cyclic monophosphate. Nat. Chem. Biol. 2007, 3, 727. [Google Scholar] [CrossRef] [Green Version]
- Shang, Y.; Zhang, L.; Jiang, Y.; Li, Y.; Lu, P. Airborne quinones induce cytotoxicity and DNA damage in human lung epithelial A549 cells: The role of reactive oxygen species. Chemosphere 2014, 100, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Gu, C.; He, W.; Ye, X.; Chen, H.; Zhang, X.; Hai, C. Glucose oxidase induces insulin resistance via influencing multiple targets in vitro and in vivo: The central role of oxidative stress. Biochimie 2012, 94, 1705–1717. [Google Scholar] [CrossRef]
- Gualtieri, M.; Ovrevik, J.; Holme, J.A.; Perrone, M.G.; Bolzacchini, E.; Schwarze, P.E.; Camatini, M. Differences in cytotoxicity versus pro-inflammatory potency of different PM fractions in human epithelial lung cells. Toxicol. Vitr. 2010, 24, 29–39. [Google Scholar] [CrossRef]
- Cachon, B.F.; Frimin, S.; Verdin, A.; Ayi-Fanou, L.; Billet, S.; Cazier, F.; Martin, P.J.; Aissi, F.; Courcot, D.; Sanni, A.; et al. Proinflammatory effects and oxidative stress within human bronchial epithelial cells exposed to atmospheric particulate matter (PM 2.5 and PM > 2.5) collected from Cotonou, Benin. Environ. Pollut. 2014, 185, 340–351. [Google Scholar] [CrossRef]
- Valko, M.; Rhodes, C.J.; Moncol, J.; Izakovic, M.; Mazur, M. Free radicals, metals and antioxidants in oxidative stress-induced cancer. Chem. Biol. Interact. 2006, 160, 1–40. [Google Scholar] [CrossRef]
- Fu, Z.; Xi, S. The effects of heavy metals on human metabolism. Toxicol. Mech. Methods 2020, 30, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Matés, J.M.; Segura, J.A.; Alonso, F.J.; Marquez, J. Roles of dioxins and heavy metals in cancer and neurological diseases using ROS-mediated mechanisms. Free Radic. Biol. Med. 2010, 49, 1328–1341. [Google Scholar] [CrossRef]
- Islam, N.; Dihingia, A.; Khare, P.; Saikia, B.K. Atmospheric particulate matters in an Indian urban area: Health implications from potentially hazardous elements, cytotoxicity, and genotoxicity studies. J. Hazard. Mater. 2020, 384, 121472. [Google Scholar] [CrossRef]
- Maté, T.; Guaita, R.; Pichiule, M.; Linares, C.; Diaz, J. Short-term effect of fine particulate matter (PM 2.5) on daily mortality due to diseases of the circulatory system in Madrid (Spain). Sci. Total Environ. 2010, 408, 5750–5757. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Tu, Y.; Yu, Z.; Lu, R. PM 2.5 and cardiovascular diseases in the elderly: An overview. Int. J. Environ. Res. Public Health 2015, 12, 8187–8197. [Google Scholar] [CrossRef] [Green Version]
- Madureira, J.; Slezakova, K.; Silva, A.I.; Lage, B.; Mendes, A.; Aguiar, L.; Pereira, M.C.; Teixeira, J.P.; Costa, C. Assessment of indoor air exposure at residential homes: Inhalation dose and lung deposition of PM 10, PM 2.5 and ultrafine particles among newborn children and their mothers. Sci. Total Environ. 2020, 717, 137293. [Google Scholar] [CrossRef] [PubMed]
- MohanKumar, S.M.; Campbell, A.; Block, M.; Veronesi, B. Particulate matter, oxidative stress and neurotoxicity. Neurotoxicology 2008, 29, 479–488. [Google Scholar] [CrossRef]
- Xu, X.; Liu, C.; Xu, Z.; Tzan, K.; Zhong, M.; Wang, Z.; Lippmann, M.; Chen, L.-C.; Rajagopalan, S.; Sun, Q. Long-term exposure to ambient fine particulate pollution induces insulin resistance and mitochondrial alteration in adipose tissue. Toxicol. Sci. 2011, 124, 88–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, S.; Gao, D.; Liao, F.; Zhou, F.; Wang, X. The health effects of ambient PM 2.5 and potential mechanisms. Ecotoxicol. Environ. Saf. 2016, 128, 67–74. [Google Scholar] [CrossRef]
- Niu, X.; Chuang, H.-C.; Wang, X.; Ho, S.S.H.; Li, L.; Qu, L.; Chow, J.C.; Watson, J.G.; Sun, J.; Lee, S.; et al. Cytotoxicity of PM 2.5 vehicular emissions in the Shing Mun Tunnel, Hong Kong. Environ. Pollut. 2020, 263, 114386. [Google Scholar] [CrossRef]
- Yan, Z.; Wang, J.; Li, J.; Jiang, N.; Zhang, R.; Yang, W.; Yao, W.; Wu, W. Oxidative stress and endocytosis are involved in upregulation of interleukin-8 expression in airway cells exposed to PM 2.5. Environ. Toxicol. 2016, 31, 1869–1878. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.C.; Cho, Y.; Song, M.-K.; Lee, E.; Ryu, J.-C. Epidermal growth factor receptor (EGFR)—MAPK—nuclear factor (NF)-κB—IL8: A possible mechanism of particulate matter (PM) 2.5-induced lung toxicity. Environ. Toxicol. 2017, 32, 1628–1636. [Google Scholar] [CrossRef] [PubMed]
- Kaneto, H.; Matsuoka, T.; Nakatani, Y.; Kawamori, D.; Miyatsuka, T.; Matsuhisa, M.; Yamasaki, Y. Oxidative stress, ER stress, and the JNK pathway in type 2 diabetes. J. Mol. Med. 2005, 83, 429–439. [Google Scholar] [CrossRef] [PubMed]
- Shim, I.; Choi, K.; Hirano, S. Oxidative stress and cytotoxic effects of silver ion in mouse lung macrophages J774. 1 cells. J. Appl. Toxicol. 2017, 37, 471–478. [Google Scholar] [CrossRef]
- Zhang, T.; Tang, M.; Kong, L.; Li, H.; Zhang, T.; Xue, Y.; Pu, Y. Surface modification of multiwall carbon nanotubes determines the pro-inflammatory outcome in macrophage. J. Hazard. Mater. 2015, 284, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Shi, Q.; Zhang, L.; Zhao, H. High molecular weight hyaluronan attenuates fine particulate matter-induced acute lung injury through inhibition of ROS-ASK1-p38/JNK-mediated epithelial apoptosis. Environ. Toxicol. Pharmacol. 2018, 59, 190–198. [Google Scholar] [CrossRef]
- Biswas, C.; Shah, N.; Muthu, M.; La, P.; Fernando, A.P.; Sengpta, S.; Yang, G.; Dennery, P.A. Nuclear heme oxygenase-1 (HO-1) modulates subcellular distribution and activation of Nrf2, impacting metabolic and anti-oxidant defenses. J. Biol. Chem. 2014, 289, 26882–26894. [Google Scholar] [CrossRef] [Green Version]
- Iida, K.; Itoh, K.; Kumagai, Y.; Oyasu, R.; Hattori, K.; Kawai, K.; Shimazui, T.; Akaza, H.; Yamamoto, M. Nrf2 is essential for the chemopreventive efficacy of oltipraz against urinary bladder carcinogenesis. Cancer Res. 2004, 64, 6424–6431. [Google Scholar] [CrossRef] [Green Version]
- Loboda, A.; Damulewicz, M.; Pyza, E.; Jozkowicz, A.; Dulak, J. Role of Nrf2/HO-1 system in development, oxidative stress response and diseases: An evolutionarily conserved mechanism. Cell Mol. Life Sci. 2016, 73, 3221–3247. [Google Scholar] [CrossRef] [Green Version]
- Hiramatsu, K.; Tsuneyoshi, T.; Ogawa, T.; Morihara, N. Aged garlic extract enhances heme oxygenase-1 and glutamate-cysteine ligase modifier subunit expression via the nuclear factor erythroid 2–related factor 2–antioxidant response element signaling pathway in human endothelial cells. Nutr. Res. 2016, 36, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Bekki, K.; Ito, T.; Yoshida, Y.; He, C.; Arashidani, K.; He, M.; Sun, G.; Zeng, Y.; Sone, H.; Kunugita, N.; et al. PM 2.5 collected in China causes inflammatory and oxidative stress responses in macrophages through the multiple pathways. Environ. Toxicol. Pharmacol. 2016, 45, 362–369. [Google Scholar] [CrossRef]
- Deng, X.; Zhang, F.; Rui, W.; Long, F.; Wang, L.; Feng, Z.; Chen, D.; Ding, W. PM 2.5-induced oxidative stress triggers autophagy in human lung epithelial A549 cells. Toxicol. Vitr. 2013, 27, 1762–1770. [Google Scholar] [CrossRef]
- Choi, B.-H.; Kwak, M.-K. Shadows of NRF2 in cancer: Resistance to chemotherapy. Curr. Opin. Toxicol. 2016, 1, 20–28. [Google Scholar] [CrossRef]
- Suzuki, M.; Otsuki, A.; Keleku-Lukwete, N.; Yamamoto, M. Overview of redox regulation by Keap1–Nrf2 system in toxicology and cancer. Curr. Opin. Toxicol. 2016, 1, 29–36. [Google Scholar] [CrossRef]
- Rudolph, T.K.; Freeman, B.A. Transduction of redox signaling by electrophile-protein reactions. Sci. Signal. 2009, 2, re7. [Google Scholar] [CrossRef] [Green Version]
- Shinkai, Y.; Kaji, T. Cellular defense mechanisms against lead toxicity in the vascular system. Biol. Pharm. Bull. 2012, 35, 1885–1891. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Qian, W.; Wang, D.; Meng, Y.; Wang, X.; Chen, Y.; Li, X.; Xie, C.; Zhong, C.; Fu, S. Resveratrol relieves particulate matter (mean diameter < 2.5 μm)-induced oxidative injury of lung cells through attenuation of autophagy deregulation. J. Appl. Toxicol. 2018, 38, 1251–1261. [Google Scholar] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Elements | PM2.5 (ng/m3) | TSP (ng/m3) |
|---|---|---|
| Cu | 9.26 ± 3.15 | 2572 ± 593 |
| Al | 899 ± 11 | 1520 ± 54 |
| Fe | 263 ± 30 | 1045 ± 32 |
| Zn | 57.3 ± 10.7 | 247.4 ± 88.5 |
| Pb | 12.96 ± 1.90 | 31.05 ± 17.51 |
| As | 3.10 ± 1.20 | 8.10 ± 5.65 |
| Cr | 3.89 ± 0.21 | 7.96 ± 2.52 |
| Cd | 0.45 ± 0.04 | 1.04 ± 0.44 |
| Ions | PM2.5 (μg/m3) | TSP (μg/m3) |
|---|---|---|
| SO42− | 8.091 ± 2.887 | 7.993 ± 2.102 |
| NO3− | 0.662 ± 0.219 | 2.319 ± 1.510 |
| NH4+ | 2.873 ± 0.920 | 1.872 ± 0.851 |
| Cl − | 0.431 ± 0.003 | 0.629 ± 0.003 |
| K+ | N.D. | 0.335 ± 0.049 |
| Mg2+ | N.D. | 0.317 ± 0.054 |
| Na+ | 0.243 ± 0.032 | 0.257 ± 0.048 |
| F− | 0.162 ± 0.003 | 0.236 ± 0.003 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shim, I.; Kim, W.; Kim, H.; Lim, Y.-M.; Shin, H.; Park, K.S.; Yu, S.M.; Kim, Y.H.; Sung, H.K.; Eom, I.-C.; et al. Comparative Cytotoxicity Study of PM2.5 and TSP Collected from Urban Areas. Toxics 2021, 9, 167. https://doi.org/10.3390/toxics9070167
Shim I, Kim W, Kim H, Lim Y-M, Shin H, Park KS, Yu SM, Kim YH, Sung HK, Eom I-C, et al. Comparative Cytotoxicity Study of PM2.5 and TSP Collected from Urban Areas. Toxics. 2021; 9(7):167. https://doi.org/10.3390/toxics9070167
Chicago/Turabian StyleShim, Ilseob, Woong Kim, Haewon Kim, Yeon-Mi Lim, Hyejung Shin, Kwang Su Park, Seok Min Yu, Young Hee Kim, Hwa Kyung Sung, Ig-Chun Eom, and et al. 2021. "Comparative Cytotoxicity Study of PM2.5 and TSP Collected from Urban Areas" Toxics 9, no. 7: 167. https://doi.org/10.3390/toxics9070167
APA StyleShim, I., Kim, W., Kim, H., Lim, Y. -M., Shin, H., Park, K. S., Yu, S. M., Kim, Y. H., Sung, H. K., Eom, I. -C., Kim, P., & Yu, S. -D. (2021). Comparative Cytotoxicity Study of PM2.5 and TSP Collected from Urban Areas. Toxics, 9(7), 167. https://doi.org/10.3390/toxics9070167