
Indirect Effect of Pesticides on Insects and Other Arthropods

{kind=link}
{kind=link}
Abstract
:1. Introduction
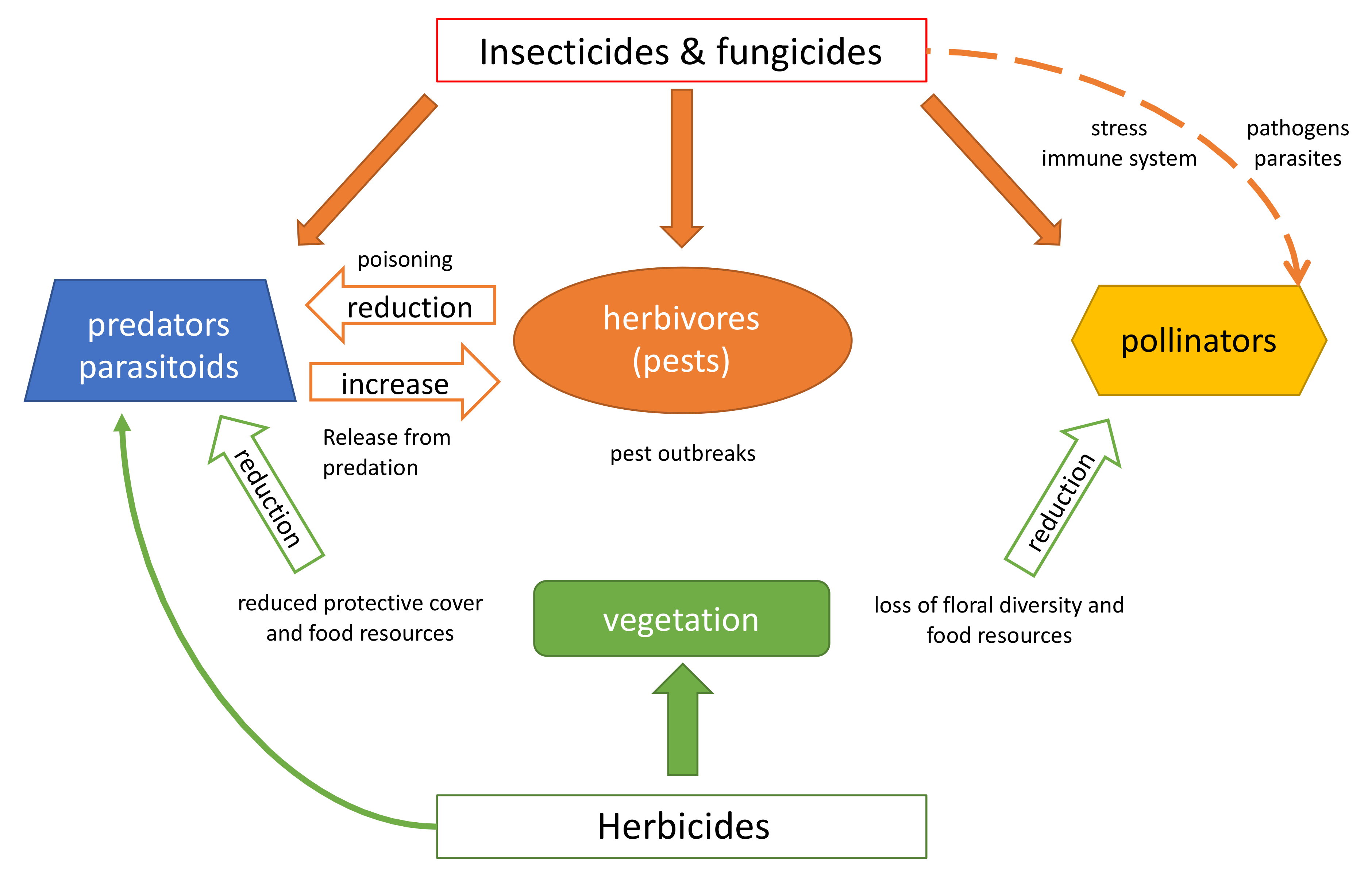
2. Indirect Effects in Terrestrial Ecosystems
2.1. Herbicides
2.2. Insecticides
2.2.1. Pest Resurgence
2.2.2. Outbreak of Secondary Pests
2.2.3. Secondary Poisoning of Natural Enemies
2.2.4. Pathogens and Diseases in Pollinators
2.3. Parasiticides
2.4. Fungicides
2.5. Pesticide Mixtures
3. Aquatic Ecosystems
3.1. Insecticides
3.2. Fungicides
3.3. Herbicides
3.4. Pesticide Mixtures
4. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bernhardt, E.S.; Rosi, E.J.; Gessner, M.O. Synthetic chemicals as agents of global change. Front. Ecol. Environ. 2017, 15, 84–90. [Google Scholar] [CrossRef]
- Beketov, M.A.; Kefford, B.J.; Schäfer, R.B.; Liess, M. Pesticides reduce regional biodiversity of stream invertebrates. Proc. Natl. Acad. Sci. USA 2013, 110, 11039–11043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habel, J.C.; Samways, M.J.; Schmitt, T. Mitigating the precipitous decline of terrestrial European insects: Requirements for a new strategy. Biodivers. Conserv. 2019, 28, 1343–1360. [Google Scholar] [CrossRef]
- Goulson, D.; Nicholls, E. The canary in the coalmine; bee declines as an indicator of environmental health. Science Progress 2016, 99, 312–326. [Google Scholar] [CrossRef] [PubMed]
- Raven, P.H.; Wagner, D.L. Agricultural intensification and climate change are rapidly decreasing insect biodiversity. Proc. Natl. Acad. Sci. USA 2021, 118, e2002548117. [Google Scholar] [CrossRef]
- Desneux, N.; Decourtye, A.; Delpuech, J.-M. The sublethal effects of pesticides on beneficial arthropods. Annu. Rev. Entomol. 2007, 52, 81–106. [Google Scholar] [CrossRef] [PubMed]
- Stehle, S.; Schulz, R. The Impact of Pesticides on Our Freshwater Resources. In Encyclopedia of the Anthropocene; Reference Module in Earth Systems and Environmental Sciences; Dellasala, D.A., Goldstein, M.I., Eds.; Elsevier: Oxford, UK, 2018; Volume 5, pp. 127–132. [Google Scholar]
- Katayama, N.; Baba, Y.G.; Kusumoto, Y.; Tanaka, K. A review of post-war changes in rice farming and biodiversity in Japan. Agric. Syst. 2015, 132, 73–84. [Google Scholar] [CrossRef]
- McLaughlin, A.; Mineau, P. The impact of agricultural practices on biodiversity. Agric. Ecosyst. Environ. 1995, 55, 201–212. [Google Scholar] [CrossRef]
- Brown, A.W.A. Ecology of Pesticides; John Wiley & Sons, Inc.: New York, NY, USA, 1978; p. 525. [Google Scholar]
- Relyea, R.A.; Hoverman, J.T. Interactive effects of predators and a pesticide on aquatic communities. Oikos 2008, 117, 1647–1658. [Google Scholar] [CrossRef]
- Rohr, J.R.; Crumrine, P.W. Effects of an herbicide and an insecticide on pond community structure and processes. Ecol. Appl. 2005, 15, 1135–1147. [Google Scholar] [CrossRef] [Green Version]
- Fleeger, J.W.; Carman, K.R.; Nisbet, R.M. Indirect effects of contaminants in aquatic ecosystems. Sci. Total Environ. 2003, 317, 207–233. [Google Scholar] [CrossRef]
- Guedes, R.N.C.; Smagghe, G.; Stark, J.D.; Desneux, N. Pesticide-induced stress in arthropod pests for optimized integrated pest management programs. Annu. Rev. Entomol. 2016, 61, 43–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clements, W.H.; Kiffney, P.M. Assessing contaminant effects at higher levels of biological organization. Environ. Toxicol. Chem. 1994, 13, 357–359. [Google Scholar] [CrossRef]
- Stark, J.D.; Banks, J.E. Population-level effects of pesticides and other toxicants on arthropods. Annu. Rev. Entomol. 2003, 48, 505–519. [Google Scholar] [CrossRef] [PubMed]
- Del Arco, A.I.; Parra, G.; Rico, A.; van den Brink, P.J. Effects of intra- and interspecific competition on the sensitivity of aquatic macroinvertebrates to carbendazim. Ecotoxicol. Environ. Saf. 2015, 120, 27–34. [Google Scholar] [CrossRef]
- Zhao, Q.; de Laender, F.; van den Brink, P.J. Community composition modifies direct and indirect effects of pesticides in freshwater food webs. Sci. Total Environ. 2020, 739, 139531. [Google Scholar] [CrossRef]
- Strong, D.R.; Lawton, J.H.; Southwood, T.R.E. Insects on plants: Community Patterns and Mechanisms; Blackwell Scientific: Oxford/Boston, MA, USA, 1984. [Google Scholar]
- Altieri, M.A. The ecological role of biodiversity in agroecosystems. Agric. Ecosyst. Environ. 1999, 74, 19–31. [Google Scholar] [CrossRef] [Green Version]
- Weber, D.C.; Mangan, F.X.; Ferro, D.N.; Marsh, H.V., Jr. Effect of weed abundance on European corn borer (Lepidoptera: Pyralidae) infestation of sweet corn. Environ. Entomol. 1990, 19, 1858–1865. [Google Scholar] [CrossRef]
- Pimentel, D. Species diversity and insect population outbreaks. Ann. Entomol. Soc. Am. 1961, 54, 76–86. [Google Scholar] [CrossRef]
- Geiger, F.; Bengtsson, J.; Berendse, F.; Weisser, W.W.; Emmerson, M.; Morales, M.B.; Ceryngier, P.; Liira, J.; Tscharntke, T.; Winqvist, C.; et al. Persistent negative effects of pesticides on biodiversity and biological control potential on European farmland. Basic Appl. Ecol. 2010, 11, 97–105. [Google Scholar] [CrossRef]
- Cohen, J.E.; Schoenly, K.; Heong, K.L.; Justo, H.; Arida, G.; Barrion, A.T.; Litsinger, J.A. A food web approach to evaluating the effect of insecticide spraying on insect pest population dynamics in a Philippine irrigated rice ecosystem. J. Appl. Ecol. 1994, 31, 747–763. [Google Scholar] [CrossRef] [Green Version]
- Tsutsui, M.H.; Kobayashi, K.; Miyashita, T. Temporal trends in arthropod abundances after the transition to organic farming in paddy fields. PLoS ONE 2018, 13, e0190946. [Google Scholar] [CrossRef] [Green Version]
- Downing, A.; DeVanna, K.; Rubeck-Schurtz, C.; Tuhela, L.; Grunkemeyer, H. Community and ecosystem responses to a pulsed pesticide disturbance in freshwater ecosystems. Ecotoxicology 2008, 17, 539–548. [Google Scholar] [CrossRef]
- Norris, R.F.; Kogan, M. Interactions between weeds, arthropod pests, and their natural enemies in managed ecosystems. Weed. Sci. 2000, 48, 94–158. [Google Scholar] [CrossRef]
- Freemark, K.; Boutin, C. Impacts of agricultural herbicide use on terrestrial wildlife in temperate landscapes: A review with special reference to North America. Agric. Ecosyst. Environ. 1995, 52, 67–91. [Google Scholar] [CrossRef]
- Gianessi, L.P. The increasing importance of herbicides in worldwide crop production. Pest Manag. Sci. 2013, 69, 1099–1105. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Jha, P.; Reddy, G.V.P. Multidimensional relationships of herbicides with insect-crop food webs. Sci. Total Environ. 2018, 643, 1522–1532. [Google Scholar] [CrossRef]
- Sotherton, N.W. The distribution and abundance of predatory arthropods overwintering on farmland. Ann. Appl. Biol. 1984, 105, 423–429. [Google Scholar] [CrossRef]
- Cameron, S.A.; Lozier, J.D.; Strange, J.P.; Koch, J.B.; Cordes, N.; Solter, L.F.; Griswold, T.L. Patterns of widespread decline in North American bumble bees. PNAS 2011, 108, 662–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zattara, E.E.; Aizen, M.A. Worldwide occurrence records suggest a global decline in bee species richness. One Earth 2021, 4, 114–123. [Google Scholar] [CrossRef]
- Kevan, P.G.; Clark, E.A.; Thomas, V.G. Insect pollinators and sustainable agriculture. Am. J. Altern. Agric. 1990, 5, 13–22. [Google Scholar] [CrossRef]
- Rands, S.A.; Whitney, H.M. Field margins, foraging distances and their impacts on nesting pollinator success. PLoS ONE 2011, 6, e25971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoyer, T.L.; Kok, L.T. Insect/plant interactions in integrating Trichosirocalus horridus (Coleoptera: Curculionidae) and 2,4-dichlorophenoxyacetic acid for Carduus thistle control. Environ. Entomol. 1987, 16, 864–868. [Google Scholar] [CrossRef]
- Kampfraath, A.A.; Giesen, D.; van Gestel, C.A.M.; Le Lann, C. Pesticide stress on plants negatively affects parasitoid fitness through a bypass of their phytophage hosts. Ecotoxicology 2017, 26, 383–395. [Google Scholar] [CrossRef] [PubMed]
- Klingauf, F.A. Are feasible methods for an ecological pest control in view? Ecol. Bull. 1988, 39, 74–81. [Google Scholar]
- Moffett, J.O.; Morton, H.L.; MacDonald, R.H. Toxicity of some herbicidal sprays to honey bees. J. Econ. Entomol. 1972, 65, 32–36. [Google Scholar] [CrossRef] [PubMed]
- Potts, G.R. The Partridge - Pesticides, Predation and Conservation; Collins: London, UK, 1986. [Google Scholar]
- Rands, M.R.W. The survival of gamebird (Galliformes) chicks in relation to pesticide use on cereals. Ibis 1986, 128, 57–64. [Google Scholar] [CrossRef]
- Aebischer, N.J. Assessing pesticide effects on non-target invertebrates using long-term monitoring and time-series modelling. Funct. Ecol. 1990, 4, 369–373. [Google Scholar] [CrossRef]
- Holland, J.M.; Luff, M.L. The effects of agricultural practices on Carabidae in temperate agroecosystems. Integr. Pest Manage. Rev. 2000, 5, 109–129. [Google Scholar] [CrossRef]
- Fox, C.J.S. The effects of five herbicides on the numbers of certain invertebrate animals in grassland soil. Can. J. Plant Sci. 1964, 44, 405–409. [Google Scholar] [CrossRef] [Green Version]
- House, G.J. Soil arthropods from weed and crop roots of an agroecosystem in a wheat-soybean-corn rotation: Impact of tillage and herbicides. Agric. Ecosyst. Environ. 1989, 25, 233–244. [Google Scholar] [CrossRef]
- Badji, C.A.; Guedes, R.N.C.; Silva, A.A.; Correa, A.S.; Queiroz, M.E.L.R.; Michereff Filho, M. Non-target impact of deltamethrin on soil arthropods of maize fields under conventional and no-tillage cultivation. J. Appl. Entomol. 2007, 131, 50–58. [Google Scholar] [CrossRef] [Green Version]
- Ingram, J.W.; Bynum, E.K.; Charpentier, L.J. Effect of 2,4-D on sugarcane borer. J. Econ. Entomol. 1947, 40, 745–746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isenhour, D.J.; Todd, J.W.; Hauser, E.W. The impact of toxaphene applied as a post-emergence herbicide for control of sicklepod, Cassia obtusifolia L., on arthropods associated with soybean. Crop Prot. 1985, 4, 434–445. [Google Scholar] [CrossRef]
- Goldman, S.E.; Cleveland, G.D.; Parker, J.A. Notes: Lightwood induction and associated beetle attacks on Slash pine. Forest Sci. 1979, 25, 80–83. [Google Scholar] [CrossRef]
- Fox, W.B. 2,4-D as a factor increasing wireworm damage of wheat. Sci. Agric. 1948, 28, 423–424. [Google Scholar]
- Oka, I.N.; Pimentel, D. Herbicide (2,4-D) increases insect and pathogen pests on corn. Science 1976, 193, 239–240. [Google Scholar] [CrossRef]
- Shelton, M.D.; Edwards, C.R. Effects of weeds on the diversity and abundance of insects in soybeans. Environ. Entomol. 1983, 12, 296–298. [Google Scholar] [CrossRef] [Green Version]
- Powell, W.; Dean, G.J.; Dewar, A. The influence of weeds on polyphagous arthropod predators in winter wheat. Crop Prot. 1985, 4, 298–312. [Google Scholar] [CrossRef]
- Thiele, H.U. Carabid Beetles in Their Environments; Springer-Verlag: Berlin, Germany, 1977. [Google Scholar]
- Aebischer, N.J. Twenty years of monitoring invertebrates and weeds in cereal fields in Sussex. In The Ecology of Temperate Cereal Fields; Firbank, L.G., Carter, N., Darbyshire, J.F., Potts, G.R., Eds.; Blackwell: Oxford, UK, 1991; pp. 305–331. [Google Scholar]
- Jepson, P.C. The temporal and spatial dynamics of pesticide side-effects on non-target invertebrates. In Pesticides and Non-target Invertebrates; Jepson, P.C., Ed.; Intercept: Wimborne, UK, 1989; pp. 95–128. [Google Scholar]
- Gutiérrez, Y.; Ott, D.; Scherber, C. Direct and indirect effects of plant diversity and phenoxy herbicide application on the development and reproduction of a polyphagous herbivore. Sci. Rep. 2020, 10, 7300. [Google Scholar] [CrossRef]
- Muratet, A.; Fontaine, B. Contrasting impacts of pesticides on butterflies and bumblebees in private gardens in France. Biol. Conserv. 2015, 182, 148–154. [Google Scholar] [CrossRef]
- Rands, M.R.W.; Sotherton, N.W. Pesticide use on cereal crops and changes in the abundance of butterflies on arable farmland in England. Biol. Conserv. 1986, 36, 71–82. [Google Scholar] [CrossRef]
- Pleasants, J.M.; Oberhauser, K.S. Milkweed loss in agricultural fields because of herbicide use: Effect on the monarch butterfly population. Ins. Conserv. Divers. 2013, 6, 135–144. [Google Scholar] [CrossRef]
- Bohnenblust, E.; Egan, J.F.; Mortensen, D.; Tooker, J. Direct and indirect effects of the synthetic-auxin herbicide dicamba on two lepidopteran species. Environ. Entomol. 2013, 42, 586–894. [Google Scholar] [CrossRef]
- Russo, L.; Buckley, Y.M.; Hamilton, H.; Kavanagh, M.; Stout, J.C. Low concentrations of fertilizer and herbicide alter plant growth and interactions with flower-visiting insects. Agric. Ecosyst. Environ. 2020, 304, 107141. [Google Scholar] [CrossRef]
- Hardin, M.R.; Benrey, B.; Coll, M.; Lamp, W.O.; Roderick, G.K.; Barbosa, P. Arthropod pest resurgence: An overview of potential mechanisms. Crop Prot. 1995, 14, 3–18. [Google Scholar] [CrossRef]
- DeBach, P. Biological Control By Natural Enemies; Cambridge University Press: London, UK, 1974. [Google Scholar]
- Perfecto, I. Indirect and direct effects in a tropical agroecosystem: The maize-pest-ant system in Nicaragua. Ecology 1990, 71, 2125–2134. [Google Scholar] [CrossRef] [Green Version]
- Frampton, G.K.; Brink, P.J.v.d. Collembola and macroarthropod community responses to carbamate, organophosphate and synthetic pyrethroid insecticides: Direct and indirect effects. Environ. Pollut. 2007, 147, 14–25. [Google Scholar] [CrossRef]
- Eveleens, K.G.; van den Bosch, R.; Ehler, L.E. Secondary outbreak induction of beet armyworm by experimental insecticide applications in cotton in California. Environ. Entomol. 1973, 2, 497–504. [Google Scholar] [CrossRef]
- Castle, S.J.; Henneberry, T.J.; Toscano, N.C. Suppression of Bemisia tabaci (Homoptera: Aleyrodidae) infestations in cantaloupe and cotton with sprinkler irrigation. Crop Prot. 1996, 15, 657–663. [Google Scholar] [CrossRef]
- Cordeiro, E.M.G.; Corrêa, A.S.; Guedes, R.N.C. Insecticide-mediated shift in ecological dominance between two competing species of grain beetles. PLoS ONE 2014, 9, e100990. [Google Scholar] [CrossRef] [PubMed]
- Prischmann, D.A.; James, D.G.; Wright, L.C.; Teneyck, R.D.; Snyder, W.E. Effects of chlorpyrifos and sulfur on spider mites (Acari: Tetranychidae) and their natural enemies. Biol. Control 2005, 33, 324–334. [Google Scholar] [CrossRef]
- Lu, Y.; Wu, K.; Jiang, Y.; Guo, Y.; Desneux, N. Widespread adoption of Bt cotton and insecticide decrease promotes biocontrol services. Nature 2012, 487, 362–365. [Google Scholar] [CrossRef] [PubMed]
- Zeilinger, A.R.; Olson, D.M.; Andow, D.A. Competitive release and outbreaks of non-target pests associated with transgenic Bt cotton. Ecol. Appl. 2016, 26, 1047–1054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, L.-Z.; Bai, S.-X.; Zhao, J.-Z.; Wang, Z.-Y.; Wu, K.-M. Progress in ecological biosafety of insect-resistant transgenic cotton and corn in relation to arthropods. Acta Entomol. Sinica 2007, 50, 727–736. [Google Scholar]
- Heong, K.L.; Escalada, M.M.; Mai, V. An analysis of insecticide use in rice: Case studies in the Philippines and Vietnam. Int. J. Pest Control 1994, 40, 173–178. [Google Scholar] [CrossRef]
- Gross, K.; Rosenheim, J.A. Quantifying secondary pest outbreaks in cotton and their monetary cost with causal-inference statistics. Ecol. Appl. 2011, 21, 2770–2780. [Google Scholar] [CrossRef]
- Resende-Silva, G.A.; Turchen, L.M.; Guedes, R.N.C.; Cutler, G.C. Imidacloprid soil drenches affect weight and functional response of spined soldier bug (Hemiptera: Pentatomidae). J. Econ. Entomol. 2019, 112, 558–564. [Google Scholar] [CrossRef]
- Qi, B.; Gordon, G.; Gimme, W. Effects of neem-fed prey on the predacious insects Harmonia conformis (Boisduval) (Coleoptera: Coccinellidae) and Mallada signatus (Schneider) (Neuroptera: Chrysopidae). Biol. Control 2001, 22, 185–190. [Google Scholar] [CrossRef]
- Walker, M.K.; Stufkens, M.A.W.; Wallace, A.R. Indirect non-target effects of insecticides on Tasmanian brown lacewing (Micromus tasmaniae) from feeding on lettuce aphid (Nasonovia ribisnigri). Biol. Control 2007, 43, 31–40. [Google Scholar] [CrossRef]
- Scarpellini, J.R.; Andrade, D.J.d. The effect of insecticides on the lady beetle Cycloneda sanguinea L. (Coleoptera, Coccinellidae) and on the aphid Aphis gossypii Glover (Hemiptera, Aphididae) on cotton plants = Efeito de inseticidas sobre a joaninha Cycloneda sanguinea L. (Coleoptera, Coccinellidae) e sobre o pulgao Aphis gossypii Glover (Hemiptera, Aphididae) em algodoeiro. Arq. Inst. Biol. 2011, 78, 393–399. [Google Scholar]
- Mauchline, A.L.; Osborne, J.L.; Powell, W. Feeding responses of carabid beetles to dimethoate-contaminated prey. Agric. Forest Entomol. 2004, 6, 99–104. [Google Scholar] [CrossRef]
- Penca, C.; Hodges, A.C. Pyriproxyfen treatment terminates Halyomorpha halys reproductive diapause, with an indirect mortality effect on its egg parasitoid Trissolcus japonicus. J. Pest Sci. 2017, 90, 1269–1276. [Google Scholar] [CrossRef]
- Lira, A.C.S.; Wanderley-Teixeira, V.; Teixeira, Á.A.C.; Cunha, F.M.; Cruz, G.S.; Neto, C.J.C.L. Physiological and behavioral interactions of a predator with its prey under indirect exposure to the insect growth regulator lufenuron. Crop Prot. 2020, 137, 105289. [Google Scholar] [CrossRef]
- Choudhury, R.A.; Sutherland, A.M.; Hengel, M.J.; Parrella, M.P.; Gubler, W.D. Imidacloprid movement into fungal conidia is lethal to mycophagous beetles. Insects 2020, 11, 496. [Google Scholar] [CrossRef]
- Gontijo, P.C.; Moscardini, V.F.; Michaud, J.P.; Carvalho, G.A. Non-target effects of two sunflower seed treatments on Orius insidiosus (Hemiptera: Anthocoridae). Pest Manag. Sci. 2015, 71, 515–522. [Google Scholar] [CrossRef]
- Gontijo, P.C.; Moscardini, V.F.; Michaud, J.P.; Carvalho, G.A. Non-target effects of chlorantraniliprole and thiamethoxam on Chrysoperla carnea when employed as sunflower seed treatments. J. Pest Sci. 2014, 87, 711–719. [Google Scholar] [CrossRef] [Green Version]
- Goulson, D.; Nicholls, E.; Botiías, C.; Rotheray, E.L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 2015, 347, 1255957. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Bayo, F.; Goulson, D.; Pennacchio, F.; Nazzi, F.; Goka, K.; Desneux, N. Are bee diseases linked to pesticides? — A brief review. Environ. Int. 2016, 89–90, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Brandt, A.; Gorenflo, A.; Siede, R.; Meixner, M.; Buüchler, R. The neonicotinoids thiacloprid, imidacloprid, and clothianidin affect the immunocompetence of honey bees (Apis mellifera L.). J. Ins. Physiol. 2016, 86, 40–47. [Google Scholar] [CrossRef]
- Long, E.Y.; Krupke, C.H. Non-cultivated plants present a season-long route of pesticide exposure for honey bees. Nat Commun 2016, 7, 11629. [Google Scholar] [CrossRef]
- Botías, C.; David, A.; Hill, E.M.; Goulson, D. Contamination of wild plants near neonicotinoid seed-treated crops, and implications for non-target insects. Sci. Total Environ. 2016, 566–567, 269–278. [Google Scholar] [CrossRef]
- Tong, L.; Nieh, J.C.; Tosi, S. Combined nutritional stress and a new systemic pesticide (flupyradifurone, Sivanto®) reduce bee survival, food consumption, flight success, and thermoregulation. Chemosphere 2019, 237, 124408. [Google Scholar] [CrossRef] [Green Version]
- Brandt, A.; Hohnheiser, B.; Sgolastra, F.; Bosch, J.; Meixner, M.D.; Büchler, R. Immunosuppression response to the neonicotinoid insecticide thiacloprid in females and males of the red mason bee Osmia bicornis L. Sci. Rep. 2020, 10, 4670. [Google Scholar] [CrossRef] [PubMed]
- Botías, C.; Jones, J.C.; Pamminger, T.; Bartomeus, I.; Hughes, W.O.H.; Goulson, D. Multiple stressors interact to impair the performance of bumblebee Bombus terrestris colonies. J. Anim. Ecol. 2021, 90. [Google Scholar] [CrossRef] [PubMed]
- Pettis, J.; van Engelsdorp, D.; Johnson, J.; Dively, G. Pesticide exposure in honey bees results in increased levels of the gut pathogen Nosema. Naturwissenschaften [Sci. Nat.] 2012, 99, 153–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higes, M.; Martín-Hernández, R.; Botías, C.; Bailón, E.G.; González-Porto, A.V.; Barrios, L.; del Nozal, M.J.; Bernal, J.L.; Jiménez, J.J.; Palencia, P.G.; et al. How natural infection by Nosema ceranae causes honeybee colony collapse. Environ. Microbiol. 2008, 10, 2659–2669. [Google Scholar] [CrossRef]
- Rothman, J.A.; Russell, K.A.; Leger, L.; McFrederick, Q.S.; Graystock, P. The direct and indirect effects of environmental toxicants on the health of bumblebees and their microbiomes: Impact of toxicants on bumblebee health. Proc. R. Soc. B 2020, 287, 20200980. [Google Scholar] [CrossRef] [PubMed]
- Alburaki, M.; Boutin, S.; Mercier, P.-L.; Loublier, Y.; Chagnon, M.; Derome, N. Neonicotinoid-coated Zea mays seeds indirectly affect honeybee performance and pathogen susceptibility in field trials. PLoS ONE 2015, 10, e0125790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandler, A.J.; Drummond, F.A.; Drummond, F.A.; Collins, J.A.; Lund, J.; Alnajjar, G. Exposure of the common eastern bumble bee, Bombus impatiens (Cresson), to sub-lethal doses of acetamiprid and propiconazole in wild blueberry. J. Agric. Urban Entomol. 2020, 36, 1–23. [Google Scholar] [CrossRef]
- Evans, A.N.; Llanos, J.E.M.; Kunin, W.E.; Evison, S.E.F. Indirect effects of agricultural pesticide use on parasite prevalence in wild pollinators. Agric. Ecosyst. Environ. 2018, 258, 40–48. [Google Scholar] [CrossRef] [Green Version]
- Anderson, J.R. Recent developments in the control of some arthropods of public health and veterinary importance: Muscoid flies. Bull. Entomol. Soc. Am. 1966, 12, 342–348. [Google Scholar] [CrossRef]
- Floate, K.D.; Wardhaugh, K.G.; Boxall, A.B.A.; Sherratt, T.N. Fecal residues of veterinary parasiticides: Non-target effects in the pasture environment. Annu. Rev. Entomol. 2005, 50, 153–180. [Google Scholar] [CrossRef]
- McKellar, Q.A. Ecotoxicology and residues of anthelmintic compounds. Veterinary Parasitology 1997, 72, 413–435. [Google Scholar] [CrossRef]
- Wardhaugh, K.G. Insecticidal activity of synthetic pyrethroids, organophosphates, insect growth regulators, and other livestock parasiticides: An Australian perspective. Environ. Toxicol. Chem. 2005, 24, 789–796. [Google Scholar] [CrossRef] [PubMed]
- Cook, C.W.; Gerhardt, R.R. Selective mortality of insects in manure from cattle fed Rabon® and Dimilin®. Environ. Entomol. 1977, 6, 589–590. [Google Scholar] [CrossRef]
- Floate, K.D. Off-target effects of ivermectin on insects and on dung degradation in southern Alberta, Canada. Bull. Entomol. Res. 1998, 88, 25–35. [Google Scholar] [CrossRef]
- Lumaret, J.P.; Galante, E.; Lumbreras, C.; Mena, J.; Bertrand, M.; Bernal, J.L.; Cooper, J.F.; Kadiri, N.; Crowe, D. Field effects of ivermectin residues on dung beetles. J. Appl. Ecol. 1993, 30, 428–436. [Google Scholar] [CrossRef]
- Krüger, K.; Scholtz, C.H. Changes in the structure of dung insect communities after ivermectin usage in a grassland ecosystem. I. Impact of ivermectin under drought conditions. Acta Oecol. 1998, 19, 425–438. [Google Scholar] [CrossRef]
- Krüger, K.; Scholtz, C.H. The effect of ivermectin on the development and reproduction of the dung-breeding fly Musca nevilli Kleynhans (Diptera, Muscidae). Agric. Ecosyst. Environ. 1995, 53, 13–18. [Google Scholar] [CrossRef]
- Sommer, C.; Jensen, K.M.V.; Jespersen, J.B. Topical treatment of calves with synthetic pyrethroids: Effects on the non-target dung fly Neomyia cornicina (Diptera: Muscidae). Bull. Entomol. Res. 2001, 91, 131–137. [Google Scholar] [CrossRef]
- Bianchin, I.; Alves, R.G.O.; Koller, W.W. Effect of pour-on tickicides/insecticides on adults of the African dung-beetle Onthophagus gazella Fabr. (Coleoptera: Scarabaeidae). An. Soc. Entomol. Bras. 1998, 27, 275–279. [Google Scholar] [CrossRef]
- Wardhaugh, K.G.; Longstaff, B.C.; Lacey, M.J. Effects of residues of deltamethrin in cattle faeces on the development and survival of three species of dung-breeding insects. Aust. Vet. J. 1998, 76, 273–280. [Google Scholar] [CrossRef]
- Suarez, V.H.; Lifschitz, A.L.; Sallovitz, J.M.; Lanusse, C.E. Effects of ivermectin and doramectin faecal residues on the invertebrate colonization of cattle dung. J. Appl. Entomol. 2003, 127, 481–488. [Google Scholar] [CrossRef]
- Dadour, I.R.; Cook, D.F.; Hennessy, D. Reproduction and survival of the dung beetle Onthophagus binodis (Coleoptera: Scarabaeidae) exposed to abamectin and doramectin residues in cattle dung. Environ. Entomol. 2000, 29, 1116–1122. [Google Scholar] [CrossRef]
- Wardhaugh, K.G.; Holter, P.; Longstaff, B. The development and survival of three species of coprophagous insect after feeding on the faeces of sheep treated with controlled-release formulations of ivermectin or albendazole. Aust. Vet. J. 2001, 79, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Vale, G.A.; Mutika, G.; Lovemore, D.F. Insecticide-treated cattle for controlling tsetse flies (Diptera: Glossinidae): Some questions answered, many posed. Bull. Entomol. Res. 1999, 89, 569–578. [Google Scholar] [CrossRef] [Green Version]
- Herrick, N.J.; Cloyd, R.A. Direct and indirect effects of pesticides on the insidious flower bug (Hemiptera: Anthocoridae) under laboratory conditions. J. Econ. Entomol. 2017, 110, 931–940. [Google Scholar] [CrossRef]
- Pozzebon, A.; Borgo, M.; Duso, C. The effects of fungicides on non-target mites can be mediated by plant pathogens. Chemosphere 2010, 79, 8–17. [Google Scholar] [CrossRef]
- Smith, R.G.; Atwood, L.W.; Morris, M.B.; Mortensen, D.A.; Koide, R.T. Evidence for indirect effects of pesticide seed treatments on weed seed banks in maize and soybean. Agric. Ecosyst. Environ. 2016, 216, 269–273. [Google Scholar] [CrossRef]
- Frampton, G.K.; Dorne, J.L.C.M. The effects on terrestrial invertebrates of reducing pesticide inputs in arable crop edges: A meta-analysis. J. Appl. Ecol. 2007, 44, 362–373. [Google Scholar] [CrossRef]
- Teklu, B.M.; Hailu, A.; Wiegant, D.A.; Scholten, B.S.; van den Brink, P.J. Impacts of nutrients and pesticides from small- and large-scale agriculture on the water quality of Lake Ziway, Ethiopia. Environ. Sci. Pollut. Res. 2018, 25, 13207–13216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basley, K.; Goulson, D. Neonicotinoids thiamethoxam and clothianidin adversely affect the colonisation of invertebrate populations in aquatic microcosms. Environ. Sci. Pollut. Res. 2018, 25, 9593–9599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Standley, L.J.; Sweeney, B.W. Organochlorine pesticides in stream mayflies and terrestrial vegetation of undisturbed tropical catchments exposed to long-range atmospheric transport. J. North Am. Benthol. Soc. 1995, 14, 38–49. [Google Scholar] [CrossRef]
- Niemi, G.J.; Hershey, A.E.; Shannon, L.; Hanowski, J.M.; Lima, A.; Axler, R.P.; Regal, R.R. Ecological effects of mosquito control on zooplankton, insects and birds. Environ. Toxicol. Chem. 1999, 18, 549–559. [Google Scholar] [CrossRef]
- Sánchez-Bayo, F. Comparative acute toxicity of organic pollutants and reference values for crustaceans. I. Branchiopoda, Copepoda and Ostracoda. Environ. Pollut. 2006, 139, 385–420. [Google Scholar] [CrossRef]
- Anderson, R.L. Toxicity of synthetic pyrethroids to freshwater invertebrates. Environ. Toxicol. Chem. 1989, 8, 403–410. [Google Scholar] [CrossRef]
- Liess, M.; Liebmann, L.; Vormeier, P.; Weisner, O.; Altenburger, R.; Borchardt, D.; Brack, W.; Chatzinotas, A.; Escher, B.; Foit, K.; et al. Pesticides are the dominant stressors for vulnerable insects in lowland streams. Water Res. 2021, 201, 117262. [Google Scholar] [CrossRef] [PubMed]
- Hanazato, T. Response of a zooplankton community to insecticide application in experimental ponds: A review and the implications of the effects of chemicals on the structure and functioning of freshwater communities. Environ. Pollut. 1998, 101, 361–373. [Google Scholar] [CrossRef]
- Farmer, D.; Hill, I.R.; Maund, S.J. A comparison of the fate and effects of two pyrethroid insecticides (lambda-cyhalothrin and cypermethrin) in pond mesocosms. Ecotoxicology 1995, 4, 219–244. [Google Scholar] [CrossRef]
- Relyea, R.A. The impact of insecticides and herbicides on the biodiversity and productivity of aquatic communities. Ecol. Appl. 2005, 15, 618–627. [Google Scholar] [CrossRef]
- Sumon, K.A.; Ritika, A.K.; Peeters, E.T.H.M.; Rashid, H.; Bosma, R.H.; Rahman, M.S.; Fatema, M.K.; van den Brink, P.J. Effects of imidacloprid on the ecology of sub-tropical freshwater microcosms. Environ. Pollut. 2018, 236, 432–441. [Google Scholar] [CrossRef] [PubMed]
- Kreutzweiser, D.P.; Capell, S.S.; Thomas, D.R. Aquatic insect responses to Bacillus thuringiensis var. kurstaki in a forest stream. Can. J. Forest Res. 1994, 24, 2041–2049. [Google Scholar] [CrossRef]
- Ward, S.; Arthington, A.H.; Pusey, B.J. The effects of a chronic application of chlorpyrifos on the macroinvertebrate fauna in an outdoor artificial stream system: Species responses. Ecotoxicol. Environ. Saf. 1995, 30, 2–23. [Google Scholar] [CrossRef] [PubMed]
- Schroll, H.; Pedersen, C.L.; Jespersen, P.H. Indirect effects of esfenvalerate (Insecticide) on the density of periphytic algae in artificial ponds. Bull. Environ. Contam. Toxicol. 1998, 60, 797–801. [Google Scholar] [CrossRef]
- Rodrigues, A.C.M.; Machado, A.L.; Bordalo, M.D.; Saro, L.; Simão, F.C.P.; Rocha, R.J.M.; Golovko, O.; Žlábek, V.; Barata, C.; Soares, A.M.V.M.; et al. Invasive species mediate insecticide effects on community and ecosystem functioning. Environ. Sci. Technol. 2018, 52, 4889–4900. [Google Scholar] [CrossRef] [PubMed]
- Hasenbein, S.; Lawler, S.P.; Geist, J.; Connon, R.E. A long-term assessment of pesticide mixture effects on aquatic invertebrate communities. Environ. Toxicol. Chem. 2016, 35, 218–232. [Google Scholar] [CrossRef]
- Woin, P. Short- and long-term effects of the pyrethroid insecticide fenvalerate on an invertebrate pond community. Ecotoxicol. Environ. Saf. 1998, 41, 137–156. [Google Scholar] [CrossRef]
- Jones, D.K.; Hua, J.; Relyea, R.A. Effects of endosulfan in freshwater pond communities. Freshwat. Sci. 2016, 35, 152–163. [Google Scholar] [CrossRef]
- Hua, J.; Relyea, R. Chemical cocktails in aquatic systems: Pesticide effects on the response and recovery of >20 animal taxa. Environ. Pollut. 2014, 189, 18–26. [Google Scholar] [CrossRef]
- Hayasaka, D.; Korenaga, T.; Sánchez-Bayo, F.; Goka, K. Differences in ecological impacts of systemic insecticides with different physicochemical properties on biocenosis of experimental paddy fields. Ecotoxicology 2012, 21, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Groner, M.L.; Relyea, R.A. A tale of two pesticides: How common insecticides affect aquatic communities. Freshwat. Biol. 2011, 56, 2391–2404. [Google Scholar] [CrossRef]
- Vilas-Boas, J.A.; Arenas-Sánchez, A.; Vighi, M.; Romo, S.; van den Brink, P.J.; Pedroso Dias, R.J.; Rico, A. Multiple stressors in Mediterranean coastal wetland ecosystems: Influence of salinity and an insecticide on zooplankton communities under different temperature conditions. Chemosphere 2021, 269, 129381. [Google Scholar] [CrossRef]
- Brogan, W.R., III; Relyea, R.A. Submerged macrophytes mitigate direct and indirect insecticide effects in freshwater communities. PLoS ONE 2015, 10, e0126677. [Google Scholar] [CrossRef]
- Graf, N.; Battes, K.P.; Cimpean, M.; Dittrich, P.; Entling, M.H.; Link, M.; Scharmüller, A.; Schreiner, V.C.; Szöcs, E.; Schäfer, R.B. Do agricultural pesticides in streams influence riparian spiders? Sci. Total Environ. 2019, 660, 126–135. [Google Scholar] [CrossRef]
- López-Mancisidor, P.; Carbonell, G.; Fernández, C.; Tarazona, J.V. Ecological impact of repeated applications of chlorpyrifos on zooplankton community in mesocosms under Mediterranean conditions. Ecotoxicology 2008, 17, 811–825. [Google Scholar] [CrossRef] [PubMed]
- Daam, M.A.; van den Brink, P.J.; Nogueira, A.J.A. Impact of single and repeated applications of the insecticide chlorpyrifos on tropical freshwater plankton communities. Ecotoxicology 2008, 17, 756–771. [Google Scholar] [CrossRef] [PubMed]
- Yamamuro, M.; Komuro, T.; Kamiya, H.; Kato, T.; Hasegawa, H.; Kameda, Y. Neonicotinoids disrupt aquatic food webs and decrease fishery yields. Science 2019, 366, 620–623. [Google Scholar] [CrossRef] [PubMed]
- Kobashi, K.; Harada, T.; Adachi, Y.; Mori, M.; Ihara, M.; Hayasaka, D. Comparative ecotoxicity of imidacloprid and dinotefuran to aquatic insects in rice mesocosms. Ecotoxicol. Environ. Saf. 2017, 138, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, K.; Kasai, A.; Hayasaka, D.; Goka, K.; Hayashi, T.I. Long-term monitoring reveals among-year consistency in the ecological impacts of insecticides on animal communities in paddies. Ecol. Ind. 2020, 113, 106227. [Google Scholar] [CrossRef]
- Liess, M. Population response to toxicants is altered by intraspecific interaction. Environ. Toxicol. Chem. 2002, 21, 138–142. [Google Scholar] [CrossRef] [PubMed]
- Liess, M.; Foit, K. Intraspecific competition delays recovery of population structure. Aquat. Toxicol. 2010, 97, 15–22. [Google Scholar] [CrossRef]
- Kreutzweiser, D.P.; Good, K.P.; Chartrand, D.T.; Scarr, T.A.; Thompson, D.G. Are leaves that fall from imidacloprid-treated maple trees to control Asian longhorned beetles toxic to non-target decomposer organisms? J. Environ. Qual. 2008, 37, 639–646. [Google Scholar] [CrossRef]
- Nyman, A.-M.; Hintermeister, A.; Schirmer, K.; Ashauer, R. The insecticide imidacloprid causes mortality of the freshwater amphipod Gammarus pulex by interfering with feeding behavior. PLoS ONE 2013, 8, e62472. [Google Scholar] [CrossRef] [Green Version]
- Englert, D.; Bundschuh, M.; Schulz, R. Thiacloprid affects trophic interaction between gammarids and mayflies. Environ. Pollut. 2012, 167, 41–46. [Google Scholar] [CrossRef]
- Englert, D.; Zubrod, J.P.; Link, M.; Mertins, S.; Schulz, R.; Bundschuh, M. Does waterborne exposure explain effects caused by neonicotinoid-contaminated plant material in aquatic systems? Environ. Sci. Technol. 2017, 51, 5793–5802. [Google Scholar] [CrossRef] [PubMed]
- Pestana, J.L.T.; Alexander, A.C.; Culp, J.M.; Baird, D.J.; Cessna, A.J.; Soares, A.M.V.M. Structural and functional responses of benthic invertebrates to imidacloprid in outdoor stream mesocosms. Environ. Pollut. 2009, 157, 2328–2334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pestana, J.L.T.; Loureiro, S.; Baird, D.J.; Soares, A.M.V.M. Fear and loathing in the benthos: Responses of aquatic insect larvae to the pesticide imidacloprid in the presence of chemical signals of predation risk. Aquat. Toxicol. 2009, 93, 138–149. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, A.C.M.; Bordalo, M.D.; Golovko, O.; Koba, O.; Barata, C.; Soares, A.M.V.M.; Pestana, J.L.T. Combined effects of insecticide exposure and predation risk on freshwater detritivores. Ecotoxicology 2018, 27, 794–802. [Google Scholar] [CrossRef]
- Pestana, J.L.T.; Loureiro, S.; Baird, D.J.; Soares, A.M.V.M. Pesticide exposure and inducible antipredator responses in the zooplankton grazer, Daphnia magna Straus. Chemosphere 2010, 78, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Zubrod, J.P.; Bundschuh, M.; Arts, G.; Brühl, C.A.; Imfeld, G.; Knäbel, A.; Payraudeau, S.; Rasmussen, J.J.; Rohr, J.; Scharmüller, A.; et al. Fungicides: An overlooked pesticide class? Environ. Sci. Technol. 2019, 53, 3347–3365. [Google Scholar] [CrossRef]
- Zubrod, J.P.; Englert, D.; Wolfram, J.; Rosenfeldt, R.R.; Feckler, A.; Bundschuh, R.; Seitz, F.; Konschak, M.; Baudy, P.; Lüderwald, S.; et al. Long-term effects of fungicides on leaf-associated microorganisms and shredder populations—An artificial stream study. Environ. Toxicol. Chem. 2017, 36, 2178–2189. [Google Scholar] [CrossRef]
- Willis, K.J.; van den Brink, P.J.; Green, J.D. Seasonal variation in plankton community responses of mesocosms dosed with pentachlorophenol. Ecotoxicology 2004, 13, 707–720. [Google Scholar] [CrossRef] [PubMed]
- Daam, M.A.; Satapornvanit, K.; van den Brink, P.J.; Nogueira, A.J.A. Direct and indirect effects of the fungicide carbendazim in tropical freshwater microcosms. Arch. Environ. Contam. Toxicol. 2010, 58, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Lin, R.; Buijse, L.; Dimitrov, M.; Dohmen, P.; Kosol, S.; Maltby, L.; Roessink, I.; Sinkeldam, J.; Smidt, H.; Van Wijngaarden, R.; et al. Effects of the fungicide metiram in outdoor freshwater microcosms: Responses of invertebrates, primary producers and microbes. Ecotoxicology 2012, 21, 1550–1569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuppen, J.G.M.; van den Brink, P.J.; Camps, E.; Uil, K.F.; Brock, T.C.M. Impact of the fungicide carbendazim in freshwater microcosms. I. Water quality, breakdown of particulate organic matter and responses of macroinvertebrates. Aquat. Toxicol. 2000, 48, 233–250. [Google Scholar] [CrossRef]
- Jacobus, L.M.; Macadam, C.R.; Sartori, M. Mayflies (Ephemeroptera) and their contributions to ecosystem services. Insects 2019, 10, 170. [Google Scholar] [CrossRef] [Green Version]
- Morse, J.C.; Frandsen, P.B.; Graf, W.; Thomas, J.A. Diversity and ecosystem services of Trichoptera. Insects 2019, 10, 125. [Google Scholar] [CrossRef] [Green Version]
- Bayona, Y.; Roucaute, M.; Cailleaud, K.; Lagadic, L.; Bassères, A.; Caquet, T. Effect of thiram and of a hydrocarbon mixture on freshwater macroinvertebrate communities in outdoor stream and pond mesocosms: I. Study design, chemicals fate and structural responses. Ecotoxicology 2015, 24, 1976–1995. [Google Scholar] [CrossRef]
- Bundschuh, M.; Zubrod, J.P.; Kosol, S.; Maltby, L.; Stang, C.; Duester, L.; Schulz, R. Fungal composition on leaves explains pollutant-mediated indirect effects on amphipod feeding. Aquat. Toxicol. 2011, 104, 32–37. [Google Scholar] [CrossRef]
- Rasmussen, J.J.; Monberg, R.J.; Baattrup-Pedersen, A.; Cedergreen, N.; Wiberg-Larsen, P.; Strobel, B.; Kronvang, B. Effects of a triazole fungicide and a pyrethroid insecticide on the decomposition of leaves in the presence or absence of macroinvertebrate shredders. Aquat. Toxicol. 2012, 118–119, 54–61. [Google Scholar] [CrossRef] [Green Version]
- Newton, K.; Zubrod, J.P.; Englert, D.; Lüderwald, S.; Schell, T.; Baudy, P.; Konschak, M.; Feckler, A.; Schulz, R.; Bundschuh, M. The evil within? Systemic fungicide application in trees enhances litter quality for an aquatic decomposer-detritivore system. Environ. Pollut. 2018, 241, 549–556. [Google Scholar] [CrossRef] [PubMed]
- Casado, J.; Brigden, K.; Santillo, D.; Johnston, P. Screening of pesticides and veterinary drugs in small streams in the European Union by liquid chromatography high resolution mass spectrometry. Sci. Total Environ. 2019, 670, 1204–1225. [Google Scholar] [CrossRef]
- Rybicki, M.; Jungmann, D. Direct and indirect effects of pesticides on a benthic grazer during its life cycle. Environ. Sci. Europe 2018, 30, 35. [Google Scholar] [CrossRef] [Green Version]
- Dewey, S.L. Effects of the herbicide atrazine on aquatic insect community structure and emergence. Ecology 1986, 67, 148–162. [Google Scholar] [CrossRef]
- Tsuruta, T.; Tada, T.; Kotera, N.; Akagawa, I.; Iguchi, K. Effects of predators and herbicides on community structure of benthic invertebrates in paddy fields around the Chikuma river. Jpn. J. Limnol. 2010, 70, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, K.; Eguchi, Y.; Oishi, H.; Tazunoki, Y.; Tokuda, M.; Sánchez-Bayo, F.; Goka, K.; Hayasaka, D. Effects of a herbicide on paddy predatory insects depend on their microhabitat use and an insecticide application. Ecol. Appl. 2019, 29, e01945. [Google Scholar] [CrossRef]
- Brock, T.C.M.; Bogaert, M.; Bos, A.R.; Breukelen, S.W.F.; Reiche, R.; Terwoert, J.; Suykerbuyk, R.E.M.; Roijackers, R.M.M. Fate and effects of the insecticide Dursban® 4E in indoor Elodea-dominated and macrophyte-free freshwater model ecosystems: II. Secondary effects on community structure. Arch. Environ. Contam. Toxicol. 1992, 23, 391–409. [Google Scholar] [CrossRef] [PubMed]
- Halstead, N.T.; McMahon, T.A.; Johnson, S.A.; Raffel, T.R.; Romansic, J.M.; Crumrine, P.W.; Rohr, J.R. Community ecology theory predicts the effects of agrochemical mixtures on aquatic biodiversity and ecosystem properties. Ecol. Lett. 2014, 17, 932–941. [Google Scholar] [CrossRef] [PubMed]
- Choung, C.B.; Hyne, R.V.; Stevens, M.M.; Hose, G.C. The ecological effects of a herbicide-insecticide mixture on an experimental freshwater ecosystem. Environ. Pollut. 2013, 172, 264–274. [Google Scholar] [CrossRef]
- Neury-Ormanni, J.; Doose, C.; Majdi, N.; Vedrenne, J.; Traunspurger, W.; Morin, S. Selective grazing behaviour of chironomids on microalgae under pesticide pressure. Sci. Total Environ. 2020, 730, 138673. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Bayo, F. Indirect Effect of Pesticides on Insects and Other Arthropods. Toxics 2021, 9, 177. https://doi.org/10.3390/toxics9080177
Sánchez-Bayo F. Indirect Effect of Pesticides on Insects and Other Arthropods. Toxics. 2021; 9(8):177. https://doi.org/10.3390/toxics9080177
Chicago/Turabian StyleSánchez-Bayo, Francisco. 2021. "Indirect Effect of Pesticides on Insects and Other Arthropods" Toxics 9, no. 8: 177. https://doi.org/10.3390/toxics9080177
APA StyleSánchez-Bayo, F. (2021). Indirect Effect of Pesticides on Insects and Other Arthropods. Toxics, 9(8), 177. https://doi.org/10.3390/toxics9080177