A Review of Bioinformatics Tools to Understand Acetaminophen-Alcohol Interaction


 , and
, and
Abstract
:1. Introduction
1.1. Drug-Related Hepatotoxicity
1.2. Acetaminophen and Ethanol Metabolism
2. Materials and Methods
2.1. Mechanism of Interaction between Acetaminophen and Ethanol
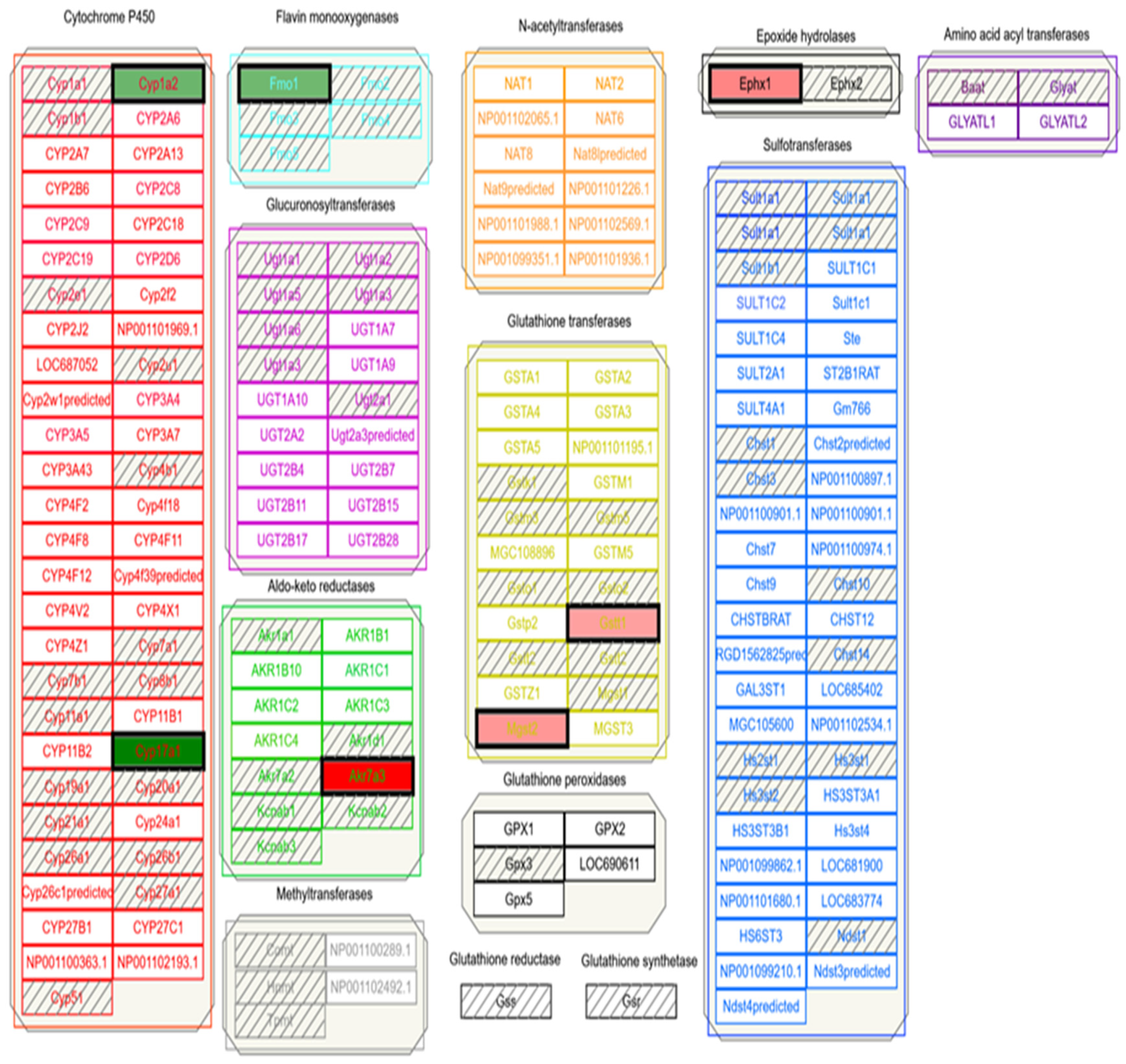
2.2. Tools to Understand Ethanol-Acetaminophen Interaction
2.3. Application of Bioinformatics Tools
2.4. Gene Expression Omnibus Data
3. Results
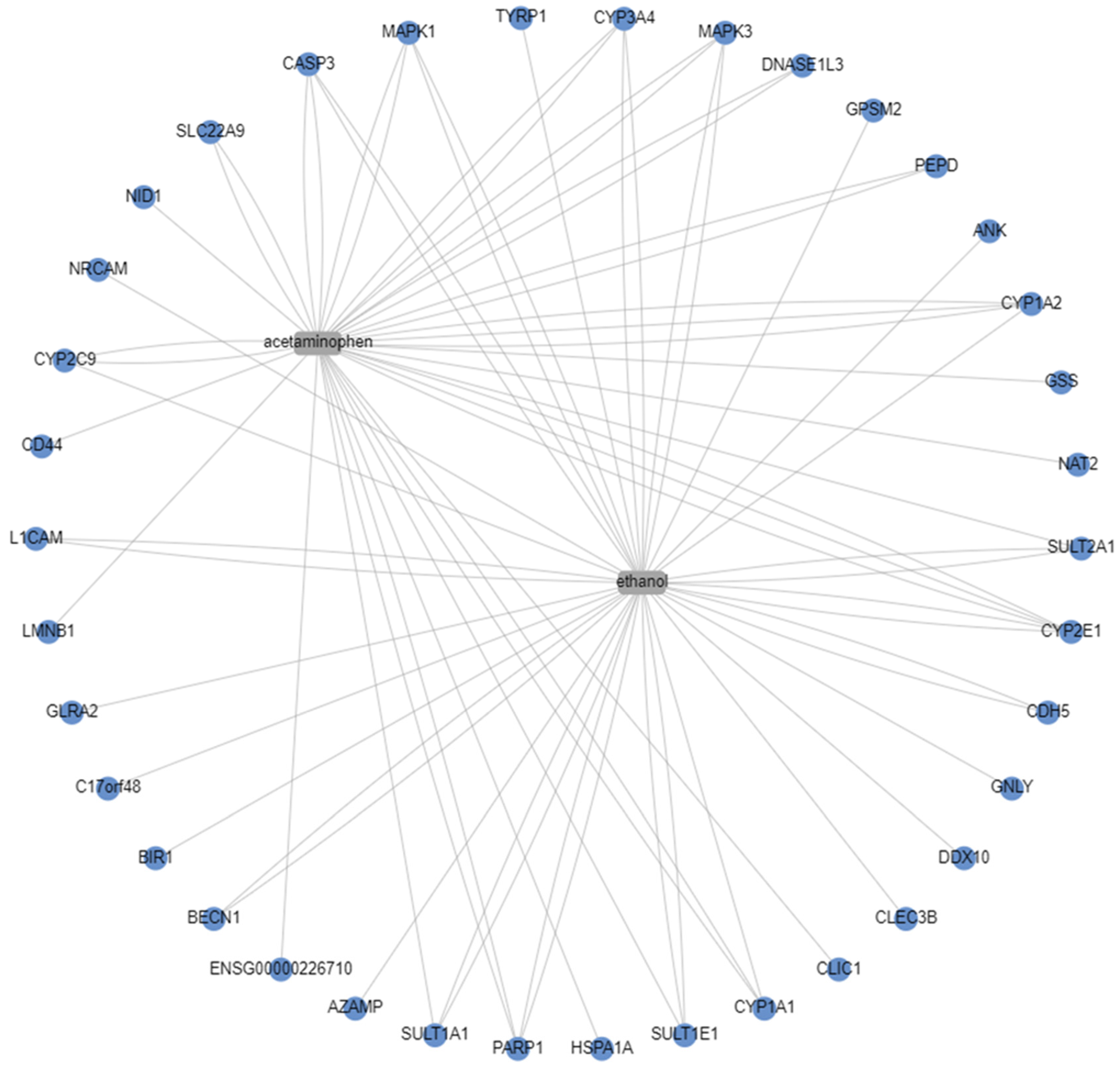
3.1. CART Galaxy Analysis
3.2. STRING Analysis
3.3. Transcriptome Analysis Console 4.0 Dataset Analysis
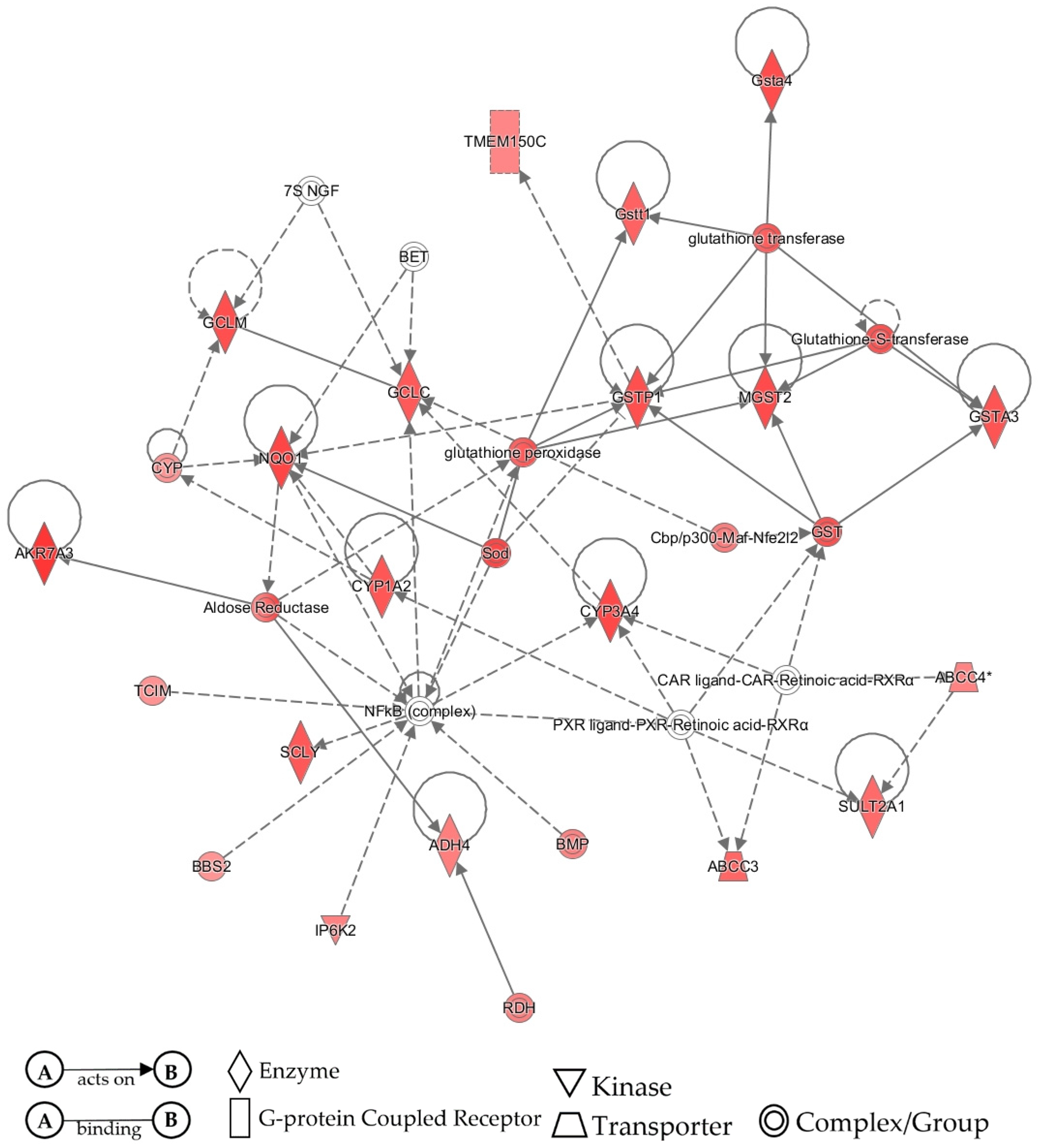
3.4. IPA Validation of Pathway Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Gurwitz, D.; Weizman, A.; Rehavi, M. Education: Teaching pharmacogenomics to prepare future physicians and researchers for personalized medicine. Trends Pharmacol. Sci. 2003, 24, 122–125. [Google Scholar] [CrossRef]
- Wang, X.; Liotta, L. Clinical bioinformatics: A new emerging science. J. Clin. Bioinform. 2011, 1, 1. [Google Scholar] [CrossRef] [PubMed]
- Yan, B.; Ban, K.H.K.; Tan, T.W. Integrating translational bioinformatics into the medical curriculum. Int. J. Med. Educ. 2014, 5, 132–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weathermon, R.; Crabb, D.W. Alcohol and medication interactions. Alcohol Res. Health 1999, 23, 40–54. [Google Scholar] [PubMed]
- Draganov, P.; Durrence, H.; Cox, C.; Reuben, A. Alcohol-acetaminophen syndrome. Postgrad. Med. 2000, 107, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Savides, M.C.; Oehme, F.W. Acetaminophen and its toxicity. J. Appl. Toxicol. 1983, 3, 96–111. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, K.K.; Jadeja, R.N.; Thadani, J.M.; Joshi, A.; Vohra, A.; Mevada, V.; Patel, R.; Khurana, S.; Devkar, R.V. Carbon monoxide releasing molecule A-1 attenuates acetaminophen-mediated hepatotoxicity and improves survival of mice by induction of Nrf2 and related genes. Toxicol. Appl. Pharmacol. 2018, 360, 99–108. [Google Scholar] [CrossRef]
- McClain, C.J.; Kromhout, J.P.; Peterson, F.J.; Holtzman, J.L. Potentiation of Acetaminophen Hepatotoxicity by Alcohol. JAMA 1980, 244, 251–253. [Google Scholar] [CrossRef]
- Adams, W.L. Interactions between alcohol and other drugs. Int. J. Addict. 1995, 30, 1903–1923. [Google Scholar] [CrossRef]
- Altomare, E.; Leo, M.A.; Lieber, C.S. Interaction of acute ethanol administration with acetaminophen metabolism and toxicity in rats fed alcohol chronically. Alcohol. Clin. Exp. Res. 1984, 8, 405–408. [Google Scholar] [CrossRef]
- Bromer, M.Q.; Black, M. Acetaminophen hepatotoxicity. Clin. Liver Dis. 2003, 7, 351–367. [Google Scholar] [CrossRef]
- Du, K.; Ramachandran, A.; Jaeschke, H. Oxidative stress during acetaminophen hepatotoxicity: Sources, pathophysiological role and therapeutic potential. Redox Biol. 2016, 10, 148–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, G.R.; Harris, R.Z. Drug interactions involving ethanol and alcoholic beverages. Expert Opin. Drug Metab. Toxicol. 2007, 3, 719–731. [Google Scholar] [CrossRef] [PubMed]
- Kuffner, E.; Green, J.; Bogdan, G.; Knox, P.; Palmer, R.; Heard, K.; Slattery, J.T.; Dart, R. The effect of acetaminophen (four grams a day for three consecutive days) on hepatic tests in alcoholic patients—A multicenter randomized study. BMC Med. 2007, 5, 13. [Google Scholar] [CrossRef] [PubMed]
- Mazaleuskaya, L.L.; Sangkuhl, K.; Thorn, C.F.; FitzGerald, G.A.; Altman, R.B.; Klein, T.E. PharmGKB summary: Pathways of acetaminophen metabolism at the therapeutic versus toxic doses. Pharm. Genom. 2015, 25, 416–426. [Google Scholar] [CrossRef] [PubMed]
- Prescott, L.F. Paracetamol, alcohol and the liver. Br. J. Clin. Pharmacol. 2000, 49, 291–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hodgman, M.J.; Garrard, A.R. A review of acetaminophen poisoning. Crit. Care Clin. 2012, 28, 499–516. [Google Scholar] [CrossRef] [PubMed]
- Bessems, J.G.; Vermeulen, N.P. Paracetamol (acetaminophen)-induced toxicity: Molecular and biochemical mechanisms, analogues and protective approaches. Crit. Rev. Toxicol. 2001, 31, 55–138. [Google Scholar] [CrossRef] [PubMed]
- Whirl-Carrillo, M.; McDonagh, E.M.; Hebert, J.M.; Gong, L.; Sangkuhl, K.; Thorn, C.F.; Altman, R.B.; Klein, T.E. Pharmacogenomics knowledge for personalized medicine. Clin. Pharmacol.Ther. 2012, 92, 414–417. [Google Scholar] [CrossRef] [PubMed]
- James, L.P.; Mayeux, P.R.; Hinson, J.A. Acetaminophen-induced hepatotoxicity. Drug Metab. Dispos. 2003, 31, 1499–1506. [Google Scholar] [CrossRef] [PubMed]
- Cederbaum, A.I. Alcohol metabolism. Clin. Liver Dis. 2012, 16, 667–685. [Google Scholar] [CrossRef]
- Bolt, H.M.; Roos, P.H.; Thier, R. The cytochrome P-450 isoenzyme CYP2E1 in the biological processing of industrial chemicals: Consequences for occupational and environmental medicine. Int. Arch. Occup. Environ. Health 2003, 76, 174–185. [Google Scholar] [CrossRef]
- Gonzalez, F.J. Role of cytochromes P450 in chemical toxicity and oxidative stress: Studies with CYP2E1. Mutat. Res. 2005, 569, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Lieber, C.S. Cytochrome P-4502E1: Its physiological and pathological role. Physiol. Rev. 1997, 77, 517–544. [Google Scholar] [CrossRef]
- Zakhari, S. Overview: How is Alcohol Metabolized by the Body? Division of Metabolism and Health Effects, National Institute on Alcohol Abuse and Alcoholism. Available online: https://pubs.niaaa.nih.gov/publications/arh294/245-255.htm (accessed on 24 July 2019).
- Caudle, K.; Thorn, C.; Klein, T.; Swen, J.; McLeod, H.; Diasio, R.B.; Schwab, M. Clinical Pharmacogenetics Implementation Consortium guidelines for dihydropyrimidine dehydrogenase genotype and fluoropyrimidine dosing. Clin. Pharmacol. Ther. 2013, 94, 640–645. [Google Scholar] [CrossRef] [PubMed]
- Klein, T.E.; Chang, J.T.; Cho, M.K.; Easton, K.L.; Fergerson, R.; Hewett, M.; Lin, Z.; Liu, Y.; Liu, S.; Oliver, D. Integrating genotype and phenotype information: An overview of the PharmGKB project. Pharm. J. 2001, 1, 167. [Google Scholar] [CrossRef]
- Lamba, J.; Hebert, J.M.; Schuetz, E.G.; Klein, T.E.; Altman, R.B. PharmGKB summary: Very important pharmacogene information for CYP3A5. Pharm. Genom. 2012, 22, 555. [Google Scholar] [CrossRef]
- Deghou, S.; Zeller, G.; Iskar, M.; Castillo, M.; Van Noort, V.; Bork, P. CART- a chemical annotation retrieval toolkit. Bioinformatics 2016, 32, 2869–2871. [Google Scholar] [CrossRef]
- Franceschini, A.; Szklarczyk, D.; Frankild, S.; Kuhn, M.; Simonovic, M.; Roth, A.; Lin, J.; Minguez, P.; Bork, P.; von Mering, C.; et al. STRING v9.1: Protein-protein interaction networks, with increased coverage and integration. Nucleic Acids Res. 2013, 41, D808–D815. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Franceschini, A.; Wyder, S.; Forslund, K.; Heller, D.; Huerta-Cepas, J.; Simonovic, M.; Roth, A.; Santos, A.; Tsafou, K.P.; et al. STRING v10: Protein-protein interaction networks, integrated over the tree of life. Nucleic Acids Res. 2015, 43, D447–D452. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Morris, J.H.; Cook, H.; Kuhn, M.; Wyder, S.; Simonovic, M.; Santos, A.; Doncheva, N.T.; Roth, A.; Bork, P.; et al. The STRING database in 2017: Quality-controlled protein-protein association networks, made broadly accessible. Nucleic Acids Res. 2017, 45, D362–D368. [Google Scholar] [CrossRef] [PubMed]
- Wagstaff, K.; Cardie, C.; Rogers, S.; Schrödl, S. Constrained k-means clustering with background knowledge. In Proceedings of the Eighteenth International Conference on Machine Learning, Williamstown, MA, USA, 28 June–1 July 2001; pp. 577–584. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| APAP | Ethanol | APAP/Ethanol |
|---|---|---|
| TYRP1 | ||
| CYP3A4 | CYP3A4 | CYP3A4 |
| MAPK3 | MAPK3 | MAPK3 |
| DNASE1L3 | ||
| GPSM2 | ||
| PEPD | ||
| ANK | ||
| CYP1A2 | CYP1A2 | CYP1A2 |
| GSS | ||
| NAT2 | ||
| SULT2A1 | SULT2A1 | SULT2A1 |
| CYP2E1 | CYP2E1 | CYP2E1 |
| CDH5 | ||
| GNLY | ||
| DDX10 | ||
| CLEC3B | ||
| CLIC1 | CLIC1 | CLIC1 |
| CYP1A1 | CYP1A1 | CYP1A1 |
| SULT1E1 | SULT1E1 | SULT1E1 |
| HSPA1A | ||
| PARP1 | PARP1 | PARP1 |
| SULT1A1 | SULT1A1 | SULT1A1 |
| AZAMP | ||
| BECN1 | ||
| BIR1 | ||
| C17ORF48 | ||
| GLRA2 | ||
| LMNB1 | ||
| L1CAM | ||
| CD44 | ||
| CYP2C9 | CYP2C9 | CYP2C9 |
| NRCAM | ||
| NID1 | ||
| SLC22A9 | ||
| CASP3 | CASP3 | CASP3 |
| MAPK1 | MAPK1 | MAPK1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hedgpeth, B.; Missall, R.; Bambaci, A.; Smolen, M.; Yavuz, S.; Cottrell, J.; Chu, T.; Chang, S.L. A Review of Bioinformatics Tools to Understand Acetaminophen-Alcohol Interaction. Medicines 2019, 6, 79. https://doi.org/10.3390/medicines6030079
Hedgpeth B, Missall R, Bambaci A, Smolen M, Yavuz S, Cottrell J, Chu T, Chang SL. A Review of Bioinformatics Tools to Understand Acetaminophen-Alcohol Interaction. Medicines. 2019; 6(3):79. https://doi.org/10.3390/medicines6030079
Chicago/Turabian StyleHedgpeth, Bryan, Roy Missall, Anna Bambaci, Matthew Smolen, Sevgi Yavuz, Jessica Cottrell, Tinchun Chu, and Sulie L. Chang. 2019. "A Review of Bioinformatics Tools to Understand Acetaminophen-Alcohol Interaction" Medicines 6, no. 3: 79. https://doi.org/10.3390/medicines6030079
APA StyleHedgpeth, B., Missall, R., Bambaci, A., Smolen, M., Yavuz, S., Cottrell, J., Chu, T., & Chang, S. L. (2019). A Review of Bioinformatics Tools to Understand Acetaminophen-Alcohol Interaction. Medicines, 6(3), 79. https://doi.org/10.3390/medicines6030079