
A Low-Cost Fertilizer Medium Supplemented with Urea for the Lutein Production of Chlorella sp. and the Ability of the Lutein to Protect Cells against Blue Light Irradiation

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microalgal Cultures, Medium, and Chemicals
2.2. Preparation of the Inoculum
2.3. Batch Cultivations of Indoor Photobioreactors
2.4. Determination of Microalgal Cell Biomass and Biomass Productivity
2.5. Determination of Microalgal Lutein Content
2.6. Cell Culture and Medium
2.7. Evaluation of the Ability of Microalgal Lutein to Protect Cells against Blue-Light Irradiation
2.8. Determination of Cell Viability
2.9. Determination of ROS Production
2.10. Statistics
3. Results and Discussion
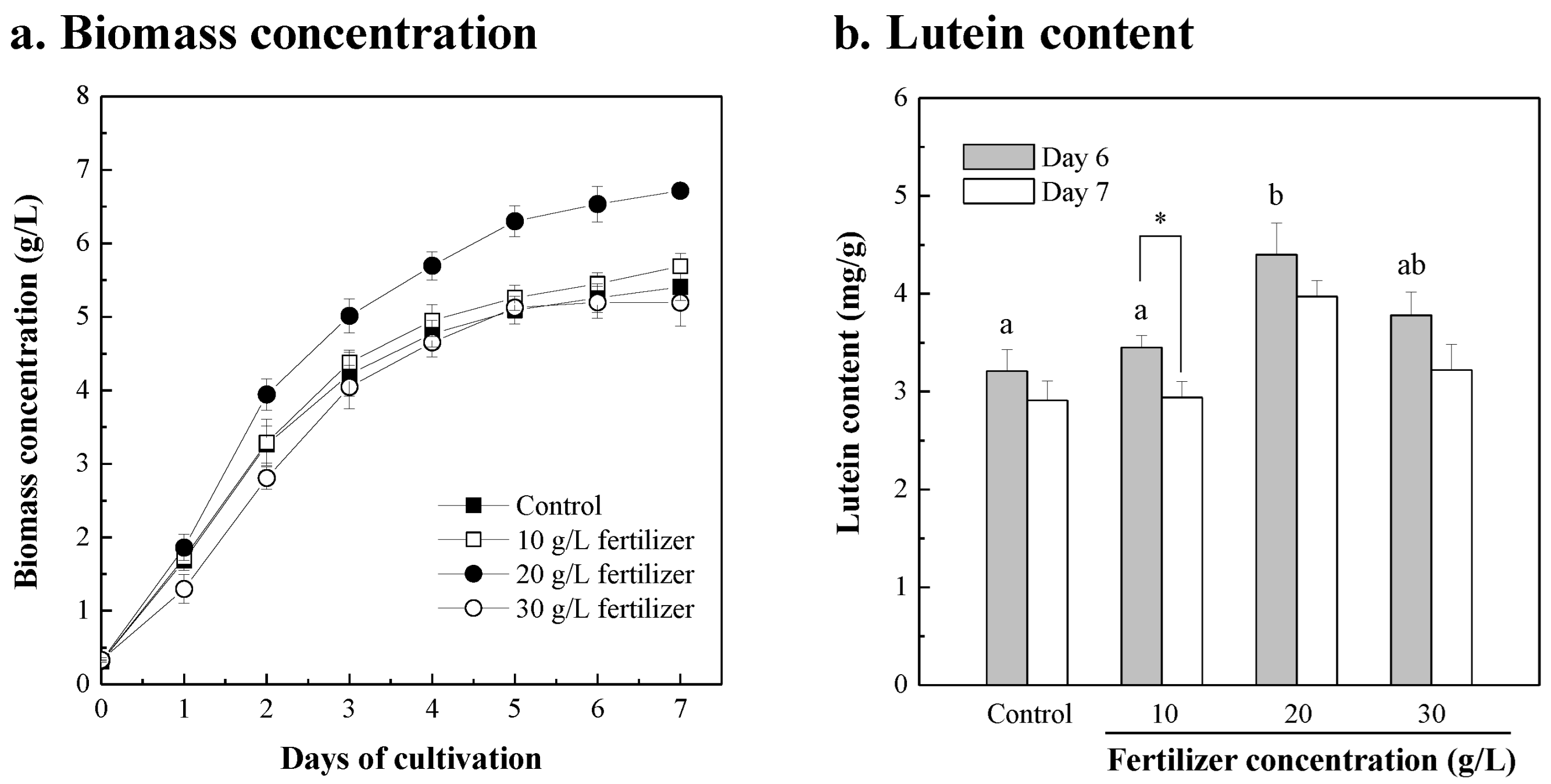
3.1. Chlorella sp. Was Cultivated in Fertilizer
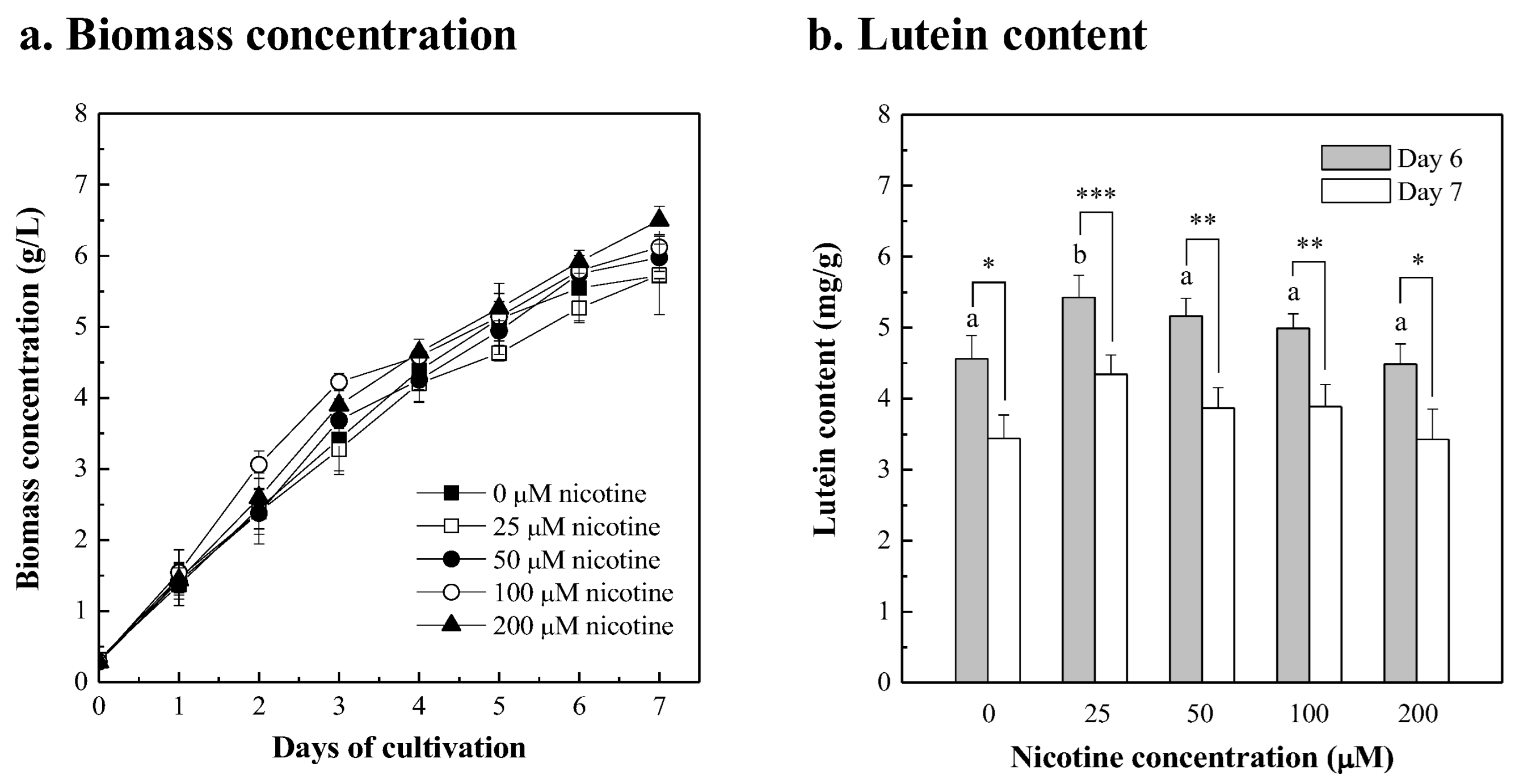
3.2. Chlorella sp. Was Cultured in Fertilizer with the Nicotine Supplement
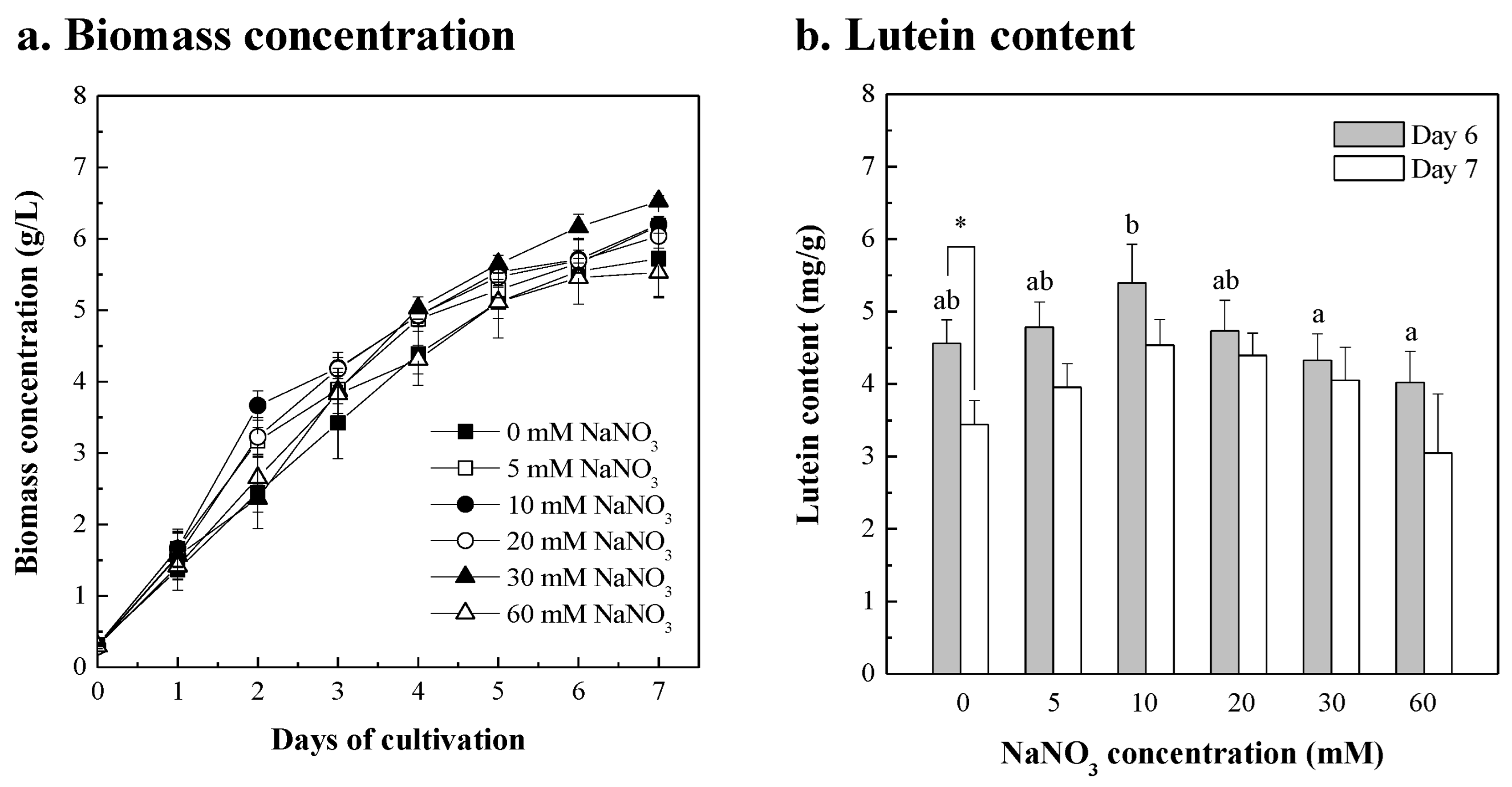
3.3. Chlorella sp. Was Cultured in Fertilizer with NaNO3 Supplement
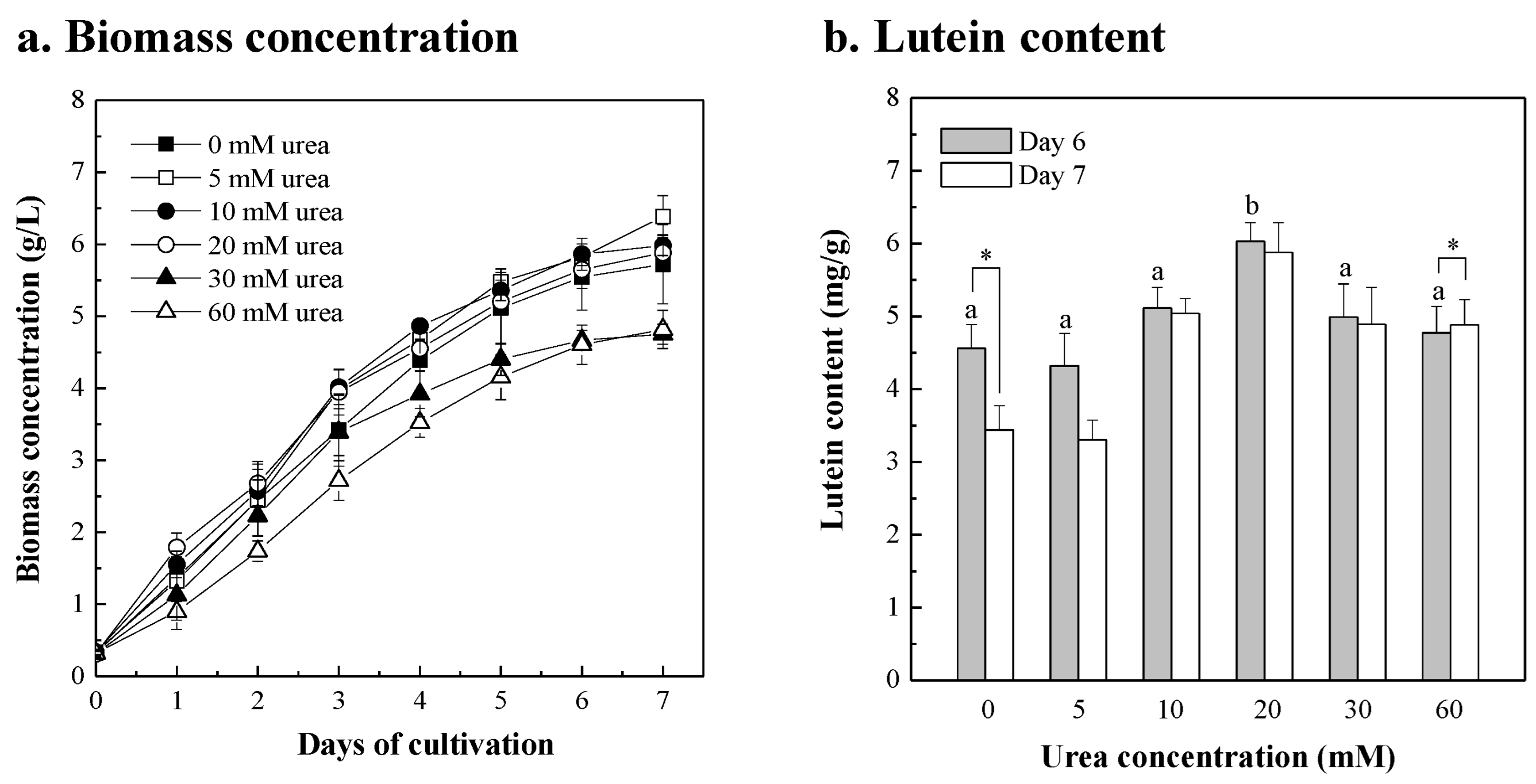
3.4. Chlorella sp. Was Cultured in Fertilizer with the Urea Supplement
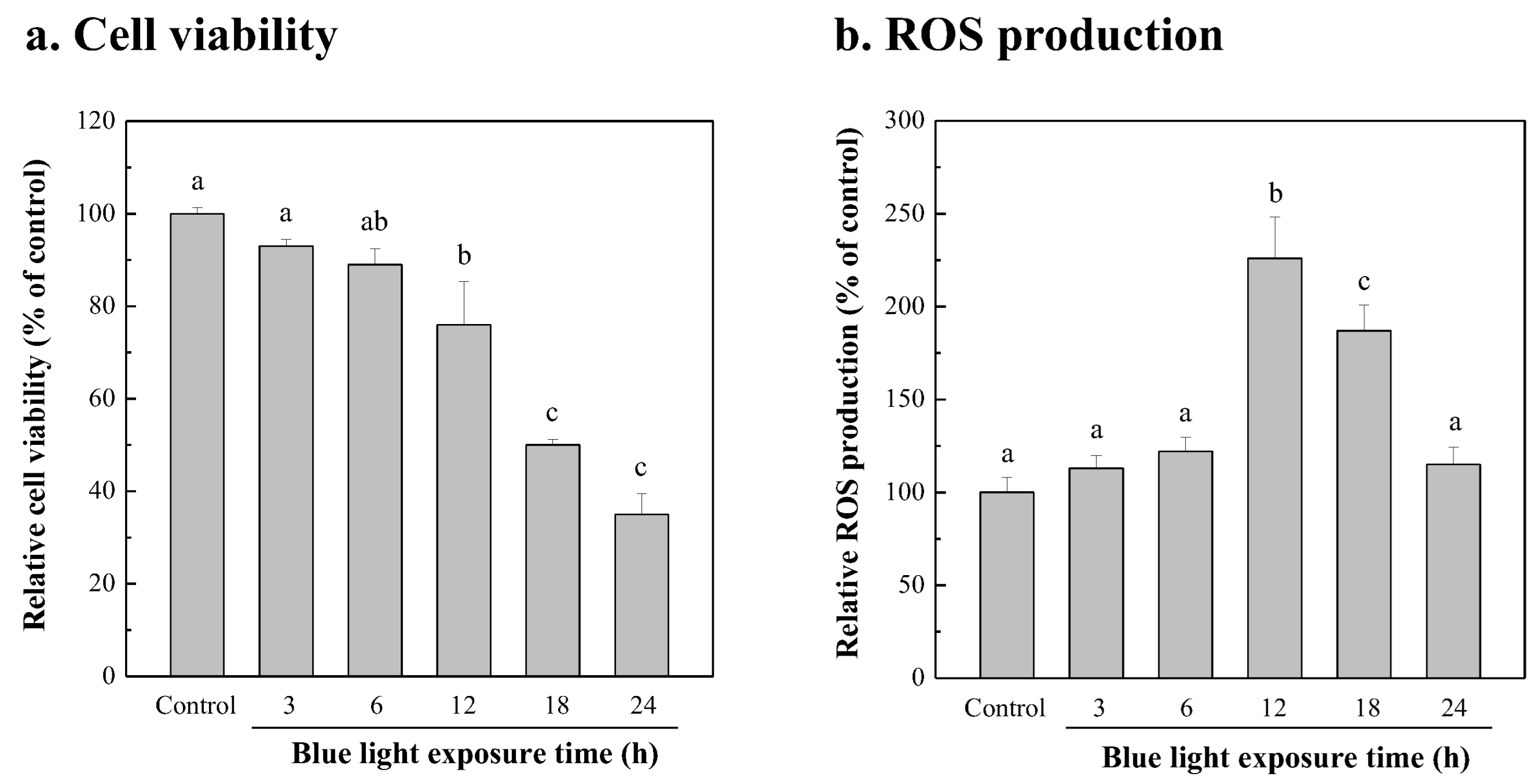
3.5. Damage of NIH/3T3 Cells from Blue-Light Irradiation
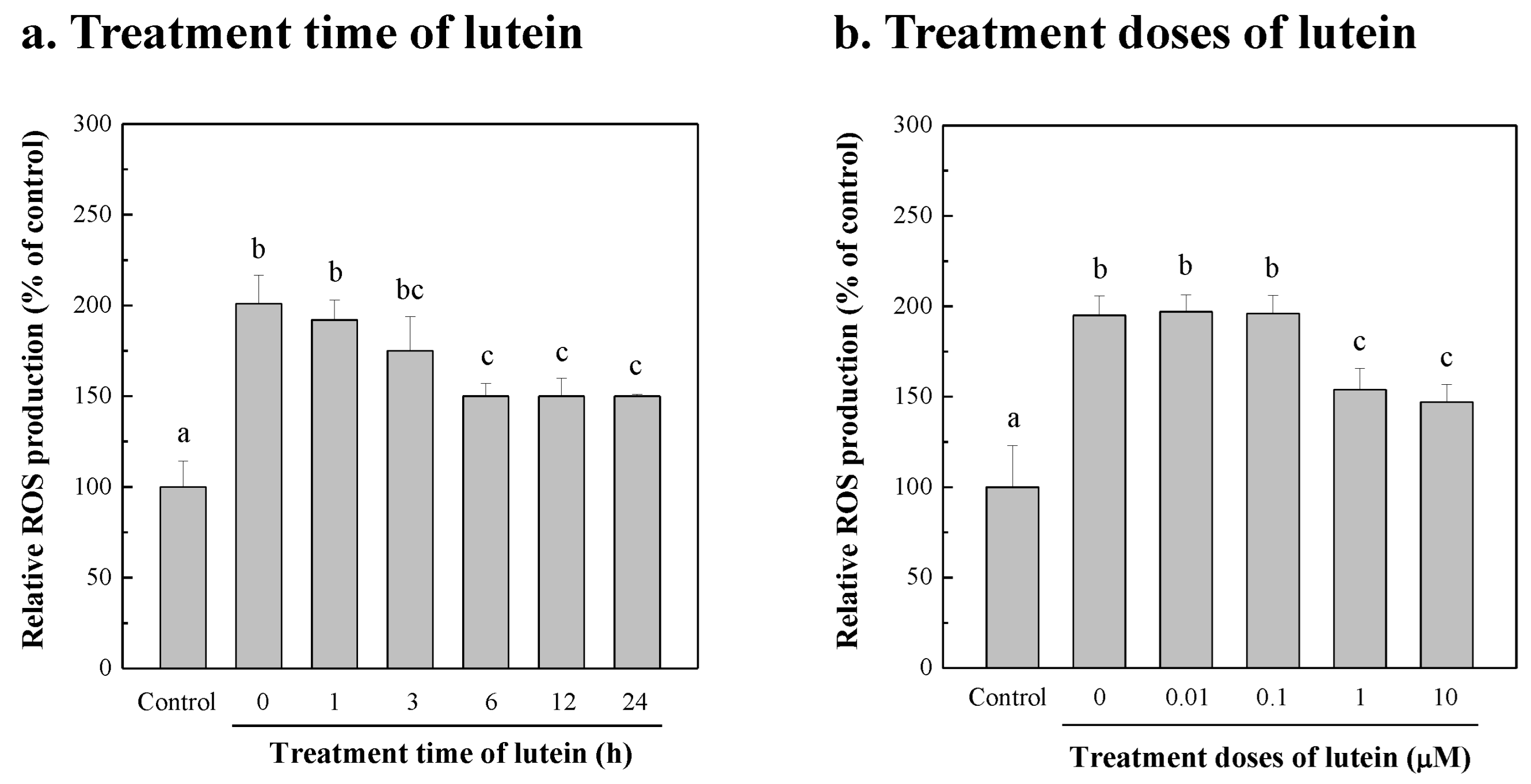
3.6. Protection of NIH/3T3 Cells against Damage from Blue-Light Exposure by Lutein of Chlorella sp.
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jaiswal, K.K.; Dutta, S.; Banerjee, I.; Pohrmen, C.B.; Kumar, V. Photosynthetic microalgae-based carbon sequestration and generation of biomass in biorefinery approach for renewable biofuels for a cleaner environment. Biomass Convers. Biorefinery 2021, 11, 1–19. [Google Scholar] [CrossRef]
- Bibi, R.; Ahmad, Z.; Imran, M.; Hussain, S.; Ditta, A.; Mahmood, S.; Khalid, A. Algal bioethanol production technology: A trend towards sustainable development. Renew. Sustain. Energy Rev. 2017, 71, 976–985. [Google Scholar] [CrossRef]
- Karpagam, R.; Jawaharraj, K.; Gnanam, R. Review on integrated biofuel production from microalgal biomass through the outset of transesterification route: A cascade approach for sustainable bioenergy. Sci. Total Environ. 2021, 766, 144236. [Google Scholar] [CrossRef] [PubMed]
- Cuevas-Castillo, G.A.; Navarro-Pineda, F.S.; Rodríguez, S.A.B.; Rivero, J.C.S. Advances on the processing of microalgal biomass for energy-driven biorefineries. Renew. Sustain. Energy Rev. 2020, 125, 109606. [Google Scholar] [CrossRef]
- Mehariya, S.; Goswami, R.K.; Karthikeysan, O.P.; Verma, P. Microalgae for high-value products: A way towards green nutraceutical and pharmaceutical compounds. Chemosphere 2021, 280, 130553. [Google Scholar] [CrossRef]
- Kuo, C.M.; Sun, Y.L.; Lin, C.H.; Lin, C.H.; Wu, H.T.; Lin, C.S. Cultivation and biorefinery of microalgae (Chlorella sp.) for producing biofuels and other byproducts: A review. Sustainability 2021, 13, 13480. [Google Scholar] [CrossRef]
- Mahmood, A.K.H.; Mohsin, K.E. Experimental study for commercial fertilizer NPK (20:20:20+TE N: P: K) in microalgae cultivation at different aeration periods. Iraqi J. Chem. Pet. Eng. 2017, 18, 99–110. [Google Scholar]
- Bagchi, S.K.; Patnaik, R.; Prasad, R. Feasibility of utilizing wastewaters for large-scale microalgal cultivation and biofuel productions using hydrothermal liquefaction technique: A comprehensive review. Front. Bioeng. Biotechnol. 2021, 9, 651138. [Google Scholar] [CrossRef]
- Xia, A.; Murphy, J.D. Microalgal cultivation in treating liquid digestate from biogas systems. Trends Biotechnol. 2016, 34, 264–275. [Google Scholar] [CrossRef]
- Nayak, M.; Karemore, A.; Sen, R. Performance evaluation of microalgae for concomitant wastewater bioremediation, CO2 biofixation and lipid biosynthesis for biodiesel application. Algal Res. 2016, 16, 216–223. [Google Scholar] [CrossRef]
- Mtaki, K.; Kyewalyanga, M.S.; Mtolera, M.S.P. Supplementing wastewater with NPK fertilizer as a cheap source of nutrients in cultivating live food (Chlorella vulgaris). Ann. Microbiol. 2021, 71, 7. [Google Scholar] [CrossRef]
- Kuo, C.M.; Chen, T.Y.; Lin, T.H.; Kao, C.Y.; Lai, J.T.; Chang, J.S.; Lin, C.S. Cultivation of Chlorella sp. GD using piggery wastewater for biomass and lipid production. Bioresour. Technol. 2015, 194, 326–333. [Google Scholar] [CrossRef]
- Tao, R.; Kinnunen, V.; Praveenkumar, R.; Lakaniemi, A.M.; Rintala, J. Comparison of Scenedesmus acuminatus and Chlorella vulgaris cultivation in liquid digestates from anaerobic digestion of pulp and paper industry and municipal wastewater treatment sludge. Environ. Boil. Fishes 2017, 29, 2845–2856. [Google Scholar] [CrossRef]
- Ge, S.; Qiu, S.; Tremblay, D.; Viner, K.; Champagne, P.; Jessop, P.G. Centrate wastewater treatment with Chlorella vulgaris: Simultaneous enhancement of nutrient removal, biomass and lipid production. Chem. Eng. J. 2018, 342, 310–320. [Google Scholar] [CrossRef]
- Zhang, W.; Li, J.; Zhang, Z.; Fan, G.; Ai, Y.; Gao, Y.; Pan, G. Comprehensive evaluation of a cost-effective method of culturing Chlorella pyrenoidosa with unsterilized piggery wastewater for biofuel production. Biotechnol. Biofuels 2019, 12, 69. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, D.; Rocha, A.C.S.; Garcia, A.; Bóia, A.; Pereira, L.; Verdelhos, T. Municipal wastewater: A sustainable source for the green microalgae Chlorella vulgaris biomass production. Appl. Sci. 2021, 11, 2207. [Google Scholar] [CrossRef]
- Yang, M.; Xue, C.; Li, L.; Gao, Z.; Liu, Q.; Qian, P.; Dong, L.; Gao, K. Design and performance of a low-cost microalgae culturing system for growing Chlorella sorokiniana on cooking cocoon wastewater. Algal Res. 2022, 62, 102607. [Google Scholar] [CrossRef]
- Udaiyappan, A.F.M.; Hasan, H.A.; Takriff, M.S.; Abdullah, S.R.S. A review of the potentials, challenges and current status of microalgae biomass applications in industrial wastewater treatment. J. Water Process. Eng. 2017, 20, 8–21. [Google Scholar] [CrossRef]
- Abdel-Raouf, N.; Al-Homaidan, A.A.; Ibraheem, I.B.M. Microalgae and wastewater treatment. Saudi. J. Biol. Sci. 2012, 19, 257–275. [Google Scholar] [CrossRef]
- Kuo, C.M.; Jian, J.F.; Lin, T.H.; Chang, Y.B.; Wan, X.H.; Lai, J.T.; Chang, J.S.; Lin, C.S. Simultaneous microalgal biomass production and CO2 fixation by cultivating Chlorella sp. GD with aquaculture wastewater and boiler flue gas. Bioresour. Technol. 2016, 221, 241–250. [Google Scholar] [CrossRef]
- Clagnan, E.; D’Imporzano, G.; Dell’Orto, M.; Sanchez-Zurano, A.; Acién-Fernandez, F.G.; Pietrangeli, B.; Adani, F. Profiling microalgal cultures growing on municipal wastewater and fertilizer media in raceway photobioreactors. Bioresour. Technol. 2022, 360, 127619. [Google Scholar] [CrossRef] [PubMed]
- Sipaúba-Tavares, L.H.; Segali, A.M.D.L.; Berchielli-Morais, F.A.; Scardoeli-Truzzi, B. Development of low-cost culture media for Ankistrodesmus gracilis based on inorganic fertilizer and macrophyte. Acta Limnol. Bras. 2017, 29, e5. [Google Scholar] [CrossRef]
- Dang, N.M.; Lee, K. Utilization of organic liquid fertilizer in microalgae cultivation for biodiesel production. Biotechnol. Bioproc. Eng. 2018, 23, 405–414. [Google Scholar] [CrossRef]
- Covell, L.; Machado, M.; Vaz, M.G.M.V.; Soares, J.; Batista, A.D.; Araújo, W.L.; Martins, M.A.; Nunes-Nesi, A. Alternative fertilizer-based growth media support high lipid contents without growth impairment in Scenedesmus obliquus BR003. Bioprocess Biosyst. Eng. 2020, 43, 1123–1131. [Google Scholar] [CrossRef] [PubMed]
- Jbari, N.; Elbaouchi, A.; Benhima, R.; Bennis, I.; Boulif, R.; Zeroual, Y.; Arroussi, H.E. Agriculture fertilizer-based media for cultivation of marine microalgae destined for biodiesel production. J. Energy Manag. Technol. 2020, 4, 49–56. [Google Scholar]
- Ribeiro, D.M.; Roncaratti, L.F.; Possa, G.C.; Garcia, L.C.; Cançado, L.J.; Williams, T.C.R.; Brasil, B.D.S.A.F. A low-cost approach for Chlorella sorokiniana production through combined use of urea, ammonia and nitrate based fertilizers. Bioresour. Technol. Rep. 2020, 9, 100354. [Google Scholar] [CrossRef]
- Koley, S.; Sonkar, S.; Bagchi, S.K.; Patnaik, R.; Mallick, N. Development of a low-cost cultivation medium for simultaneous production of biodiesel and bio-crude from the chlorophycean microalga Tetradesmus obliquus: A renewable energy prospective. J. Clean Prod. 2022, 364, 132658. [Google Scholar] [CrossRef]
- Nayak, M.; Thirunavoukkarasu, M.; Mohanty, R.C. Cultivation of freshwater microalga Scenedesmus sp. using a low-cost inorganic fertilizer for enhanced biomass and lipid yield. J. Gen. Appl. Microbiol. 2016, 62, 7–13. [Google Scholar] [CrossRef]
- Benavides, A.M.S.; Rudin, M.C.; Villalobos, N.; Touloupakis, E.; Torzillo, G. Growth and hydrogen production by three Chlamydomonas strains cultivated in a commercial fertilizer. Int. J. Hydrog. Energy 2019, 44, 9849–9855. [Google Scholar] [CrossRef]
- Kijlstra, A.; Tian, Y.; Kelly, E.R.; Berendschot, T.T.J.M. Lutein: More than just a filter for blue light. Prog. Retin. Eye Res. 2012, 31, 303–315. [Google Scholar] [CrossRef]
- Godley, B.F.; Shamsi, F.A.; Liang, F.Q.; Jarrett, S.G.; Davies, S.; Boulton, M. Blue light induces mitochondrial DNA damage and free radical production in epithelial cells. J. Biol. Chem. 2005, 280, 21061–21066. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Cui, L.; Li, Y.; Choi, J.S.; Choi, J.H.; Li, Z.; Kim, G.E.; Choi, W.; Yoon, K.C. Influence of light emitting diode-derived blue light overexposure on mouse ocular surface. PLoS ONE 2016, 11, e0161041. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, M.; Yako, T.; Kuse, Y.; Inoue, Y.; Nishinaka, A.; Nakamura, S.; Shimazawa, M.; Hara, H. Exposure to excessive blue LED light damages retinal pigment epithelium and photoreceptors of pigmented mice. Exp. Eye Res. 2018, 177, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Cristaldi, M.; Anfuso, C.D.; Spampinato, G.; Rusciano, D.; Lupo, G. Comparative efficiency of lutein and astaxanthin in the protection of human corneal epithelial cells in vitro from blue-violet light photo-oxidative damage. Appl. Sci. 2022, 12, 1268. [Google Scholar] [CrossRef]
- Lutein Market. Available online: https://www.futuremarketinsights.com/reports/lutein-market (accessed on 10 April 2023).
- Manzoor, S.; Rashid, R.; Panda, B.P.; Sharma, V.; Azhar, M. Green extraction of lutein from marigold flower petals, process optimization and its potential to improve the oxidative stability of sunflower oil. Ultrason. Sonochem. 2022, 85, 105994. [Google Scholar] [CrossRef]
- Patel, A.; Rova, U.; Christakopoulos, P.; Matsakas, L. Microalgal lutein biosynthesis: Recent trends and challenges to enhance the lutein content in microalgal cell factories. Front. Mar. Sci. 2022, 9, 1015419. [Google Scholar] [CrossRef]
- Lin, J.H.; Lee, D.J.; Chang, J.S. Lutein production from biomass: Marigold flowers versus microalgae. Bioresour. Technol. 2015, 184, 421–428. [Google Scholar] [CrossRef]
- Fu, Y.; Wang, Y.; Yi, L.; Liu, J.; Yang, S.; Liu, B.; Chen, F.; Sun, H. Lutein production from microalgae: A review. Bioresour. Technol. 2023, 376, 128875. [Google Scholar] [CrossRef]
- Dineshkumar, R.; Dash, S.K.; Sen, R. Process integration for microalgal lutein and biodiesel production with concomitant flue gas CO2 sequestration: A biorefinery model for healthcare, energy and environment. RSC Adv. 2015, 5, 73381–73394. [Google Scholar] [CrossRef]
- Chen, C.Y.; Lu, I.C.; Nagarajan, D.; Chang, C.H.; Ng, I.S.; Lee, D.J.; Chang, J.S. A highly efficient two-stage cultivation strategy for lutein production using heterotrophic culture of Chlorella sorokiniana MB-1-M12. Bioresour. Technol. 2018, 253, 141–147. [Google Scholar] [CrossRef]
- Li, D.; Yuan, Y.; Cheng, D.; Zhao, Q. Effect of light quality on growth rate, carbohydrate accumulation, fatty acid profile and lutein biosynthesis of Chlorella sp. AE10. Bioresour. Technol. 2019, 291, 121783. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.; Zhang, Z.; Ho, S.H.; Ruan, C.; Li, J.; Xie, Y.; Shi, X.; Liu, L.; Chen, J. Two-stage bioprocess for hyper-production of lutein from microalga Chlorella sorokiniana FZU60: Effects of temperature, light intensity, and operation strategies. Algal Res. 2020, 52, 102119. [Google Scholar] [CrossRef]
- Xie, Y.; Zhang, Z.; Ma, R.; Liu, X.; Miao, M.; Ho, S.H.; Chen, J.; Leong, Y.K.; Chang, J.S. High-cell-density heterotrophic cultivation of microalga Chlorella sorokiniana FZU60 for achieving ultra-high lutein production efficiency. Bioresour. Technol. 2022, 365, 128130. [Google Scholar] [CrossRef]
- Zhao, X.; Yan, J.; Yang, T.; Xiong, P.; Zheng, X.; Lu, Y.; Jing, K. Exploring engineering reduced graphene oxide-titanium dioxide (RGO-TiO2) nanoparticles treatment to effectively enhance lutein biosynthesis with Chlorella sorokiniana F31 under different light intensity. Bioresour. Technol. 2022, 348, 126816. [Google Scholar] [CrossRef] [PubMed]
- Vadrale, A.P.; Dong, C.D.; Haldar, D.; Wu, C.H.; Chen, C.W.; Singhania, R.R.; Patel, A.K. Bioprocess development to enhance biomass and lutein production from Chlorella sorokiniana Kh12. Bioresour. Technol. 2023, 370, 128583. [Google Scholar] [CrossRef]
- Kuo, C.M.; Lin, T.H.; Yang, Y.C.; Zhang, W.X.; Lai, J.T.; Wu, H.T.; Chang, J.S.; Lin, C.S. Ability of an alkali-tolerant mutant strain of the microalga Chlorella sp. AT1 to capture carbon dioxide for increasing carbon dioxide utilization efficiency. Bioresour. Technol. 2017, 244, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.C. Producing Lutein from a Microalga Mutant Strain and Studying Biological Effect of the Lutein Against Blue Light Irradiation. Master’s Thesis, National Chiao Tung University, Taiwan, China, 3 July 2018. [Google Scholar]
- Kuo, C.M.; Jian, J.F.; Sun, Y.L.; Lin, T.H.; Yang, Y.C.; Zhang, W.X.; Chang, H.F.; Lai, J.T.; Chang, J.S.; Lin, C.S. An efficient Photobioreactors/Raceway circulating system combined with alkaline-CO2 capturing medium for microalgal cultivation. Bioresour. Technol. 2018, 266, 398–406. [Google Scholar] [CrossRef]
- Xie, Y.; Ho, S.H.; Chen, C.N.N.; Chen, C.Y.; Ng, I.S.; Jing, K.J.; Chang, J.S.; Lu, Y. Phototrophic cultivation of a thermo-tolerant Desmodesmus sp. for lutein production: Effects of nitrate concentration, light intensity and fed-batch operation. Bioresour. Technol. 2013, 144, 435–444. [Google Scholar] [CrossRef]
- Taucher, J.; Baer, S.; Schwerna, P.; Hofmann, D.; Hümmer, M.; Buchholz, R.; Becker, A. Cell disruption and pressurized liquid extraction of carotenoids from microalgae. J. Thermodyn. Catal. 2016, 7, 158. [Google Scholar] [CrossRef]
- van Tonder, A.; Joubert, A.M.; Cromarty, A.D. Limitations of the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) assay when compared to three commonly used cell enumeration assays. BMC Res. Notes 2015, 8, 47. [Google Scholar] [CrossRef]
- Gong, X.; Draper, C.S.; Allison, G.S.; Marisiddaiah, R.; Rubin, L.P. Effects of the macular carotenoid lutein in human retinal pigment epithelial cells. Antioxidants 2017, 6, 100. [Google Scholar] [CrossRef] [PubMed]
- Montel, A.M.; Dos Santos, R.G.; da Costa, P.R.; Silveira-Lacerda, E.P.; Batista, A.A.; Dos Santos, W.G. Neutron activation increases activity of ruthenium-based complexes and induces cell death in glioma cells independent of p53 tumor suppressor gene. Biometals 2017, 30, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Camacho-Rodríguez, J.; Cerón-García, M.C.; González-López, C.; López-Rosales, L.; Contreras-Gómez, A.; Molina-Grima, E. Use of continuous culture to develop an economical medium for the mass production of Isochrysis galbana for aquaculture. J. Appl. Phycol. 2020, 32, 851–863. [Google Scholar] [CrossRef]
- Tokunaga, S.; Morimoto, D.; Koyama, T.; Kubo, Y.; Shiroi, M.; Ohara, K.; Higashine, T.; Mori, Y.; Nakagawa, S.; Sawayama, S. Enhanced lutein production in Chlamydomonas reinhardtii by overexpression of the lycopene epsilon cyclase gene. Appl. Biochem. Biotechnol. 2021, 193, 1967–1978. [Google Scholar] [CrossRef]
- Yildirim, A.; Akgün, İ.H.; Dalay, M. Converted carotenoid production in Dunaliella salina by using cyclization inhibitors 2-methylimidazole and 3-amino-1,2,4-triazole. Turkish J. Biol. 2017, 41, 213–219. [Google Scholar] [CrossRef]
- Saha, S.K.; Ermis, H.; Murray, P. Marine microalgae for potential lutein production. Appl. Sci. 2020, 10, 6457. [Google Scholar] [CrossRef]
- García-Cañedo, J.C.; Cristiani-Urbina, E.; Flores-Ortiz, C.M.; Ponce-Noyola, T.; Esparza-García, F.; Cañizares-Villanueva, R.O. Batch and fed-batch culture of Scenedesmus incrassatulus: Effect over biomass, carotenoid profile and concentration, photosynthetic efficiency and non-photochemical quenching. Algal Res. 2016, 13, 41–52. [Google Scholar] [CrossRef]
- McClure, D.D.; Luiz, A.; Gerber, B.; Barton, G.W.; Kavanagh, J.M. An investigation into the effect of culture conditions on fucoxanthin production using the marine microalgae Phaeodactylum tricornutum. Algal Res. 2018, 29, 41–48. [Google Scholar] [CrossRef]
- Coulombier, N.; Nicolau, E.; Le Déan, L.; Barthelemy, V.; Schreiber, N.; Brun, P.; Lebouvier, N.; Jauffrais, T. Effects of nitrogen availability on the antioxidant activity and carotenoid content of the microalgae Nephroselmis sp. Mar. Drugs 2020, 18, 453. [Google Scholar] [CrossRef]
- Altın, N.; Kutluk, T.; Uyar, B.; Kapucu, N. Effect of different nitrogen sources on the growth and lipid accumulation of Chlorella variabilis. J. Appl. Biol. Sci. 2019, 12, 38–40. [Google Scholar]
- Ho, S.H.; Xie, Y.; Chan, M.C.; Liu, C.C.; Chen, C.Y.; Lee, D.J.; Huang, C.C.; Chang, J.S. Effects of nitrogen source availability and bioreactor operating strategies on lutein production with Scenedesmus obliquus FSP-3. Bioresour. Technol. 2015, 184, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Araujo, G.S.; Silva, J.W.A.; Viana, C.A.S.; Fernandes, F.A.N. Effect of sodium nitrate concentration on biomass and oil production of four microalgae species. Int. J. Sustain. Energy 2019, 39, 41–50. [Google Scholar] [CrossRef]
- Klein, B.C.; Bastos, R.G.; Filho, R.M.; Maciel, M.R.W. Effect of light, CO2 and nitrate concentration on Chlorella vulgaris growth and composition in a flat-plate photobioreactor. Braz. J. Chem. Eng. 2021, 38, 251–263. [Google Scholar] [CrossRef]
- Del Campo, J.A.; Moreno, J.; Rodrıguez, H.; Vargas, M.A.; Rivas, J.; Guerrero, M.G. Carotenoid content of chlorophycean microalgae: Factors determining lutein accumulation in Muriellopsis sp. (Chlorophyta). J. Biotechnol. 2000, 76, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Fatini, M.A.; Basri, E.M.; Maznah, W.O.W. Effect of different nitrogen sources on cell growth and biochemical compositions of Chlorococcum sp. cultivated under laboratory conditions. IOP Conf. Ser.: Earth Environ. Sci. 2021, 711, 012010. [Google Scholar] [CrossRef]
- Sirohi, P.; Verma, H.; Singh, S.K.; Singh, V.K.; Pandey, J.; Khusharia, S.; Kumar, D.; Kaushalendra; Teotia, P.; Kumar, A. Microalgal carotenoids: Therapeutic application and latest approaches to enhance the production. Curr. Issues Mol. Biol. 2022, 44, 6257–6279. [Google Scholar] [CrossRef]
- Kim, H.S.; Park, W.K.; Lee, B.; Seon, G.; Suh, W.I.; Moon, M.; Chang, Y.K. Optimization of heterotrophic cultivation of Chlorella sp. HS2 using screening, statistical assessment, and validation. Sci Rep. 2019, 9, 19383. [Google Scholar] [CrossRef]
- Shi, X.M.; Zhang, X.W.; Chen, F. Heterotrophic production of biomass and lutein by Chlorella protothecoides on various nitrogen sources. Enzyme Microb. Technol. 2000, 27, 312–318. [Google Scholar] [CrossRef]
- Kumbhar, A.N.; He, M.; Rajper, A.R.; Memon, K.A.; Rizwan, M.; Nagi, M.; Woldemicael, A.G.; Li, D.; Wang, C.; Wang, C. The use of urea and kelp waste extract is a promising strategy for maximizing the biomass productivity and lipid content in Chlorella sorokiniana. Plants 2020, 9, 463. [Google Scholar] [CrossRef]
- Yu, B.S.; Sung, Y.J.; Hong, M.E.; Sim, S.J. Improvement of photoautotrophic algal biomass production after interrupted CO2 supply by urea and KH2PO4 injection. Energies 2021, 14, 778. [Google Scholar] [CrossRef]
- Lockwood, D.B.; Wataha, J.C.; Lewis, J.B.; Tseng, W.Y.; Messer, R.L.; Hsu, S.D. Blue light generates reactive oxygen species (ROS) differentially in tumor vs. normal epithelial cells. Dent. Mater. 2005, 21, 683–688. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, A.; Shiotsu-Ogura, Y.; Wada-Takahashi, S.; Takahashi, S.S.; Toyama, T.; Yoshino, F. Blue light irradiation-induced oxidative stress in vivo via ROS generation in rat gingival tissue. J. Photochem. Photobiol. B-Biol. 2015, 151, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Li, D.; Zhang, Y.; Zhang, L.; Liao, Z.; Aihemaitijiang, S.; Hou, Y.; Zhan, Z.; Xie, K.; Zhang, Z. Lutein protected the retina from light induced retinal damage by inhibiting increasing oxidative stress and inflammation. J. Funct. Food 2020, 73, 104107. [Google Scholar] [CrossRef]
- Algan, A.H.; Gungor-Ak, A.; Karatas, A. Nanoscale delivery systems of lutein: An updated review from a pharmaceutical perspective. Pharmaceutics 2022, 14, 1852. [Google Scholar] [CrossRef]
- Li, X.; Holt, R.R.; Keen, C.L.; Morse, L.S.; Yiu, G.; Hackman, R.M. Goji berry intake increases macular pigment optical density in healthy adults: A randomized pilot trial. Nutrients 2021, 13, 4409. [Google Scholar] [CrossRef] [PubMed]
- Maheshwari, N.; Arya, R.K.; Verros, G.D.; Dhamole, P.B.; Kannan, A. Surfactant-enhanced extraction of lutein from marigold petals using an aqueous two-phase system. Separations 2023, 10, 133. [Google Scholar] [CrossRef]
- Bian, Q.; Zhou, P.; Yao, Z.; Li, M.; Yu, H.; Ye, L. Heterologous biosynthesis of lutein in S. cerevisiae enabled by temporospatial pathway control. Metab. Eng. 2021, 67, 19–28. [Google Scholar] [CrossRef]
- Takemura, M.; Kubo, A.; Watanabe, A.; Sakuno, H.; Minobe, Y.; Sahara, T.; Murata, M.; Araki, M.; Harada, H.; Terada, Y.; et al. Pathway engineering for high-yield production of lutein in Escherichia coli. Synth. Biol. 2021, 6, ysab012. [Google Scholar] [CrossRef]
- Diaz-MacAdoo, D.; Mata, M.T.; Riquelme, C. Influence of irradiance and wavelength on the antioxidant activity and carotenoids accumulation in Muriellopsis sp. isolated from the antofagasta coastal desert. Molecules 2022, 27, 2412. [Google Scholar] [CrossRef]
- Ávila-Román, J.; García-Gil, S.; Rodríguez-Luna, A.; Motilva, V.; Talero, E. Anti-inflammatory and anticancer effects of microalgal carotenoids. Mar. Drugs 2021, 19, 531. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Medium Costs | Modified f/2 Medium | 20 g/L Fertilizer | 20 g/L Fertilizer + 20 mM Urea |
|---|---|---|---|
| Cost per L working volume (US$) | 0.276 | 0.011 | 0.023 |
| Cost reduction (%) 1 | 96 | 92 | |
| Cost per g of biomass production (US$) | 0.052 | 0.002 | 0.004 |
| Cost reduction (%) 2 | 97 | 92 | |
| Cost per g of lutein production (US$) | 16.35 | 0.382 | 0.665 |
| Cost reduction (%) 3 | 98 | 96 |
| Extra Added Components | Concentration Added | Biomass Concentration (g/L) | Biomass Productivity 1 (g/L/d) | Max. Lutein Content 2 (mg/g) | Lutein Productivity (mg/L/d) | ||||
|---|---|---|---|---|---|---|---|---|---|
| Control 3 | 5.547 ± 0.464 | ad | 0.871 ± 0.079 | ad | 4.561 ± 0.327 | bc | 3.986 ± 0.564 | bd | |
| Nicotine | 25 μM | 5.263 ± 0.203 | ab | 0.826 ± 0.030 | ab | 5.425 ± 0.315 | ac | 4.472 ± 0.098 | abcd |
| 50 μM | 5.747 ± 0.100 | ad | 0.910 ± 0.019 | ad | 5.163 ± 0.250 | ac | 4.701 ± 0.304 | abcd | |
| 100 μM | 5.791 ± 0.121 | ad | 0.917 ± 0.020 | ad | 4.992 ± 0.203 | abc | 4.581 ± 0.280 | abcd | |
| 200 μM | 5.916 ± 0.163 | ad | 0.939 ± 0.026 | ad | 4.486 ± 0.284 | bc | 4.209 ± 0.255 | abcd | |
| NaNO3 | 5 mM | 5.658 ± 0.140 | ad | 0.891 ± 0.023 | ad | 4.784 ± 0.347 | bc | 4.268 ± 0.420 | abcd |
| 10 mM | 5.710 ± 0.132 | ad | 0.901 ± 0.023 | ad | 5.396 ± 0.533 | ac | 4.867 ± 0.563 | cd | |
| 20 mM | 5.694 ± 0.034 | ad | 0.901 ± 0.010 | ad | 4.732 ± 0.424 | bc | 4.259 ± 0.339 | abcd | |
| 30 mM | 6.165 ± 0.177 | d | 0.977 ± 0.023 | d | 4.326 ± 0.366 | bc | 4.226 ± 0.326 | abcd | |
| 60 mM | 5.456 ± 0.075 | acd | 0.860 ± 0.011 | acd | 4.022 ± 0.428 | b | 3.458 ± 0.413 | b | |
| Urea | 5 mM | 5.822 ± 0.116 | ad | 0.915 ± 0.023 | ad | 4.322 ± 0.443 | bc | 3.955 ± 0.459 | bd |
| 10 mM | 5.863 ± 0.222 | ad | 0.921 ± 0.030 | ad | 5.114 ± 0.286 | abc | 4.712 ± 0.315 | abcd | |
| 20 mM | 5.648 ± 0.258 | ad | 0.890 ± 0.036 | ad | 6.031 ± 0.258 | a | 5.363 ± 0.231 | ac | |
| 30 mM | 4.667 ± 0.140 | bc | 0.725 ± 0.020 | bc | 4.990 ± 0.456 | abc | 3.611 ± 0.225 | bd | |
| 60 mM | 4.606 ± 0.272 | b | 0.714 ± 0.053 | b | 4.776 ± 0.362 | bc | 3.423 ± 0.497 | b | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuo, C.-M.; Yang, Y.-C.; Zhang, W.-X.; Wu, J.-X.; Chen, Y.-T.; Lin, C.-H.; Lin, M.-W.; Lin, C.-S. A Low-Cost Fertilizer Medium Supplemented with Urea for the Lutein Production of Chlorella sp. and the Ability of the Lutein to Protect Cells against Blue Light Irradiation. Bioengineering 2023, 10, 594. https://doi.org/10.3390/bioengineering10050594
Kuo C-M, Yang Y-C, Zhang W-X, Wu J-X, Chen Y-T, Lin C-H, Lin M-W, Lin C-S. A Low-Cost Fertilizer Medium Supplemented with Urea for the Lutein Production of Chlorella sp. and the Ability of the Lutein to Protect Cells against Blue Light Irradiation. Bioengineering. 2023; 10(5):594. https://doi.org/10.3390/bioengineering10050594
Chicago/Turabian StyleKuo, Chiu-Mei, Yi-Chun Yang, Wen-Xin Zhang, Jia-Xun Wu, Yu-Tso Chen, Cheng-Han Lin, Meng-Wei Lin, and Chih-Sheng Lin. 2023. "A Low-Cost Fertilizer Medium Supplemented with Urea for the Lutein Production of Chlorella sp. and the Ability of the Lutein to Protect Cells against Blue Light Irradiation" Bioengineering 10, no. 5: 594. https://doi.org/10.3390/bioengineering10050594
APA StyleKuo, C. -M., Yang, Y. -C., Zhang, W. -X., Wu, J. -X., Chen, Y. -T., Lin, C. -H., Lin, M. -W., & Lin, C. -S. (2023). A Low-Cost Fertilizer Medium Supplemented with Urea for the Lutein Production of Chlorella sp. and the Ability of the Lutein to Protect Cells against Blue Light Irradiation. Bioengineering, 10(5), 594. https://doi.org/10.3390/bioengineering10050594