
Bioengineered Organoids Offer New Possibilities for Liver Cancer Studies: A Review of Key Milestones and Challenges

 , , ,
, , ,  , , , , ,
, , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
Current Methodologies for the Detection and Treatment of Hepatocellular Carcinoma
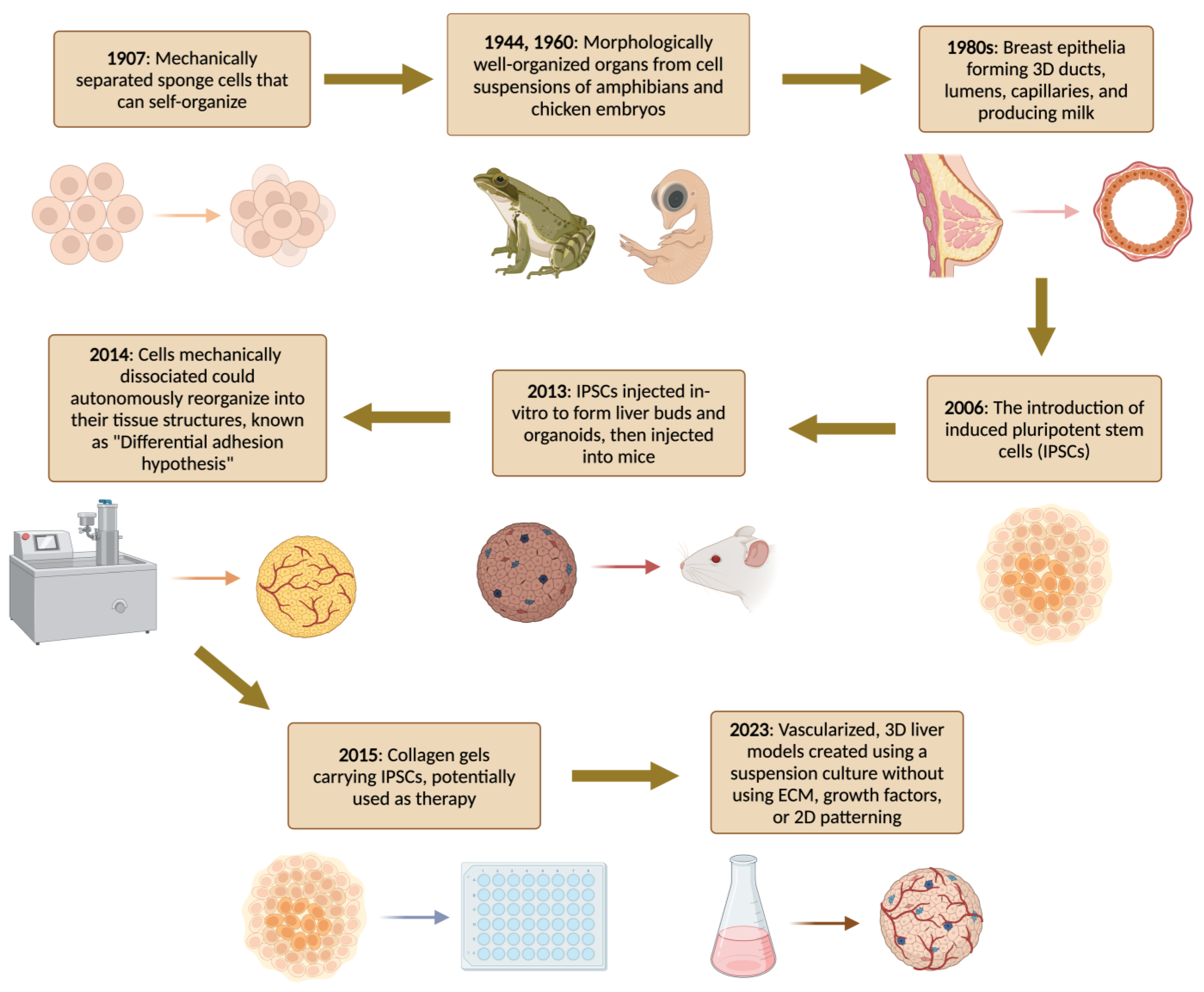
2. A Brief History of Organoids

Overview of the Organoids Concept
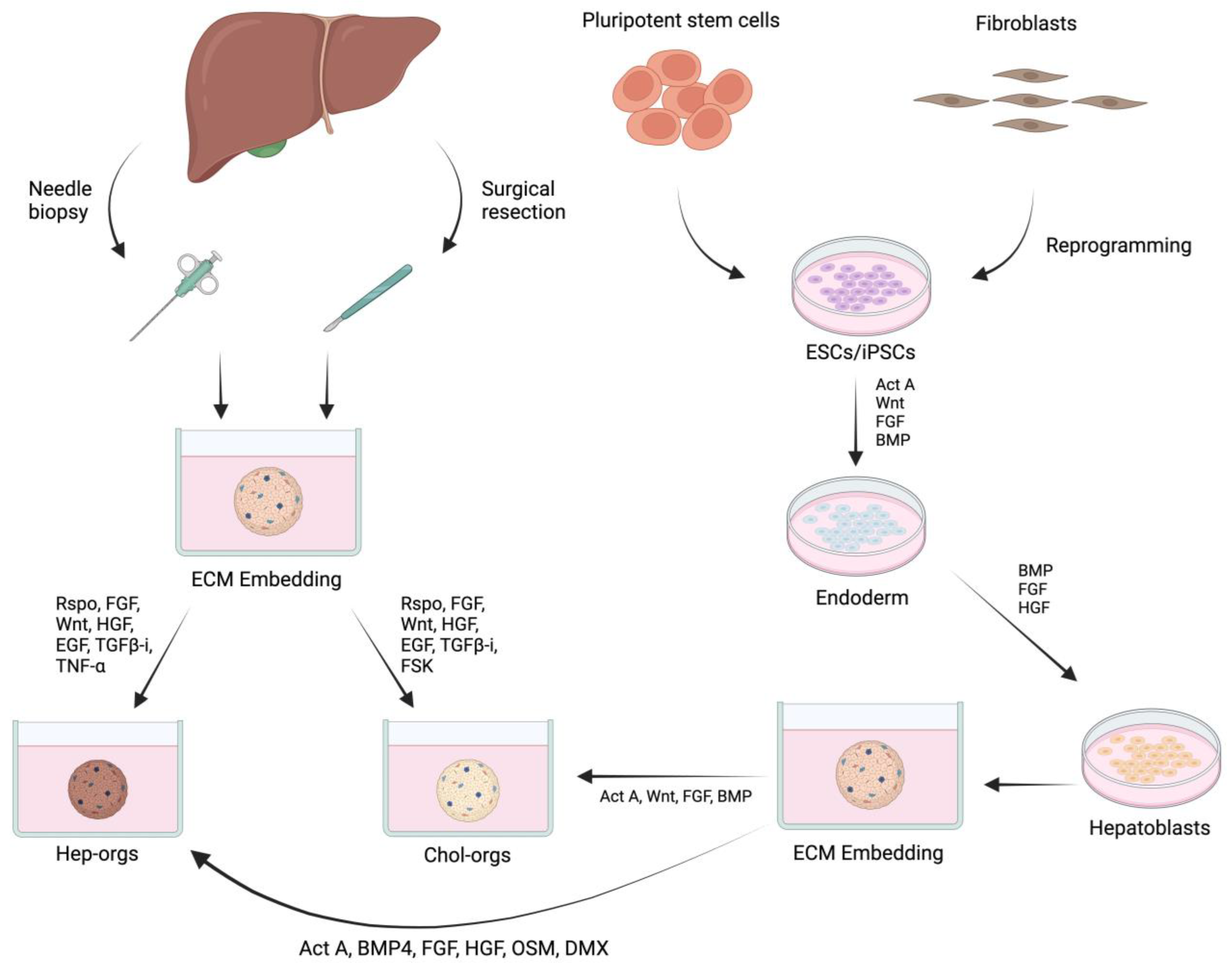
3. Cell Source and Biological Factors for Growing Liver Organoids
4. Bioengineering Strategies for Growing Liver Organoids
Microfabrication Strategies to Control the Assembly of Organoids
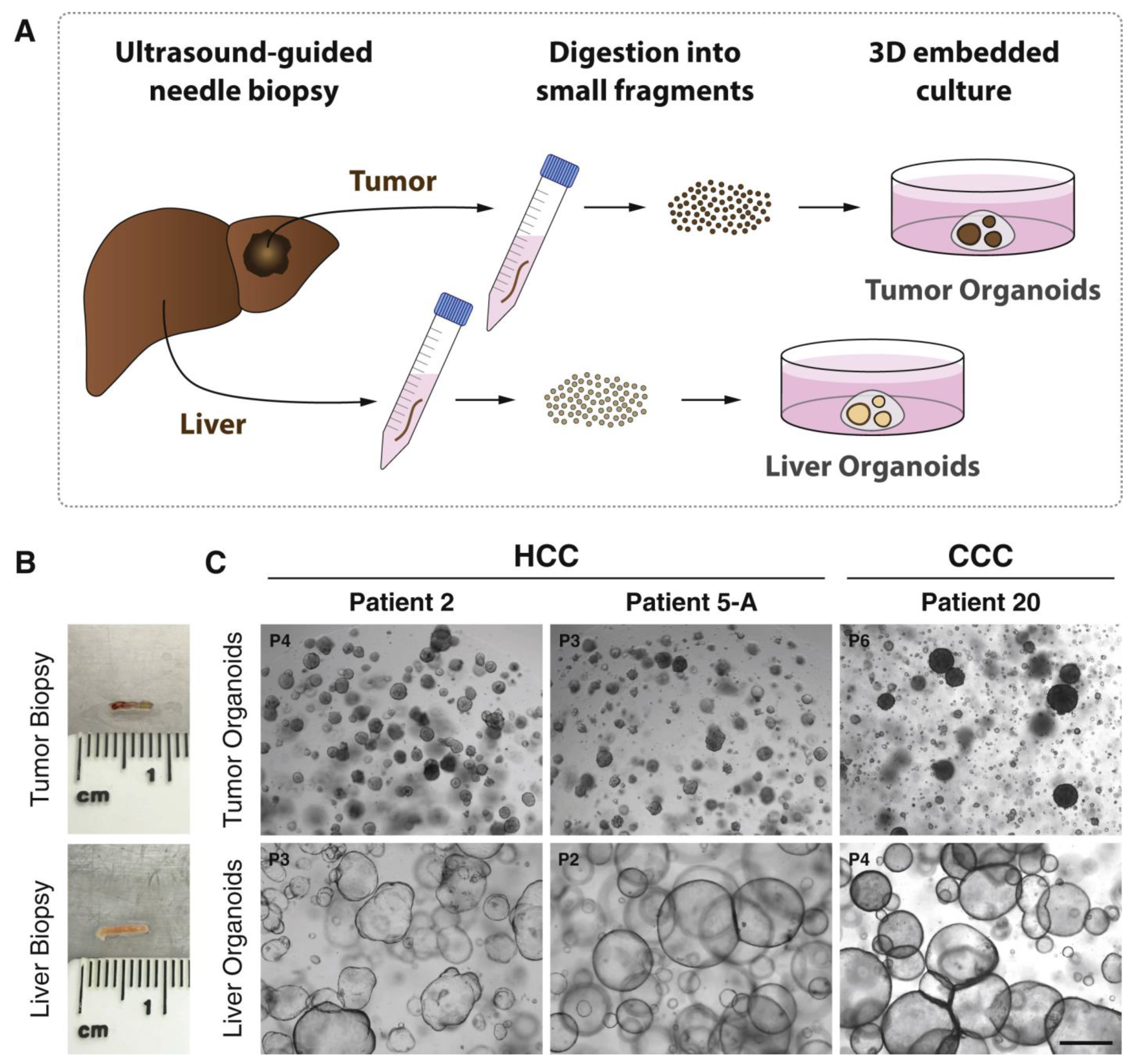
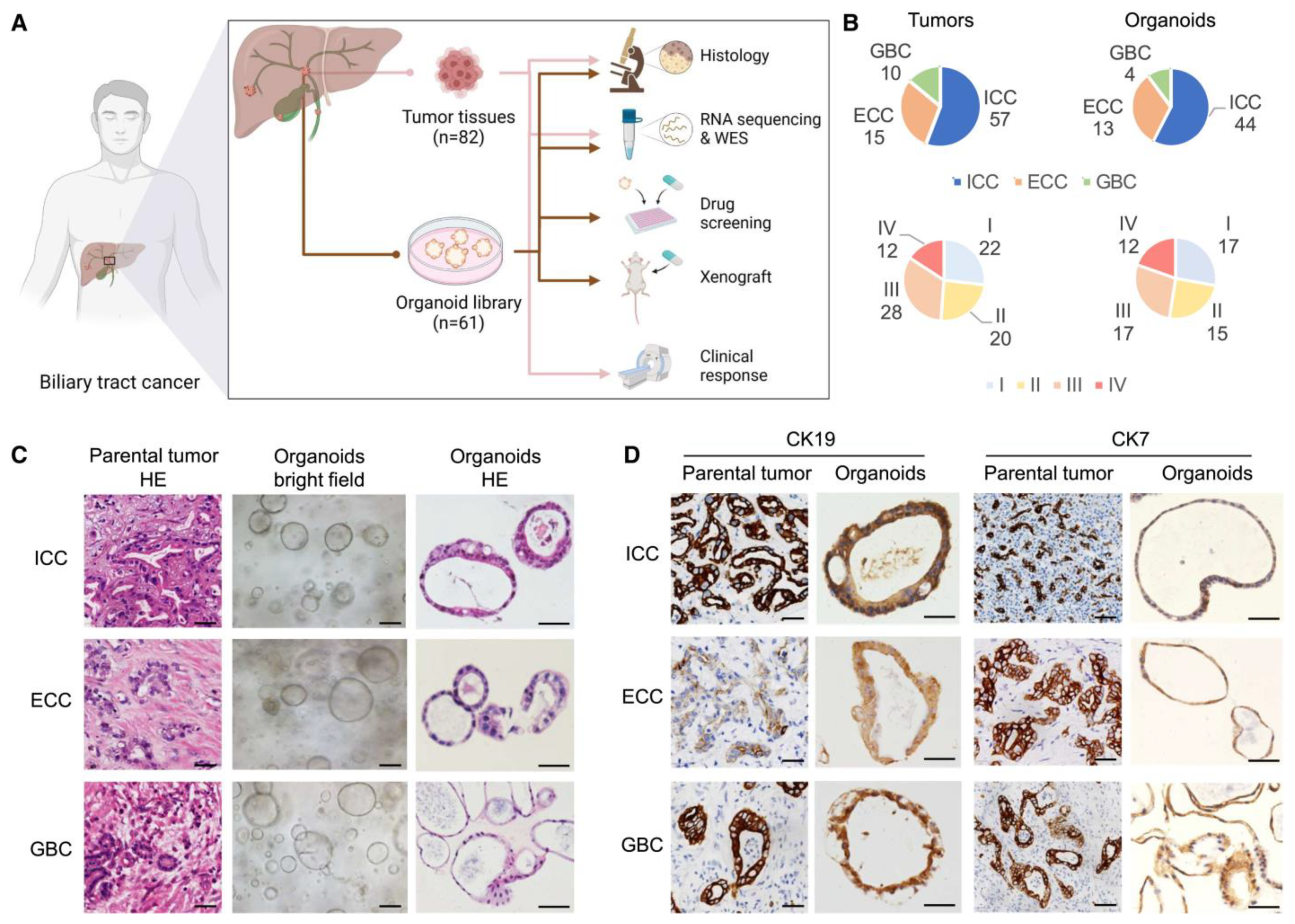
5. Organoids for Liver Cancer Research


Current Limitations of Organoid Models
6. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Sia, D.; Villanueva, A.; Friedman, S.L.; Llovet, J.M. Liver Cancer Cell of Origin, Molecular Class, and Effects on Patient Prognosis. Gastroenterology 2017, 152, 745–761. [Google Scholar] [CrossRef] [PubMed]
- Brindley, P.J.; Bachini, M.; Ilyas, S.I.; Khan, S.A.; Loukas, A.; Sirica, A.E.; Teh, B.T.; Wongkham, S.; Gores, G.J. Cholangiocarcinoma. Nat. Rev. Dis. Primers 2021, 7, 65. [Google Scholar] [CrossRef]
- Sripa, B.; Tangkawattana, S.; Brindley, P.J. Update on Pathogenesis of Opisthorchiasis and Cholangiocarcinoma. Adv. Parasitol. 2018, 102, 97–113. [Google Scholar]
- Rumgay, H.; Ferlay, J.; de Martel, C.; Georges, D.; Ibrahim, A.S.; Zheng, R.; Wei, W.; Lemmens, V.E.; Soerjomataram, I. Global, Regional and National Burden of Primary Liver Cancer by Subtype. Eur. J. Cancer 2022, 161, 108–118. [Google Scholar] [CrossRef] [PubMed]
- Mondaca, S.; Yarmohammadi, H.; Kemeny, N.E. Regional Chemotherapy for Biliary Tract Tumors and Hepatocellular Carcinoma. Surg. Oncol. Clin. 2019, 28, 717–729. [Google Scholar] [CrossRef]
- Chinnappan, R.; Mir, T.A.; Alsalameh, S.; Makhzoum, T.; Alzhrani, A.; Al-Kattan, K.; Yaqinuddin, A. Low-Cost Point-of-Care Monitoring of ALT and AST Is Promising for Faster Decision Making and Diagnosis of Acute Liver Injury. Diagnostics 2023, 13, 2967. [Google Scholar] [CrossRef]
- Chinnappan, R.; Mir, T.A.; Alsalameh, S.; Makhzoum, T.; Adeeb, S.; Al-Kattan, K.; Yaqinuddin, A. Aptasensors Are Conjectured as Promising ALT and AST Diagnostic Tools for the Early Diagnosis of Acute Liver Injury. Life 2023, 13, 1273. [Google Scholar] [CrossRef] [PubMed]
- Kashefi-Kheyrabadi, L.; Mehrgardi, M.A.; Wiechec, E.; Turner, A.P.F.; Tiwari, A. Ultrasensitive Detection of Human Liver Hepatocellular Carcinoma Cells Using a Label-Free Aptasensor. Anal. Chem. 2014, 86, 4956–4960. [Google Scholar] [CrossRef]
- Park, S.; Cho, E.; Chueng, S.-T.D.; Yoon, J.-S.; Lee, T.; Lee, J.-H. Aptameric Fluorescent Biosensors for Liver Cancer Diagnosis. Biosensors 2023, 13, 617. [Google Scholar] [CrossRef]
- Ohashi, K.; Yokoyama, T.; Yamato, M.; Kuge, H.; Kanehiro, H.; Tsutsumi, M.; Amanuma, T.; Iwata, H.; Yang, J.; Okano, T. Engineering Functional Two-and Three-Dimensional Liver Systems In Vivo Using Hepatic Tissue Sheets. Nat. Med. 2007, 13, 880–885. [Google Scholar] [CrossRef]
- Smith, A.J.; Lilley, E. The Role of the Three Rs in Improving the Planning and Reproducibility of Animal Experiments. Animals 2019, 9, 975. [Google Scholar] [CrossRef]
- Langer, R.; Vacanti, J.P. Tissue Engineering. Science 1993, 260, 920–926. [Google Scholar] [CrossRef]
- Lauschke, V.M.; Hendriks, D.F.; Bell, C.C.; Andersson, T.B.; Ingelman-Sundberg, M. Novel 3D Culture Systems for Studies of Human Liver Function and Assessments of the Hepatotoxicity of Drugs and Drug Candidates. Chem. Res. Toxicol. 2016, 29, 1936–1955. [Google Scholar] [CrossRef] [PubMed]
- Mir, T.A.; Nakamura, M. Three-Dimensional Bioprinting: Toward the Era of Manufacturing Human Organs as Spare Parts for Healthcare and Medicine. Tissue Eng. Part B Rev. 2017, 23, 245–256. [Google Scholar] [CrossRef]
- Arai, K.; Yoshida, T.; Okabe, M.; Goto, M.; Mir, T.A.; Soko, C.; Tsukamoto, Y.; Akaike, T.; Nikaido, T.; Zhou, K. Fabrication of 3D-culture Platform with Sandwich Architecture for Preserving Liver-specific Functions of Hepatocytes Using 3D Bioprinter. J. Biomed. Mater. Res. Part A 2017, 105, 1583–1592. [Google Scholar] [CrossRef] [PubMed]
- Mir, T.A.; Iwanaga, S.; Kurooka, T.; Toda, H.; Sakai, S.; Nakamura, M. Biofabrication Offers Future Hope for Tackling Various Obstacles and Challenges in Tissue Engineering and Regenerative Medicine: A Perspective. Int. J. Bioprinting 2019, 5, 153. [Google Scholar] [CrossRef]
- Nair, D.G.; Weiskirchen, R. Recent Advances in Liver Tissue Engineering as an Alternative and Complementary Approach for Liver Transplantation. Curr. Issues Mol. Biol. 2024, 46, 262–278. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, M.; Mir, T.A.; Arai, K.; Ito, S.; Yoshida, T.; Iwanaga, S.; Kitano, H.; Obara, C.; Nikaido, T. Bioprinting with Pre-Cultured Cellular Constructs towards Tissue Engineering of Hierarchical Tissues. Int. J. Bioprinting 2015, 1, 39–48. [Google Scholar]
- Kim, J.; Kang, W.; Sinn, D.H.; Gwak, G.-Y.; Paik, Y.-H.; Choi, M.S.; Lee, J.H.; Koh, K.C.; Paik, S.W. Substantial Risk of Recurrence Even after 5 Recurrence-Free Years in Early-Stage Hepatocellular Carcinoma Patients. Clin. Mol. Hepatol. 2020, 26, 516. [Google Scholar] [CrossRef]
- Kulik, L.; El-Serag, H.B. Epidemiology and Management of Hepatocellular Carcinoma. Gastroenterology 2019, 156, 477–491. [Google Scholar] [CrossRef] [PubMed]
- Chernyak, V.; Fowler, K.J.; Kamaya, A.; Kielar, A.Z.; Elsayes, K.M.; Bashir, M.R.; Kono, Y.; Do, R.K.; Mitchell, D.G.; Singal, A.G. Liver Imaging Reporting and Data System (LI-RADS) Version 2018: Imaging of Hepatocellular Carcinoma in at-Risk Patients. Radiology 2018, 289, 816–830. [Google Scholar] [CrossRef]
- Faccia, M.; Garcovich, M.; Ainora, M.E.; Riccardi, L.; Pompili, M.; Gasbarrini, A.; Zocco, M.A. Contrast-Enhanced Ultrasound for Monitoring Treatment Response in Different Stages of Hepatocellular Carcinoma. Cancers 2022, 14, 481. [Google Scholar] [CrossRef] [PubMed]
- Bartolotta, T.V.; Taibbi, A.; Picone, D.; Anastasi, A.; Midiri, M.; Lagalla, R. Detection of Liver Metastases in Cancer Patients with Geographic Fatty Infiltration of the Liver: The Added Value of Contrast-Enhanced Sonography. Ultrasonography 2017, 36, 160. [Google Scholar] [CrossRef]
- Simmons, O.; Fetzer, D.T.; Yokoo, T.; Marrero, J.A.; Yopp, A.; Kono, Y.; Parikh, N.; Browning, T.; Singal, A.G. Predictors of Adequate Ultrasound Quality for Hepatocellular Carcinoma Surveillance in Patients with Cirrhosis. Aliment. Pharmacol. Ther. 2017, 45, 169–177. [Google Scholar] [CrossRef]
- Gregory, J.; Burgio, M.D.; Corrias, G.; Vilgrain, V.; Ronot, M. Evaluation of Liver Tumour Response by Imaging. JHEP Rep. 2020, 2, 100100. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.T.; Tan, N.E.; Volk, M.L. Utility of Positron Emission Tomography-Computed Tomography Scan in Detecting Residual Hepatocellular Carcinoma Post Treatment: Series of Case Reports. World J. Gastrointest. Oncol. 2020, 12, 358. [Google Scholar] [CrossRef]
- Shao, Y.; Liu, T.; Hsu, C.; Lu, L.; Shen, Y.; Lin, Z.; Cheng, A.; Hsu, C. Early Alpha-foetoprotein Response Associated with Treatment Efficacy of Immune Checkpoint Inhibitors for Advanced Hepatocellular Carcinoma. Liver Int. 2019, 39, 2184–2189. [Google Scholar] [CrossRef]
- Piñero, F.; Dirchwolf, M.; Pessôa, M.G. Biomarkers in Hepatocellular Carcinoma: Diagnosis, Prognosis and Treatment Response Assessment. Cells 2020, 9, 1370. [Google Scholar] [CrossRef]
- Kim, H.I.; Lim, J.; Shim, J.H. Role of the Alpha-Fetoprotein Response in Immune Checkpoint Inhibitor-Based Treatment of Patients with Hepatocellular Carcinoma. J. Cancer Res. Clin. Oncol. 2022, 148, 2069–2077. [Google Scholar] [CrossRef]
- He, C.; Peng, W.; Liu, X.; Li, C.; Li, X.; Wen, T.-F. Post-Treatment Alpha-Fetoprotein Response Predicts Prognosis of Patients with Hepatocellular Carcinoma: A Meta-Analysis. Medicine 2019, 98, e16557. [Google Scholar] [CrossRef]
- Wang, T.; Zhang, K.-H. New Blood Biomarkers for the Diagnosis of AFP-Negative Hepatocellular Carcinoma. Front. Oncol. 2020, 10, 1316. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Ha, S.-A.; Kim, H.K.; Yoo, J.; Kim, S.; Lee, Y.S.; Hur, S.Y.; Kim, Y.W.; Kim, T.E.; Park, Y.G. Combined Analysis of AFP and HCCR-1 as an Useful Serological Marker for Small Hepatocellular Carcinoma: A Prospective Cohort Study. Dis. Markers 2012, 32, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.; Li, B.; Li, Z.; Wei, Q.; Ren, L. PIVKA-II Serves as a Potential Biomarker That Complements AFP for the Diagnosis of Hepatocellular Carcinoma. BMC Cancer 2021, 21, 401. [Google Scholar] [CrossRef] [PubMed]
- Omar, M.A.; Omran, M.M.; Farid, K.; Tabll, A.A.; Shahein, Y.E.; Emran, T.M.; Petrovic, A.; Lucic, N.R.; Smolic, R.; Kovac, T.; et al. Biomarkers for Hepatocellular Carcinoma: From Origin to Clinical Diagnosis. Biomedicines 2023, 11, 1852. [Google Scholar] [CrossRef]
- Zhu, H.-F.; Feng, J.-K.; Xiang, Y.-J.; Wang, K.; Zhou, L.-P.; Liu, Z.-H.; Cheng, Y.-Q.; Shi, J.; Guo, W.-X.; Cheng, S.-Q. Combination of Alpha-Fetoprotein and Neutrophil-to-Lymphocyte Ratio to Predict Treatment Response and Survival Outcomes of Patients with Unresectable Hepatocellular Carcinoma Treated with Immune Checkpoint Inhibitors. BMC Cancer 2023, 23, 547. [Google Scholar] [CrossRef]
- Pons, F.; Varela, M.; Llovet, J.M. Staging Systems in Hepatocellular Carcinoma. Hpb 2005, 7, 35–41. [Google Scholar] [CrossRef]
- Vogel, A.; Cervantes, A.; Chau, I.; Daniele, B.; Llovet, J.M.; Meyer, T.; Nault, J.-C.; Neumann, U.; Ricke, J.; Sangro, B. Hepatocellular Carcinoma: ESMO Clinical Practice Guidelines for Diagnosis, Treatment and Follow-Up. Ann. Oncol. 2018, 29, iv238–iv255. [Google Scholar] [CrossRef]
- Tümen, D.; Heumann, P.; Gülow, K.; Demirci, C.-N.; Cosma, L.-S.; Müller, M.; Kandulski, A. Pathogenesis and Current Treatment Strategies of Hepatocellular Carcinoma. Biomedicines 2022, 10, 3202. [Google Scholar] [CrossRef] [PubMed]
- Galle, P.R.; Forner, A.; Llovet, J.M.; Mazzaferro, V.; Piscaglia, F.; Raoul, J.-L.; Schirmacher, P.; Vilgrain, V. EASL Clinical Practice Guidelines: Management of Hepatocellular Carcinoma. J. Hepatol. 2018, 69, 182–236. [Google Scholar] [CrossRef]
- Heimbach, J.K.; Kulik, L.M.; Finn, R.S.; Sirlin, C.B.; Abecassis, M.M.; Roberts, L.R.; Zhu, A.X.; Murad, M.H.; Marrero, J.A. AASLD Guidelines for the Treatment of Hepatocellular Carcinoma. Hepatology 2018, 67, 358–380. [Google Scholar] [CrossRef] [PubMed]
- Raoul, J.-L.; Forner, A.; Bolondi, L.; Cheung, T.T.; Kloeckner, R.; de Baere, T. Updated Use of TACE for Hepatocellular Carcinoma Treatment: How and When to Use It Based on Clinical Evidence. Cancer Treat. Rev. 2019, 72, 28–36. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, C.; Mahler, M.; Soulen, M.C. Liver-Directed Therapy for Hepatocellular Carcinoma. Chin. Clin. Oncol. 2021, 10, 8. [Google Scholar] [CrossRef] [PubMed]
- Calderaro, J.; Ziol, M.; Paradis, V.; Zucman-Rossi, J. Molecular and Histological Correlations in Liver Cancer. J. Hepatol. 2019, 71, 616–630. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.S.; Qin, S.; Ikeda, M.; Galle, P.R.; Ducreux, M.; Kim, T.-Y.; Kudo, M.; Breder, V.; Merle, P.; Kaseb, A.O. Atezolizumab plus Bevacizumab in Unresectable Hepatocellular Carcinoma. N. Engl. J. Med. 2020, 382, 1894–1905. [Google Scholar] [CrossRef] [PubMed]
- Qin, S.; Jiang, J.; Lu, Y.; Nice, E.C.; Huang, C.; Zhang, J.; He, W. Emerging Role of Tumor Cell Plasticity in Modifying Therapeutic Response. Signal Transduct. Target. Ther. 2020, 5, 228. [Google Scholar] [CrossRef] [PubMed]
- Roehlen, N.; Muller, M.; Nehme, Z.; Crouchet, E.; Jühling, F.; Del Zompo, F.; Cherradi, S.; Duong, F.H.T.; Almeida, N.; Saviano, A.; et al. Treatment of HCC with Claudin-1-Specific Antibodies Suppresses Carcinogenic Signaling and Reprograms the Tumor Microenvironment. J. Hepatol. 2023, 78, 343–355. [Google Scholar] [CrossRef] [PubMed]
- Llovet, J.M.; Kelley, R.K.; Villanueva, A.; Singal, A.G.; Pikarsky, E.; Roayaie, S. Hepatocellular Carcinoma. Nat. Rev. Dis. Primers 2021, 7, 7. [Google Scholar] [CrossRef] [PubMed]
- Prior, N.; Inacio, P.; Huch, M. Liver Organoids: From Basic Research to Therapeutic Applications. Gut 2019, 68, 2228–2237. [Google Scholar] [CrossRef]
- Chang, M.; Bogacheva, M.S.; Lou, Y.-R. Challenges for the Applications of Human Pluripotent Stem Cell-Derived Liver Organoids. Front. Cell Dev. Biol. 2021, 9, 748576. [Google Scholar] [CrossRef]
- Obeid, D.A.; Mir, T.A.; Alzhrani, A.; Altuhami, A.; Shamma, T.; Ahmed, S.; Kazmi, S.; Fujitsuka, I.; Ikhlaq, M.; Shabab, M. Using Liver Organoids as Models to Study the Pathobiology of Rare Liver Diseases. Biomedicines 2024, 12, 446. [Google Scholar] [CrossRef]
- Wilson, H. A New Method by Which Sponges May Be Artificially Reared. Science 1907, 25, 912–915. [Google Scholar] [CrossRef] [PubMed]
- Corrò, C.; Novellasdemunt, L.; Li, V.S. A Brief History of Organoids. Am. J. Physiol.-Cell Physiol. 2020, 319, C151–C165. [Google Scholar] [CrossRef] [PubMed]
- Holtfreter, J. Experimental Studies on the Development of the Pronephros. Rev. Can. Biol. 1943, 3, 220–250. [Google Scholar]
- Weiss, P.; Taylor, A. Reconstitution of Complete Organs from Single-Cell Suspensions of Chick Embryos in Advanced Stages of Differentiation. Proc. Natl. Acad. Sci. USA 1960, 46, 1177–1185. [Google Scholar] [CrossRef] [PubMed]
- Stefanelli, A.; Zacchei, A.; Ceccherini, V. Retinal Reconstitution in Vitro after Disaggregation of Embryonic Chicken Eyes (Translated from Italian). Acta Embryol. Morphol. Exper. 1961, 4, 47–55. [Google Scholar]
- Evans, M. Origin of Mouse Embryonal Carcinoma Cells and the Possibility of Their Direct Isolation into Tissue Culture. Reproduction 1981, 62, 625–631. [Google Scholar] [CrossRef] [PubMed]
- Shannon, J.M.; Mason, R.J.; Jennings, S.D. Functional Differentiation of Alveolar Type II Epithelial Cells in Vitro: Effects of Cell Shape, Cell-Matrix Interactions and Cell-Cell Interactions. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 1987, 931, 143–156. [Google Scholar] [CrossRef]
- Thomson, J.A.; Itskovitz-Eldor, J.; Shapiro, S.S.; Waknitz, M.A.; Swiergiel, J.J.; Marshall, V.S.; Jones, J.M. Embryonic Stem Cell Lines Derived from Human Blastocysts. Science 1998, 282, 1145–1147. [Google Scholar] [CrossRef]
- Takahashi, K.; Yamanaka, S. Induction of Pluripotent Stem Cells from Mouse Embryonic and Adult Fibroblast Cultures by Defined Factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef]
- Eiraku, M.; Watanabe, K.; Matsuo-Takasaki, M.; Kawada, M.; Yonemura, S.; Matsumura, M.; Wataya, T.; Nishiyama, A.; Muguruma, K.; Sasai, Y. Self-Organized Formation of Polarized Cortical Tissues from ESCs and Its Active Manipulation by Extrinsic Signals. Cell Stem Cell 2008, 3, 519–532. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Vries, R.G.; Snippert, H.J.; Van De Wetering, M.; Barker, N.; Stange, D.E.; Van Es, J.H.; Abo, A.; Kujala, P.; Peters, P.J. Single Lgr5 Stem Cells Build Crypt-Villus Structures in Vitro without a Mesenchymal Niche. Nature 2009, 459, 262–265. [Google Scholar] [CrossRef] [PubMed]
- Barker, N.; Huch, M.; Kujala, P.; van de Wetering, M.; Snippert, H.J.; van Es, J.H.; Sato, T.; Stange, D.E.; Begthel, H.; van den Born, M. Lgr5+ ve Stem Cells Drive Self-Renewal in the Stomach and Build Long-Lived Gastric Units In Vitro. Cell Stem Cell 2010, 6, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Eiraku, M.; Takata, N.; Ishibashi, H.; Kawada, M.; Sakakura, E.; Okuda, S.; Sekiguchi, K.; Adachi, T.; Sasai, Y. Self-Organizing Optic-Cup Morphogenesis in Three-Dimensional Culture. Nature 2011, 472, 51–56. [Google Scholar] [CrossRef]
- Nakano, T.; Ando, S.; Takata, N.; Kawada, M.; Muguruma, K.; Sekiguchi, K.; Saito, K.; Yonemura, S.; Eiraku, M.; Sasai, Y. Self-Formation of Optic Cups and Storable Stratified Neural Retina from Human ESCs. Cell Stem Cell 2012, 10, 771–785. [Google Scholar] [CrossRef] [PubMed]
- Takebe, T.; Sekine, K.; Enomura, M.; Koike, H.; Kimura, M.; Ogaeri, T.; Zhang, R.-R.; Ueno, Y.; Zheng, Y.-W.; Koike, N. Vascularized and Functional Human Liver from an iPSC-Derived Organ Bud Transplant. Nature 2013, 499, 481–484. [Google Scholar] [CrossRef]
- Chua, C.W.; Shibata, M.; Lei, M.; Toivanen, R.; Barlow, L.J.; Bergren, S.K.; Badani, K.K.; McKiernan, J.M.; Benson, M.C.; Hibshoosh, H. Single Luminal Epithelial Progenitors Can Generate Prostate Organoids in Culture. Nat. Cell Biol. 2014, 16, 951–961. [Google Scholar] [CrossRef]
- Post, Y.; Puschhof, J.; Beumer, J.; Kerkkamp, H.M.; de Bakker, M.A.; Slagboom, J.; de Barbanson, B.; Wevers, N.R.; Spijkers, X.M.; Olivier, T. Snake Venom Gland Organoids. Cell 2020, 180, 233–247. [Google Scholar] [CrossRef]
- Harrison, S.P.; Siller, R.; Tanaka, Y.; Chollet, M.E.; de la Morena-Barrio, M.E.; Xiang, Y.; Patterson, B.; Andersen, E.; Bravo-Pérez, C.; Kempf, H. Scalable Production of Tissue-like Vascularized Liver Organoids from Human PSCs. Exp. Mol. Med. 2023, 55, 2005–2024. [Google Scholar] [CrossRef]
- Hendriks, D.; Pagliaro, A.; Andreatta, F.; Ma, Z.; van Giessen, J.; Massalini, S.; López-Iglesias, C.; van Son, G.J.; DeMartino, J.; Damen, J.M.A. Human Fetal Brain Self-Organizes into Long-Term Expanding Organoids. Cell 2024, 187, 712–732. [Google Scholar] [CrossRef]
- Maimets, M.; Rocchi, C.; Bron, R.; Pringle, S.; Kuipers, J.; Giepmans, B.N.; Vries, R.G.; Clevers, H.; De Haan, G.; Van Os, R. Long-Term in Vitro Expansion of Salivary Gland Stem Cells Driven by Wnt Signals. Stem Cell Rep. 2016, 6, 150–162. [Google Scholar] [CrossRef]
- Huch, M.; Gehart, H.; van Boxtel, R.; Hamer, K.; Blokzijl, F.; Verstegen, M.M.A.; Ellis, E.; van Wenum, M.; Fuchs, S.A.; de Ligt, J.; et al. Long-Term Culture of Genome-Stable Bipotent Stem Cells from Adult Human Liver. Cell 2015, 160, 299–312. [Google Scholar] [CrossRef]
- Liu, J.; Li, P.; Wang, L.; Li, M.; Ge, Z.; Noordam, L.; Lieshout, R.; Verstegen, M.M.A.; Ma, B.; Su, J.; et al. Cancer-Associated Fibroblasts Provide a Stromal Niche for Liver Cancer Organoids That Confers Trophic Effects and Therapy Resistance. Cell. Mol. Gastroenterol. Hepatol. 2021, 11, 407–431. [Google Scholar] [CrossRef] [PubMed]
- Koike, H.; Iwasawa, K.; Ouchi, R.; Maezawa, M.; Giesbrecht, K.; Saiki, N.; Ferguson, A.; Kimura, M.; Thompson, W.L.; Wells, J.M. Modelling Human Hepato-Biliary-Pancreatic Organogenesis from the Foregut–Midgut Boundary. Nature 2019, 574, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Takebe, T.; Sekine, K.; Kimura, M.; Yoshizawa, E.; Ayano, S.; Koido, M.; Funayama, S.; Nakanishi, N.; Hisai, T.; Kobayashi, T. Massive and Reproducible Production of Liver Buds Entirely from Human Pluripotent Stem Cells. Cell Rep. 2017, 21, 2661–2670. [Google Scholar] [CrossRef]
- Harrison, S.P.; Baumgarten, S.F.; Verma, R.; Lunov, O.; Dejneka, A.; Sullivan, G.J. Liver Organoids: Recent Developments, Limitations and Potential. Front. Med. 2021, 8, 574047. [Google Scholar] [CrossRef]
- Takeishi, K.; de l’Hortet, A.C.; Wang, Y.; Handa, K.; Guzman-Lepe, J.; Matsubara, K.; Morita, K.; Jang, S.; Haep, N.; Florentino, R.M. Assembly and Function of a Bioengineered Human Liver for Transplantation Generated Solely from Induced Pluripotent Stem Cells. Cell Rep. 2020, 31, 107711. [Google Scholar] [CrossRef] [PubMed]
- Velazquez, J.J.; LeGraw, R.; Moghadam, F.; Tan, Y.; Kilbourne, J.; Maggiore, J.C.; Hislop, J.; Liu, S.; Cats, D.; de Sousa Lopes, S.M.C. Gene Regulatory Network Analysis and Engineering Directs Development and Vascularization of Multilineage Human Liver Organoids. Cell Syst. 2021, 12, 41–55. [Google Scholar] [CrossRef]
- Zhu, X.; Zhang, B.; He, Y.; Bao, J. Liver Organoids: Formation Strategies and Biomedical Applications. Tissue Eng. Regen. Med. 2021, 18, 573–585. [Google Scholar] [CrossRef]
- Liu, Q.; Zeng, A.; Liu, Z.; Wu, C.; Song, L. Liver Organoids: From Fabrication to Application in Liver Diseases. Front. Physiol. 2022, 13, 956244. [Google Scholar] [CrossRef]
- Rossant, J.; Tam, P.P. Early Human Embryonic Development: Blastocyst Formation to Gastrulation. Dev. Cell 2022, 57, 152–165. [Google Scholar] [CrossRef] [PubMed]
- Vogt, N. Human Embryogenesis in a Dish. Nat. Methods 2020, 17, 125. [Google Scholar] [CrossRef] [PubMed]
- Fan, T.; Huang, Y. Accessible Chromatin Reveals Regulatory Mechanisms Underlying Cell Fate Decisions during Early Embryogenesis. Sci. Rep. 2021, 11, 7896. [Google Scholar] [CrossRef] [PubMed]
- Shahbazi, M.N. Mechanisms of Human Embryo Development: From Cell Fate to Tissue Shape and Back. Development 2020, 147, dev190629. [Google Scholar] [CrossRef]
- Huch, M.; Dorrell, C.; Boj, S.F.; Van Es, J.H.; Li, V.S.; Van De Wetering, M.; Sato, T.; Hamer, K.; Sasaki, N.; Finegold, M.J. In Vitro Expansion of Single Lgr5+ Liver Stem Cells Induced by Wnt-Driven Regeneration. Nature 2013, 494, 247–250. [Google Scholar] [CrossRef] [PubMed]
- Kruitwagen, H.S.; Oosterhoff, L.A.; Vernooij, I.G.; Schrall, I.M.; van Wolferen, M.E.; Bannink, F.; Roesch, C.; van Uden, L.; Molenaar, M.R.; Helms, J.B. Long-Term Adult Feline Liver Organoid Cultures for Disease Modeling of Hepatic Steatosis. Stem Cell Rep. 2017, 8, 822–830. [Google Scholar] [CrossRef]
- Schneeberger, K.; Sánchez-Romero, N.; Ye, S.; van Steenbeek, F.G.; Oosterhoff, L.A.; Pla Palacin, I.; Chen, C.; van Wolferen, M.E.; van Tienderen, G.; Lieshout, R. Large-scale Production of LGR5-positive Bipotential Human Liver Stem Cells. Hepatology 2020, 72, 257–270. [Google Scholar] [CrossRef]
- Si-Tayeb, K.; Noto, F.K.; Nagaoka, M.; Li, J.; Battle, M.A.; Duris, C.; North, P.E.; Dalton, S.; Duncan, S.A. Highly Efficient Generation of Human Hepatocyte–like Cells from Induced Pluripotent Stem Cells. Hepatology 2010, 51, 297–305. [Google Scholar] [CrossRef]
- Yusa, K.; Rashid, S.T.; Strick-Marchand, H.; Varela, I.; Liu, P.-Q.; Paschon, D.E.; Miranda, E.; Ordóñez, A.; Hannan, N.R.; Rouhani, F.J. Targeted Gene Correction of A1-Antitrypsin Deficiency in Induced Pluripotent Stem Cells. Nature 2011, 478, 391–394. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Wu, J.; Qiu, W.-L.; Yang, L.; Deng, X.; Zhou, Y.; Chen, Y.; Li, X.; Yu, L.; Li, H. Large-Scale Generation of Functional and Transplantable Hepatocytes and Cholangiocytes from Human Endoderm Stem Cells. Cell Rep. 2020, 33, 108455. [Google Scholar] [CrossRef]
- Chen, X.; Jiang, W.; Ahmed, A.; Mahon, C.S.; Müllner, M.; Cao, B.; Xia, T. Engineering Protective Polymer Coatings for Liver Microtissues. Chem. Res. Toxicol. 2018, 32, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, A.; Moll, M.; Gottwald, E.; Nies, C.; Zantl, R.; Wagner, H.; Burkhardt, B.; Sánchez, J.J.M.; Ladurner, R.; Thasler, W. 3D Cultivation Techniques for Primary Human Hepatocytes. Microarrays 2015, 4, 64–83. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Hunziker, W.; Choudhury, D. Engineering Microfluidic Organoid-on-a-Chip Platforms. Micromachines 2019, 10, 165. [Google Scholar] [CrossRef]
- Zaret, K.S. Genetic Programming of Liver and Pancreas Progenitors: Lessons for Stem-Cell Differentiation. Nat. Rev. Genet. 2008, 9, 329–340. [Google Scholar] [CrossRef] [PubMed]
- Broughton, J.P.; Deng, X.; Yu, G.; Fasching, C.L.; Servellita, V.; Singh, J.; Miao, X.; Streithorst, J.A.; Granados, A.; Sotomayor-Gonzalez, A.; et al. CRISPR–Cas12-Based Detection of SARS-CoV-2. Nat. Biotechnol. 2020, 38, 870–874. [Google Scholar] [CrossRef] [PubMed]
- Hendriks, D.; Brouwers, J.F.; Hamer, K.; Geurts, M.H.; Luciana, L.; Massalini, S.; López-Iglesias, C.; Peters, P.J.; Rodríguez-Colman, M.J.; Chuva de Sousa Lopes, S. Engineered Human Hepatocyte Organoids Enable CRISPR-Based Target Discovery and Drug Screening for Steatosis. Nat. Biotechnol. 2023, 41, 1567–1581. [Google Scholar] [CrossRef]
- Lancaster, M.A.; Huch, M. Disease Modelling in Human Organoids. Dis. Models Mech. 2019, 12, dmm039347. [Google Scholar] [CrossRef] [PubMed]
- Calamaio, S.; Serzanti, M.; Boniotti, J.; Fra, A.; Garrafa, E.; Cominelli, M.; Verardi, R.; Poliani, P.L.; Dotti, S.; Villa, R.; et al. Human iPSC-Derived 3D Hepatic Organoids in a Miniaturized Dynamic Culture System. Biomedicines 2023, 11, 2114. [Google Scholar] [CrossRef]
- Nie, Y.-Z.; Zheng, Y.-W.; Ogawa, M.; Miyagi, E.; Taniguchi, H. Human Liver Organoids Generated with Single Donor-Derived Multiple Cells Rescue Mice from Acute Liver Failure. Stem Cell Res. Ther. 2018, 9, 5. [Google Scholar] [CrossRef]
- Kim, S.K.; Kim, Y.H.; Park, S.; Cho, S.-W. Organoid Engineering with Microfluidics and Biomaterials for Liver, Lung Disease, and Cancer Modeling. Acta Biomater. 2021, 132, 37–51. [Google Scholar] [CrossRef]
- Ye, S.; Boeter, J.W.; Penning, L.C.; Spee, B.; Schneeberger, K. Hydrogels for Liver Tissue Engineering. Bioengineering 2019, 6, 59. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Alkhawaji, A.; Ding, Y.; Mei, J. Decellularized Scaffolds in Regenerative Medicine. Oncotarget 2016, 7, 58671. [Google Scholar] [CrossRef] [PubMed]
- Mir, T.A.; Alzhrani, A.; Nakamura, M.; Iwanaga, S.; Wani, S.I.; Altuhami, A.; Kazmi, S.; Arai, K.; Shamma, T.; Obeid, D.A. Whole Liver Derived Acellular Extracellular Matrix for Bioengineering of Liver Constructs: An Updated Review. Bioengineering 2023, 10, 1126. [Google Scholar] [CrossRef] [PubMed]
- Mir, T.A.; Nakamura, M.; Sakai, S.; Iwanaga, S.; Wani, S.I.; Alzhrani, A.; Arai, K.; Mir, B.A.; Kazmi, S.; Assiri, A.M. Mammalian-Specific Decellularized Matrices Derived Bioink for Bioengineering of Liver Tissue Analogues: A Review. Int. J. Bioprinting 2023, 9, 714. [Google Scholar] [CrossRef] [PubMed]
- Ramli, M.N.B.; Lim, Y.S.; Koe, C.T.; Demircioglu, D.; Tng, W.; Gonzales, K.A.U.; Tan, C.P.; Szczerbinska, I.; Liang, H.; Soe, E.L. Human Pluripotent Stem Cell-Derived Organoids as Models of Liver Disease. Gastroenterology 2020, 159, 1471–1486. [Google Scholar] [CrossRef] [PubMed]
- Poling, H.M.; Wu, D.; Brown, N.; Baker, M.; Hausfeld, T.A.; Huynh, N.; Chaffron, S.; Dunn, J.C.; Hogan, S.P.; Wells, J.M. Mechanically Induced Development and Maturation of Human Intestinal Organoids in Vivo. Nat. Biomed. Eng. 2018, 2, 429–442. [Google Scholar] [CrossRef] [PubMed]
- Hoang, P.; Ma, Z. Biomaterial-Guided Stem Cell Organoid Engineering for Modeling Development and Diseases. Acta Biomater. 2021, 132, 23–36. [Google Scholar] [CrossRef]
- Magno, V.; Meinhardt, A.; Werner, C. Polymer Hydrogels to Guide Organotypic and Organoid Cultures. Adv. Funct. Mater. 2020, 30, 2000097. [Google Scholar] [CrossRef]
- Jian, H.; Li, X.; Dong, Q.; Tian, S.; Bai, S. In Vitro Construction of Liver Organoids with Biomimetic Lobule Structure by a Multicellular 3D Bioprinting Strategy. Cell Prolif. 2023, 56, e13465. [Google Scholar] [CrossRef]
- Broguiere, N.; Isenmann, L.; Hirt, C.; Ringel, T.; Placzek, S.; Cavalli, E.; Ringnalda, F.; Villiger, L.; Züllig, R.; Lehmann, R. Growth of Epithelial Organoids in a Defined Hydrogel. Adv. Mater. 2018, 30, 1801621. [Google Scholar] [CrossRef]
- Ye, S.; Boeter, J.W.; Mihajlovic, M.; van Steenbeek, F.G.; van Wolferen, M.E.; Oosterhoff, L.A.; Marsee, A.; Caiazzo, M.; van der Laan, L.J.; Penning, L.C. A Chemically Defined Hydrogel for Human Liver Organoid Culture. Adv. Funct. Mater. 2020, 30, 2000893. [Google Scholar] [CrossRef] [PubMed]
- Bragheri, F.; Martinez Vazquez, R.; Osellame, R. Chapter 12.3—Microfluidics. In Three-Dimensional Microfabrication Using Two-photon Polymerization; Baldacchini, T., Ed.; William Andrew Publishing: Oxford, UK, 2016; pp. 310–334. ISBN 978-0-323-35321-2. [Google Scholar]
- Hofer, M.; Lutolf, M.P. Engineering Organoids. Nat. Rev. Mater. 2021, 6, 402–420. [Google Scholar] [CrossRef] [PubMed]
- Rennert, K.; Steinborn, S.; Gröger, M.; Ungerböck, B.; Jank, A.-M.; Ehgartner, J.; Nietzsche, S.; Dinger, J.; Kiehntopf, M.; Funke, H. A Microfluidically Perfused Three Dimensional Human Liver Model. Biomaterials 2015, 71, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Banaeiyan, A.A.; Theobald, J.; Paukštyte, J.; Wölfl, S.; Adiels, C.B.; Goksör, M. Design and Fabrication of a Scalable Liver-Lobule-on-a-Chip Microphysiological Platform. Biofabrication 2017, 9, 015014. [Google Scholar] [CrossRef] [PubMed]
- Prodanov, L.; Jindal, R.; Bale, S.S.; Hegde, M.; McCarty, W.J.; Golberg, I.; Bhushan, A.; Yarmush, M.L.; Usta, O.B. Long-term Maintenance of a Microfluidic 3D Human Liver Sinusoid. Biotechnol. Bioeng. 2016, 113, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Au, S.H.; Chamberlain, M.D.; Mahesh, S.; Sefton, M.V.; Wheeler, A.R. Hepatic Organoids for Microfluidic Drug Screening. Lab Chip 2014, 14, 3290–3299. [Google Scholar] [CrossRef] [PubMed]
- Esch, M.B.; Mahler, G.J.; Stokol, T.; Shuler, M.L. Body-on-a-Chip Simulation with Gastrointestinal Tract and Liver Tissues Suggests That Ingested Nanoparticles Have the Potential to Cause Liver Injury. Lab Chip 2014, 14, 3081–3092. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Han, H.; Wang, L.; Wang, W.; Yang, M.; Qin, Y. Overcoming the Therapeutic Resistance of Hepatomas by Targeting the Tumor Microenvironment. Front. Oncol. 2022, 12, 988956. [Google Scholar] [CrossRef]
- Gong, L.; Giacomini, M.M.; Giacomini, C.; Maitland, M.L.; Altman, R.B.; Klein, T.E. PharmGKB Summary: Sorafenib Pathways. Pharmacogenet. Genom. 2017, 27, 240–246. [Google Scholar] [CrossRef]
- Della Corte, C.M.; Viscardi, G.; Papaccio, F.; Esposito, G.; Martini, G.; Ciardiello, D.; Martinelli, E.; Ciardiello, F.; Morgillo, F. Implication of the Hedgehog Pathway in Hepatocellular Carcinoma. World J. Gastroenterol. 2017, 23, 4330–4340. [Google Scholar] [CrossRef]
- Rani, B.; Malfettone, A.; Dituri, F.; Soukupova, J.; Lupo, L.; Mancarella, S.; Fabregat, I.; Giannelli, G. Galunisertib Suppresses the Staminal Phenotype in Hepatocellular Carcinoma by Modulating CD44 Expression. Cell Death Dis. 2018, 9, 373. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wang, Y.; Xun, X.; Zhang, C.; Xiang, X.; Cheng, Q.; Hu, S.; Li, Z.; Zhu, J. Hedgehog Signaling Promotes Sorafenib Resistance in Hepatocellular Carcinoma Patient-Derived Organoids. J. Exp. Clin. Cancer Res. 2020, 39, 22. [Google Scholar] [CrossRef]
- Martin, S.P.; Fako, V.; Dang, H.; Dominguez, D.A.; Khatib, S.; Ma, L.; Wang, H.; Zheng, W.; Wang, X.W. PKM2 Inhibition May Reverse Therapeutic Resistance to Transarterial Chemoembolization in Hepatocellular Carcinoma. J. Exp. Clin. Cancer Res. 2020, 39, 99. [Google Scholar] [CrossRef]
- Nuciforo, S.; Fofana, I.; Matter, M.S.; Blumer, T.; Calabrese, D.; Boldanova, T.; Piscuoglio, S.; Wieland, S.; Ringnalda, F.; Schwank, G. Organoid Models of Human Liver Cancers Derived from Tumor Needle Biopsies. Cell Rep. 2018, 24, 1363–1376. [Google Scholar] [CrossRef]
- Chong, Y.C.; Toh, T.B.; Chan, Z.; Lin, Q.X.X.; Thng, D.K.H.; Hooi, L.; Ding, Z.; Shuen, T.; Toh, H.C.; Dan, Y.Y.; et al. Targeted Inhibition of Purine Metabolism Is Effective in Suppressing Hepatocellular Carcinoma Progression. Hepatol. Commun. 2020, 4, 1362–1381. [Google Scholar] [CrossRef]
- Firestein, R.; Marcinkiewicz, C.; Nie, L.; Chua, H.K.; Velazquez Quesada, I.; Torelli, M.; Sternberg, M.; Gligorijevic, B.; Shenderova, O.; Schirhagl, R.; et al. Pharmacodynamic Studies of Fluorescent Diamond Carriers of Doxorubicin in Liver Cancer Cells and Colorectal Cancer Organoids. Nanotechnol. Sci. Appl. 2021, 14, 139–159. [Google Scholar] [CrossRef] [PubMed]
- Jia, D.; Liu, C.; Zhu, Z.; Cao, Y.; Wen, W.; Hong, Z.; Liu, Y.; Liu, E.; Chen, L.; Chen, C.; et al. Novel Transketolase Inhibitor Oroxylin A Suppresses the Non-Oxidative Pentose Phosphate Pathway and Hepatocellular Carcinoma Tumour Growth in Mice and Patient-Derived Organoids. Clin. Transl. Med. 2022, 12, e1095. [Google Scholar] [CrossRef]
- Lim, J.J.; Hooi, L.; Dan, Y.Y.; Bonney, G.K.; Zhou, L.; Chow, P.K.-H.; Chee, C.E.; Toh, T.B.; Chow, E.K.-H. Rational Drug Combination Design in Patient-Derived Avatars Reveals Effective Inhibition of Hepatocellular Carcinoma with Proteasome and CDK Inhibitors. J. Exp. Clin. Cancer Res. 2022, 41, 249. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, V.; Plunkett, W.; Cortes, J.E. Omacetaxine: A Protein Translation Inhibitor for Treatment of Chronic Myelogenous Leukemia. Clin. Cancer Res. 2014, 20, 1735–1740. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Halpert, G.; Lerner, M.G.; Hu, H.; Dimitrion, P.; Weiss, M.J.; He, J.; Philosophe, B.; Burkhart, R.; Burns, W.R.; et al. Protein Synthesis Inhibitor Omacetaxine Is Effective against Hepatocellular Carcinoma. JCI Insight 2021, 6, e138197. [Google Scholar] [CrossRef]
- Ren, X.; Huang, M.; Weng, W.; Xie, Y.; Wu, Y.; Zhu, S.; Zhang, Y.; Li, D.; Lai, J.; Shen, S. Personalized Drug Screening in Patient-Derived Organoids of Biliary Tract Cancer and Its Clinical Application. Cell Rep. Med. 2023, 4, 101277. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Wang, Y.; Cen, J.; Ma, X.; Cui, L.; Qiu, Z.; Zhang, Z.; Li, H.; Yang, R.-Z.; Wang, C.; et al. Modelling Liver Cancer Initiation with Organoids Derived from Directly Reprogrammed Human Hepatocytes. Nat. Cell Biol. 2019, 21, 1015–1026. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Qian, M.; Chen, I.-H.; Finkelstein, D.; Onar-Thomas, A.; Johnson, M.; Calabrese, C.; Bahrami, A.; López-Terrada, D.H.; Yang, J.J.; et al. Acquisition of Cholangiocarcinoma Traits during Advanced Hepatocellular Carcinoma Development in Mice. Am. J. Pathol. 2018, 188, 656–671. [Google Scholar] [CrossRef] [PubMed]
- Saito, Y.; Nakaoka, T.; Muramatsu, T.; Ojima, H.; Sukeda, A.; Sugiyama, Y.; Uchida, R.; Furukawa, R.; Kitahara, A.; Sato, T.; et al. Induction of Differentiation of Intrahepatic Cholangiocarcinoma Cells to Functional Hepatocytes Using an Organoid Culture System. Sci. Rep. 2018, 8, 2821. [Google Scholar] [CrossRef] [PubMed]
- Porporato, P.E.; Filigheddu, N.; Pedro, J.M.B.-S.; Kroemer, G.; Galluzzi, L. Mitochondrial Metabolism and Cancer. Cell Res. 2018, 28, 265–280. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Wang, L.; Wang, Y.; Zhang, S.; Zhou, G.; Lieshout, R.; Ma, B.; Liu, J.; Qu, C.; Verstegen, M.M.A.; et al. Mitochondrial Fusion Via OPA1 and MFN1 Supports Liver Tumor Cell Metabolism and Growth. Cells 2020, 9, 121. [Google Scholar] [CrossRef]
- Yoshikawa, N.; Saito, Y.; Manabe, H.; Nakaoka, T.; Uchida, R.; Furukawa, R.; Muramatsu, T.; Sugiyama, Y.; Kimura, M.; Saito, H. Glucose Depletion Enhances the Stem Cell Phenotype and Gemcitabine Resistance of Cholangiocarcinoma Organoids through AKT Phosphorylation and Reactive Oxygen Species. Cancers 2019, 11, 1993. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Guo, H. Hepatitis B Virus cccDNA: Formation, Regulation and Therapeutic Potential. Antivir. Res. 2020, 180, 104824. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Li, Y.; Wang, Y.; Liu, J.; Lavrijsen, M.; Li, Y.; Zhang, R.; Verstegen, M.M.A.; Wang, Y.; Li, T.-C.; et al. Recapitulating Hepatitis E Virus-Host Interactions and Facilitating Antiviral Drug Discovery in Human Liver-Derived Organoids. Sci. Adv. 2022, 8, eabj5908. [Google Scholar] [CrossRef]
- Clark, M.P.; Huynh, T.; Rao, S.; Mackiewicz, L.; Mason, H.; Romal, S.; Stutz, M.D.; Ahn, S.H.; Earnest, L.; Sozzi, V.; et al. Clinical Stage Drugs Targeting Inhibitor of Apoptosis Proteins Purge Episomal Hepatitis B Viral Genome in Preclinical Models. Cell Death Dis. 2021, 12, 641. [Google Scholar] [CrossRef]
- Baktash, Y.; Madhav, A.; Coller, K.E.; Randall, G. Single Particle Imaging of Polarized Hepatoma Organoids upon Hepatitis C Virus Infection Reveals an Ordered and Sequential Entry Process. Cell Host Microbe 2018, 23, 382–394.e5. [Google Scholar] [CrossRef] [PubMed]
- Collett, S.; Torresi, J.; Silveira, L.E.; Truong, V.K.; Christiansen, D.; Tran, B.M.; Vincan, E.; Ramsland, P.A.; Elbourne, A. Investigating Virus-Host Cell Interactions: Comparative Binding Forces between Hepatitis C Virus-like Particles and Host Cell Receptors in 2D and 3D Cell Culture Models. J. Colloid Interface Sci. 2021, 592, 371–384. [Google Scholar] [CrossRef]
- Yu, J.H.; Ma, S. Organoids as Research Models for Hepatocellular Carcinoma. Exp. Cell Res. 2022, 411, 112987. [Google Scholar] [CrossRef] [PubMed]
- Van de Wetering, M.; Francies, H.E.; Francis, J.M.; Bounova, G.; Iorio, F.; Pronk, A.; van Houdt, W.; van Gorp, J.; Taylor-Weiner, A.; Kester, L. Prospective Derivation of a Living Organoid Biobank of Colorectal Cancer Patients. Cell 2015, 161, 933–945. [Google Scholar] [CrossRef]
- Boj, S.F.; Hwang, C.-I.; Baker, L.A.; Chio, I.I.C.; Engle, D.D.; Corbo, V.; Jager, M.; Ponz-Sarvise, M.; Tiriac, H.; Spector, M.S. Organoid Models of Human and Mouse Ductal Pancreatic Cancer. Cell 2015, 160, 324–338. [Google Scholar] [CrossRef]
- Broutier, L.; Mastrogiovanni, G.; Verstegen, M.M.; Francies, H.E.; Gavarró, L.M.; Bradshaw, C.R.; Allen, G.E.; Arnes-Benito, R.; Sidorova, O.; Gaspersz, M.P. Human Primary Liver Cancer–Derived Organoid Cultures for Disease Modeling and Drug Screening. Nat. Med. 2017, 23, 1424–1435. [Google Scholar] [CrossRef]
- Dong, R.; Zhang, B.; Zhang, X. Liver Organoids: An in Vitro 3D Model for Liver Cancer Study. Cell Biosci. 2022, 12, 1–16. [Google Scholar] [CrossRef]
- Sullivan, S.; Stacey, G.N.; Akazawa, C.; Aoyama, N.; Baptista, R.; Bedford, P.; Bennaceur Griscelli, A.; Chandra, A.; Elwood, N.; Girard, M. Quality Control Guidelines for Clinical-Grade Human Induced Pluripotent Stem Cell Lines. Regen. Med. 2018, 13, 859–866. [Google Scholar] [CrossRef]
- Yoshihara, M.; Hayashizaki, Y.; Murakawa, Y. Genomic Instability of iPSCs: Challenges towards Their Clinical Applications. Stem Cell Rev. Rep. 2017, 13, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Doi, A.; Wen, B.; Ng, K.; Zhao, R.; Cahan, P.; Kim, J.; Aryee, M.; Ji, H.; Ehrlich, L. Epigenetic Memory in Induced Pluripotent Stem Cells. Nature 2010, 467, 285–290. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jabri, A.; Khan, J.; Taftafa, B.; Alsharif, M.; Mhannayeh, A.; Chinnappan, R.; Alzhrani, A.; Kazmi, S.; Mir, M.S.; Alsaud, A.W.; et al. Bioengineered Organoids Offer New Possibilities for Liver Cancer Studies: A Review of Key Milestones and Challenges. Bioengineering 2024, 11, 346. https://doi.org/10.3390/bioengineering11040346
Jabri A, Khan J, Taftafa B, Alsharif M, Mhannayeh A, Chinnappan R, Alzhrani A, Kazmi S, Mir MS, Alsaud AW, et al. Bioengineered Organoids Offer New Possibilities for Liver Cancer Studies: A Review of Key Milestones and Challenges. Bioengineering. 2024; 11(4):346. https://doi.org/10.3390/bioengineering11040346
Chicago/Turabian StyleJabri, Abdullah, Jibran Khan, Bader Taftafa, Mohamed Alsharif, Abdulaziz Mhannayeh, Raja Chinnappan, Alaa Alzhrani, Shadab Kazmi, Mohammad Shabab Mir, Aljohara Waleed Alsaud, and et al. 2024. "Bioengineered Organoids Offer New Possibilities for Liver Cancer Studies: A Review of Key Milestones and Challenges" Bioengineering 11, no. 4: 346. https://doi.org/10.3390/bioengineering11040346
APA StyleJabri, A., Khan, J., Taftafa, B., Alsharif, M., Mhannayeh, A., Chinnappan, R., Alzhrani, A., Kazmi, S., Mir, M. S., Alsaud, A. W., Yaqinuddin, A., Assiri, A. M., AlKattan, K., Vashist, Y. K., Broering, D. C., & Mir, T. A. (2024). Bioengineered Organoids Offer New Possibilities for Liver Cancer Studies: A Review of Key Milestones and Challenges. Bioengineering, 11(4), 346. https://doi.org/10.3390/bioengineering11040346