
N-Acetylglucosamine Sensing and Metabolic Engineering for Attenuating Human and Plant Pathogens

 ,
,  ,
,  and
and
Abstract
:1. Introduction
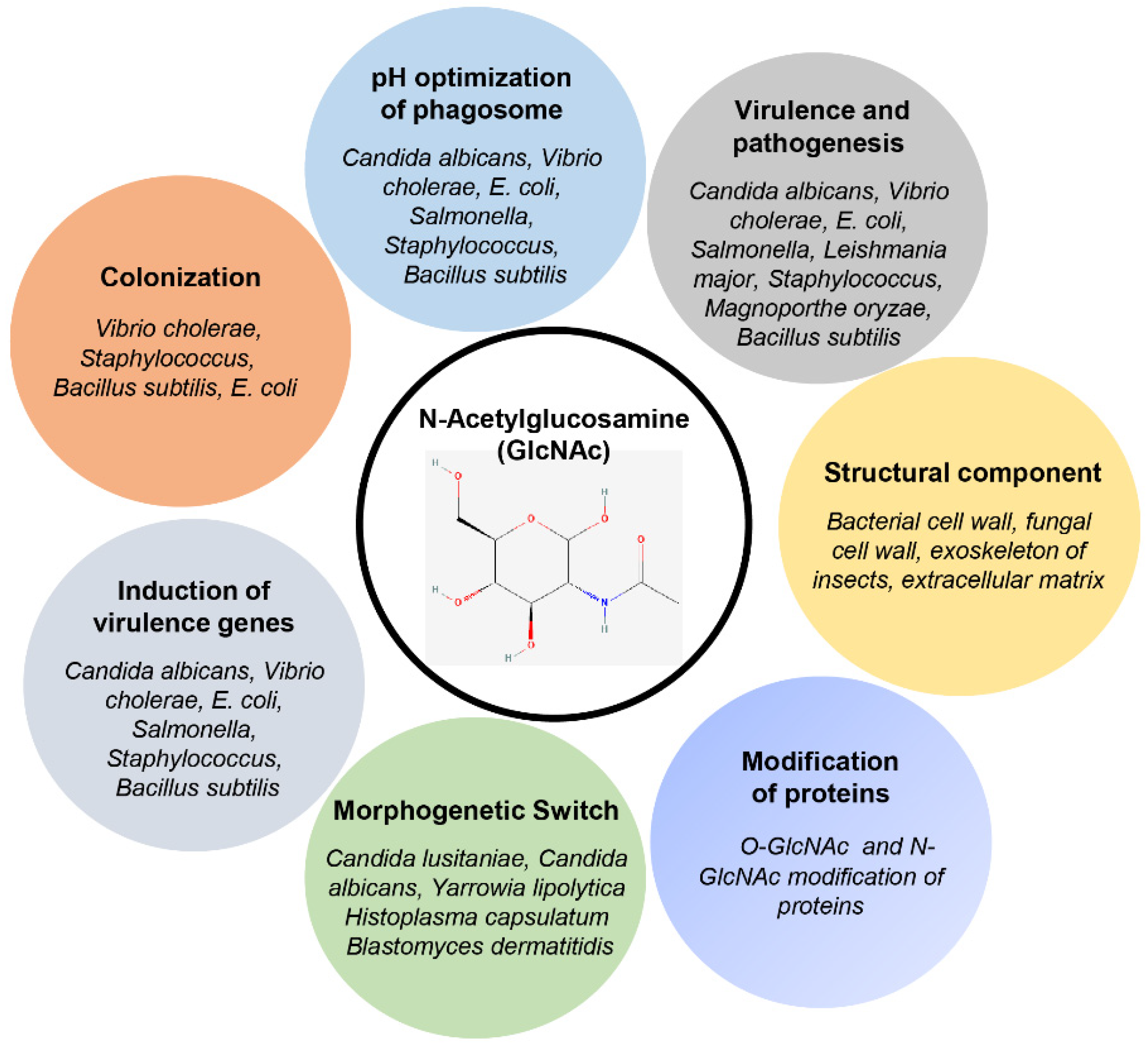
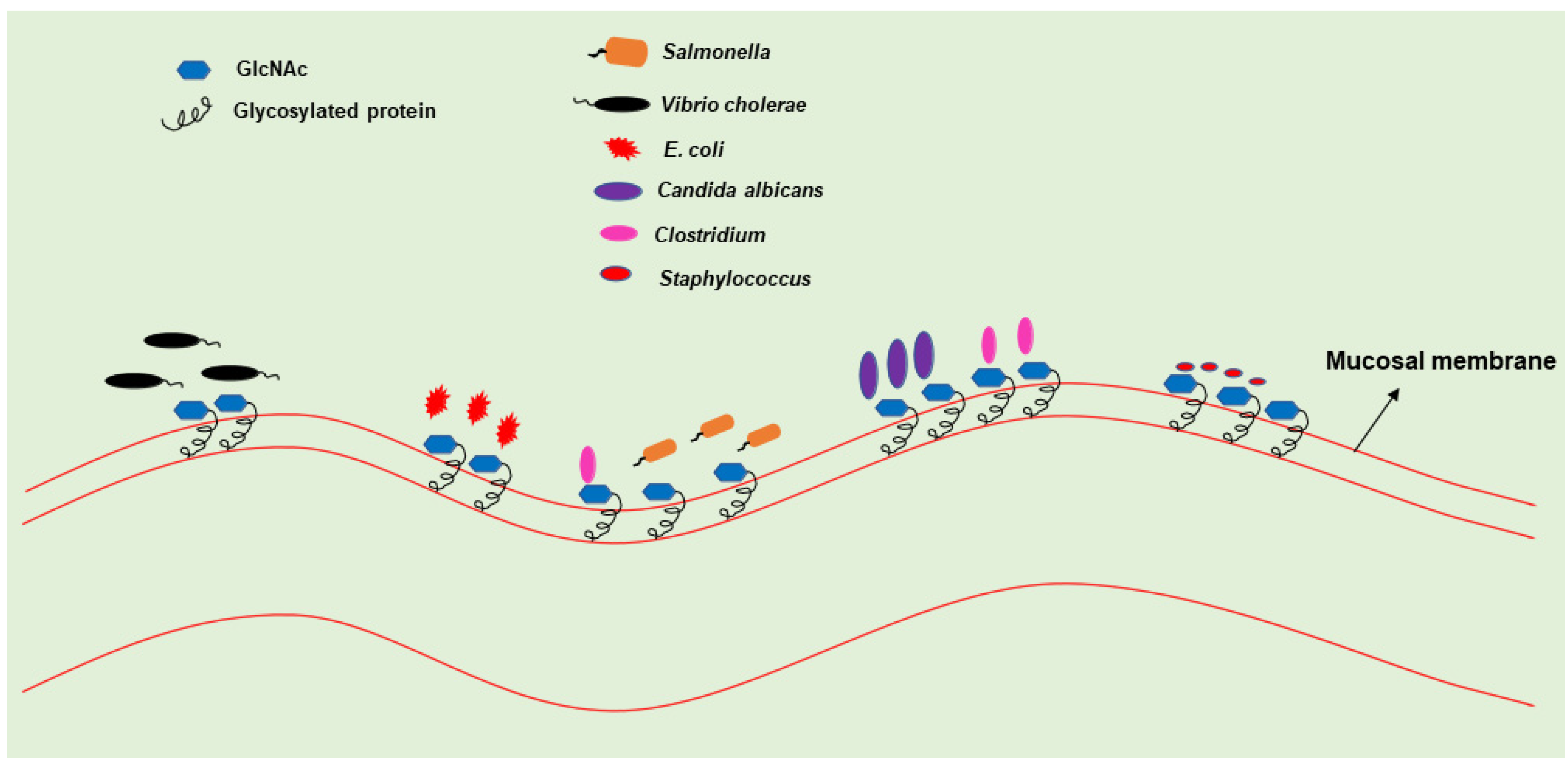
2. GlcNAc: A Ubiquitous Amino Sugar and Signaling Molecule
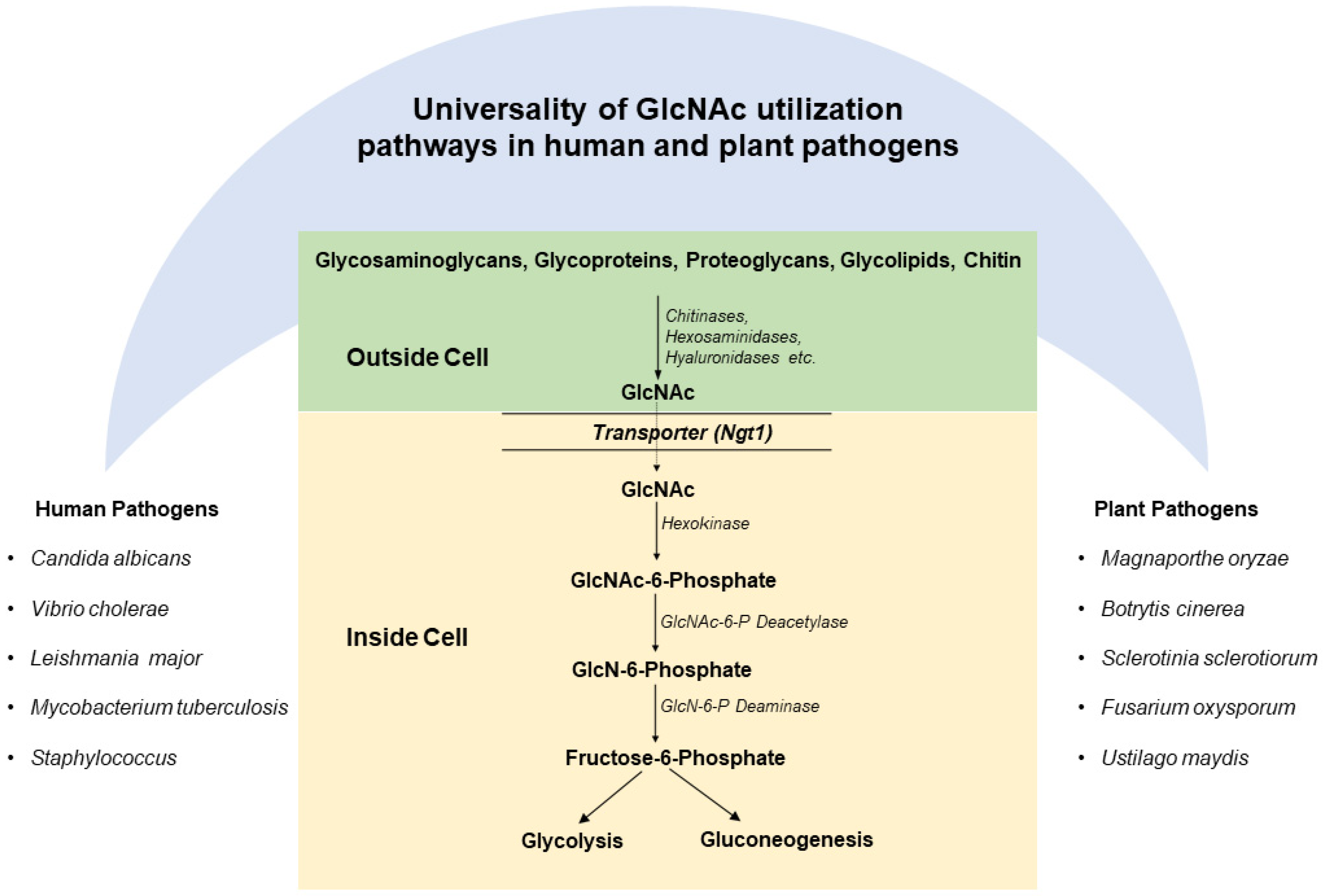
3. Universality of GlcNAc Catabolic Pathway and Genes in Human and Plant Pathogens
4. Engineering GlcNAc Catabolic Pathway for Reducing the Virulence of Human Pathogens
5. GlcNAc Metabolic Engineering for Reducing the Virulence of Plant Pathogens
6. Conclusions and Future Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Avr9 | Avirulence gene |
| Camp | Cyclic adenosine monophosphate |
| CBLs | Chitin-binding lectins |
| CERK1 | Chitin elicitor receptor kinase 1 |
| CRISPR | Clustered regularly interspaced short palindromic repeats |
| DAC | N-acetyl glucosamine-6-phosphate deacetylase |
| DAMPs | Damage-associated molecular patterns |
| ECM | Extracellular Matrix |
| GI | Gastrointestinal |
| GlcNAc-1-P | N-acetylglucosamine-1-phosphate |
| GlcNAc-6-P | N-acetylglucosamine-6-phosphate |
| GlcNAc | N-acetylglucosamine |
| HXK | Hexokinase |
| LYK | LysM domain receptor-like kinase |
| lyk3 | LysM domain receptor-like kinase 3 |
| LysM | Lysin motif domain |
| MAMPs | Microbe-associated molecular patterns |
| NAG | glucosamine-6-phosphate deaminase |
| NFP | Nod factor perception |
| PRR | Pattern recognition receptor |
| PTS | Phosphotransferase system |
| PV | Parasitophorous vacuole |
| ROS | Reactive oxygen species |
| UDP-GlcNAc | Uridine diphosphate N-acetylglucosamine |
References
- Ansari, S.; Bhatt, D.N.; Sood, C.; Datta, A. Functional Characterization of the LdNAGD Gene in Leishmania Donovani. Microbiol. Res. 2021, 251, 126830. [Google Scholar] [CrossRef] [PubMed]
- Naderer, T.; Heng, J.; McConville, M.J. Evidence That Intracellular Stages of Leishmania Major Utilize Amino Sugars as a Major Carbon Source. PLoS Pathog. 2010, 6, e1001245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horta, M.F.; Mendes, B.P.; Roma, E.H.; Noronha, F.S.M.; Macêdo, J.P.; Oliveira, L.S.; Duarte, M.M.; Vieira, L.Q. Reactive Oxygen Species and Nitric Oxide in Cutaneous Leishmaniasis. J. Parasitol. Res. 2012, 2012, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Rao, K.H.; Sengupta, M.; Bhattacharya, S.K.; Datta, A. Two Gene Clusters Co-Ordinate for a Functional N-Acetylglucosamine Catabolic Pathway in Vibrio Cholerae: New Insights into GlcNAc Catabolism of Vibrio Cholerae. Mol. Microbiol. 2011, 80, 1549–1560. [Google Scholar] [CrossRef]
- Swiatek, M.A.; Tenconi, E.; Rigali, S.; van Wezel, G.P. Functional Analysis of the N-Acetylglucosamine Metabolic Genes of Streptomyces Coelicolor and Role in Control of Development and Antibiotic Production. J. Bacteriol. 2012, 194, 1136–1144. [Google Scholar] [CrossRef]
- Miller, K.A.; Phillips, R.S.; Mrazek, J.; Hoover, T.R. Salmonella Utilizes D-Glucosaminate via a Mannose Family Phosphotransferase System Permease and Associated Enzymes. J. Bacteriol. 2013, 195, 4057–4066. [Google Scholar] [CrossRef] [Green Version]
- Ruhela, D.; Kamthan, M.; Saha, P.; Majumdar, S.S.; Datta, K.; Abdin, M.Z.; Datta, A. In Vivo Role of Candida Albicans β-hexosaminidase (HEX 1) in Carbon Scavenging. MicrobiologyOpen 2015, 4, 730–742. [Google Scholar] [CrossRef]
- Konopka, J.B. N-Acetylglucosamine Functions in Cell Signaling. Scientifica 2012, 2012, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Pennisi, E. Armed and Dangerous. Science 2010, 327, 804–805. [Google Scholar] [CrossRef] [Green Version]
- Dean, R.; Van Kan, J.A.L.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 Fungal Pathogens in Molecular Plant Pathology: Top 10 Fungal Pathogens. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doehlemann, G.; Wahl, R.; Horst, R.J.; Voll, L.M.; Usadel, B.; Poree, F.; Stitt, M.; Pons-Kühnemann, J.; Sonnewald, U.; Kahmann, R.; et al. Reprogramming a Maize Plant: Transcriptional and Metabolic Changes Induced by the Fungal Biotroph. Ustilago Maydis: Plant Response to U. Maydis Infection. Plant J. 2008, 56, 181–195. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Chattopadhyay, A.; Ghosh, S.; Irfan, M.; Chakraborty, N.; Chakraborty, S.; Datta, A. Improving Nutritional Quality and Fungal Tolerance in Soya Bean and Grass Pea by Expressing an Oxalate Decarboxylase. Plant Biotechnol. J. 2016, 14, 1394–1405. [Google Scholar] [CrossRef] [PubMed]
- Chandra, H.; Kumari, P.; Bisht, R.; Prasad, R.; Yadav, S. Plant Growth Promoting Pseudomonas Aeruginosa from Valeriana Wallichii Displays Antagonistic Potential against Three Phytopathogenic Fungi. Mol. Biol. Rep. 2020, 47, 6015–6026. [Google Scholar] [CrossRef]
- Chandra, H.; Kumari, P.; Prasad, R.; Chandra Gupta, S.; Yadav, S. Antioxidant and Antimicrobial Activity Displayed by a Fungal Endophyte Alternaria Alternata Isolated from Picrorhiza Kurroa from Garhwal Himalayas, India. Biocatal. Agric. Biotechnol. 2021, 33, 101955. [Google Scholar] [CrossRef]
- van Kan, J.A.L. Licensed to Kill: The Lifestyle of a Necrotrophic Plant Pathogen. Trends Plant Sci. 2006, 11, 247–253. [Google Scholar] [CrossRef]
- Divon, H.H.; Fluhr, R. Nutrition Acquisition Strategies during Fungal Infection of Plants. FEMS Microbiol. Lett. 2007, 266, 65–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, R.A.; Talbot, N.J. Under Pressure: Investigating the Biology of Plant Infection by Magnaporthe Oryzae. Nat. Rev. Microbiol 2009, 7, 185–195. [Google Scholar] [CrossRef]
- Jaiswal, P.; Cheruku, J.R.; Kumar, K.; Yadav, S.; Singh, A.; Kumari, P.; Dube, S.C.; Upadhyaya, K.C.; Verma, P.K. Differential Transcript Accumulation in Chickpea during Early Phases of Compatible Interaction with a Necrotrophic Fungus Ascochyta Rabiei. Mol. Biol. Rep. 2012, 39, 4635–4646. [Google Scholar] [CrossRef]
- Torres, M.A.; Jones, J.D.G.; Dangl, J.L. Reactive Oxygen Species Signaling in Response to Pathogens. Plant Physiol. 2006, 141, 373–378. [Google Scholar] [CrossRef] [Green Version]
- Yadav, S.; Kushwaha, H.R.; Kumar, K.; Verma, P.K. Comparative Structural Modeling of a Monothiol GRX from Chickpea: Insight in Iron–Sulfur Cluster Assembly. Int. J. Biol. Macromol. 2012, 51, 266–273. [Google Scholar] [CrossRef]
- Kliebenstein, D.J.; Rowe, H.C. Ecological Costs of Biotrophic versus Necrotrophic Pathogen Resistance, the Hypersensitive Response and Signal Transduction. Plant Sci. 2008, 174, 551–556. [Google Scholar] [CrossRef]
- Glazebrook, J. Contrasting Mechanisms of Defense Against Biotrophic and Necrotrophic Pathogens. Annu. Rev. Phytopathol. 2005, 43, 205–227. [Google Scholar] [CrossRef] [PubMed]
- Kumari, P.; Gupta, A.; Yadav, S. Thioredoxins as Molecular Players in Plants, Pests, and Pathogens. In Plant-Pest Interactions: From Molecular Mechanisms to Chemical Ecology; Singh, I.K., Singh, A., Eds.; Springer: Singapore, 2021; pp. 107–125. [Google Scholar] [CrossRef]
- Naseem, S.; Konopka, J.B. N-Acetylglucosamine Regulates Virulence Properties in Microbial Pathogens. PLoS Pathog. 2015, 11, e1004947. [Google Scholar] [CrossRef] [PubMed]
- Moussian, B. The Role of GlcNAc in Formation and Function of Extracellular Matrices. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2008, 149, 215–226. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Guan, G.; Li, X.; Gulati, M.; Tao, L.; Cao, C.; Johnson, A.D.; Nobile, C.J.; Huang, G. N-Acetylglucosamine-Induced Cell Death in Candida Albicans and Its Implications for Adaptive Mechanisms of Nutrient Sensing in Yeasts. mBio 2015, 6, e01376-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Datta, A.; Ganesan, K.; Natarajan, K. Current Trends in Candida Albicans Research. In Advances in Microbial Physiology; Elsevier: Amsterdam, The Netherlands, 1990; Volume 30, pp. 53–88. [Google Scholar] [CrossRef]
- Singh, P.; Ghosh, S.; Datta, A. Attenuation of Virulence and Changes in Morphology in Candida Albicans by Disruption of the N-Acetylglucosamine Catabolic Pathway. Infect. Immun. 2001, 69, 7898–7903. [Google Scholar] [CrossRef] [Green Version]
- Olszewski, N.E.; West, C.M.; Sassi, S.O.; Hartweck, L.M. O-GlcNAc Protein Modification in Plants: Evolution and Function. Biochim. Et Biophys. Acta (BBA) Gen. Subj. 2010, 1800, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Meli, V.S.; Ghosh, S.; Prabha, T.N.; Chakraborty, N.; Chakraborty, S.; Datta, A. Enhancement of Fruit Shelf Life by Suppressing N-Glycan Processing Enzymes. Proc. Natl. Acad. Sci. USA 2010, 107, 2413–2418. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, S.; Meli, V.S.; Kumar, A.; Thakur, A.; Chakraborty, N.; Chakraborty, S.; Datta, A. The N-Glycan Processing Enzymes α-Mannosidase and β-D-N-Acetylhexosaminidase Are Involved in Ripening-Associated Softening in the Non-Climacteric Fruits of Capsicum. J. Exp. Bot. 2011, 62, 571–582. [Google Scholar] [CrossRef]
- Irfan, M.; Ghosh, S.; Meli, V.S.; Kumar, A.; Kumar, V.; Chakraborty, N.; Chakraborty, S.; Datta, A. Fruit Ripening Regulation of α-Mannosidase Expression by the MADS Box Transcription Factor RIPENING INHIBITOR and Ethylene. Front. Plant Sci. 2016, 7, 10. [Google Scholar] [CrossRef] [Green Version]
- Irfan, M.; Ghosh, S.; Kumar, V.; Chakraborty, N.; Chakraborty, S.; Datta, A. Insights into Transcriptional Regulation of -D-N-Acetylhexosaminidase, an N-Glycan-Processing Enzyme Involved in Ripening-Associated Fruit Softening. J. Exp. Bot. 2014, 65, 5835–5848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sumit Ghosh, S.G.; Mohammad Irfan, M.I.; Asis Datta, A.D. Application of RNA Silencing in Improving Plant Traits for Industrial Use. In Plant Gene Silencing: Mechanisms and Applications; Dalmay, T., Ed.; CABI: Wallingford, UK, 2017; pp. 128–146. [Google Scholar] [CrossRef]
- Ishikawa, T.; Fang, L.; Rennie, E.A.; Sechet, J.; Yan, J.; Jing, B.; Moore, W.; Cahoon, E.B.; Scheller, H.V.; Kawai-Yamada, M.; et al. GLUCOSAMINE INOSITOLPHOSPHORYLCERAMIDE TRANSFERASE1 (GINT1) Is a GlcNAc-Containing Glycosylinositol Phosphorylceramide Glycosyltransferase. Plant Physiol. 2018, 177, 938–952. [Google Scholar] [CrossRef]
- Skoog, E.C.; Sjöling, Å.; Navabi, N.; Holgersson, J.; Lundin, S.B.; Lindén, S.K. Human Gastric Mucins Differently Regulate Helicobacter Pylori Proliferation, Gene Expression and Interactions with Host Cells. PLoS ONE 2012, 7, e36378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddi, G.; Pruss, K.; Cottingham, K.L.; Taylor, R.K.; Almagro-Moreno, S. Catabolism of Mucus Components Influences Motility of Vibrio Cholerae in the Presence of Environmental Reservoirs. PLoS ONE 2018, 13, e0201383. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Bleumink-Pluym, N.M.C.; Luijkx, Y.M.C.A.; Wubbolts, R.W.; van Putten, J.P.M.; Strijbis, K. MUC1 Is a Receptor for the Salmonella SiiE Adhesin That Enables Apical Invasion into Enterocytes. PLoS Pathog. 2019, 15, e1007566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavalcante, T.; Medeiros, M.M.; Mule, S.N.; Palmisano, G.; Stolf, B.S. The Role of Sialic Acids in the Establishment of Infections by Pathogens, With Special Focus on Leishmania. Front. Cell. Infect. Microbiol. 2021, 11, 671913. [Google Scholar] [CrossRef]
- Kato, K.; Ishiwa, A. The Role of Carbohydrates in Infection Strategies of Enteric Pathogens. Trop. Med. Health 2015, 43, 41–52. [Google Scholar] [CrossRef] [Green Version]
- Azzouz, N.; Shams-Eldin, H.; Niehus, S.; Debierre-Grockiego, F.; Bieker, U.; Schmidt, J.; Mercier, C.; Delauw, M.-F.; Dubremetz, J.-F.; Smith, T.K.; et al. Toxoplasma Gondii Grown in Human Cells Uses GalNAc-Containing Glycosylphosphatidylinositol Precursors to Anchor Surface Antigens While the Immunogenic Glc–GalNAc-Containing Precursors Remain Free at the Parasite Cell Surface. Int. J. Biochem. Cell Biol. 2006, 38, 1914–1925. [Google Scholar] [CrossRef]
- Sheikh, A.; Wangdi, T.; Vickers, T.J.; Aaron, B.; Palmer, M.; Miller, M.J.; Kim, S.; Herring, C.; Simoes, R.; Crainic, J.A.; et al. Enterotoxigenic Escherichia Coli Degrades the Host MUC2 Mucin Barrier to Facilitate Critical Pathogen-Enterocyte Interactions in Human Small Intestine. Infect. Immun. 2021, IAI-00572. [Google Scholar] [CrossRef]
- Van Ende, M.; Wijnants, S.; Van Dijck, P. Sugar Sensing and Signaling in Candida Albicans and Candida Glabrata. Front. Microbiol. 2019, 10, 99. [Google Scholar] [CrossRef]
- Puri, S.; Friedman, J.; Saraswat, D.; Kumar, R.; Li, R.; Ruszaj, D.; Edgerton, M. Candida Albicans Shed Msb2 and Host Mucins Affect the Candidacidal Activity of Salivary Hst 5. Pathogens 2015, 4, 752–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Everard, A.; Belzer, C.; Geurts, L.; Ouwerkerk, J.P.; Druart, C.; Bindels, L.B.; Guiot, Y.; Derrien, M.; Muccioli, G.G.; Delzenne, N.M.; et al. Cross-Talk between Akkermansia Muciniphila and Intestinal Epithelium Controls Diet-Induced Obesity. Proc. Natl. Acad. Sci. USA 2013, 110, 9066–9071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geerlings, S.; Kostopoulos, I.; de Vos, W.; Belzer, C. Akkermansia Muciniphila in the Human Gastrointestinal Tract: When, Where, and How? Microorganisms 2018, 6, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, F.C.; Wasmund, K.; Cobankovic, I.; Jehmlich, N.; Herbold, C.W.; Lee, K.S.; Sziranyi, B.; Vesely, C.; Decker, T.; Stocker, R.; et al. Rational Design of a Microbial Consortium of Mucosal Sugar Utilizers Reduces Clostridiodes Difficile Colonization. Nat. Commun. 2020, 11, 5104. [Google Scholar] [CrossRef] [PubMed]
- Shuter, J.; Hatcher, V.B.; Lowy, F.D. Staphylococcus Aureus Binding to Human Nasal Mucin. Infect. Immun. 1996, 64, 310–318. [Google Scholar] [CrossRef] [Green Version]
- Olson, M.E.; King, J.M.; Yahr, T.L.; Horswill, A.R. Sialic Acid Catabolism in Staphylococcus Aureus. J. Bacteriol. 2013, 195, 1779–1788. [Google Scholar] [CrossRef] [Green Version]
- Kloehn, J.; Blume, M.; Cobbold, S.; Saunders, E.; Dagley, M.; McConville, M. Using Metabolomics to Dissect Host–Parasite Interactions. Curr. Opin. Microbiol. 2016, 32, 59–65. [Google Scholar] [CrossRef]
- Ghosh, A.K.; Sardar, A.H.; Mandal, A.; Saini, S.; Abhishek, K.; Kumar, A.; Purkait, B.; Singh, R.; Das, S.; Mukhopadhyay, R.; et al. Metabolic Reconfiguration of the Central Glucose Metabolism: A Crucial Strategy of Leishmania Donovani for Its Survival during Oxidative Stress. FASEB J. 2015, 29, 2081–2098. [Google Scholar] [CrossRef]
- Kamthan, A.; Kamthan, M.; Datta, A. Biotechnology for Drug Discovery and Crop Improvement. Nucleus 2017, 60, 237–242. [Google Scholar] [CrossRef]
- Biswas, S.; Van Dijck, P.; Datta, A. Environmental Sensing and Signal Transduction Pathways Regulating Morphopathogenic Determinants of Candida Albicans. Microbiol. Mol. Biol. Rev. 2007, 71, 348–376. [Google Scholar] [CrossRef] [Green Version]
- Ghorai, P.; Irfan, M.; Narula, A.; Datta, A. A Comprehensive Analysis of Candida Albicans Phosphoproteome Reveals Dynamic Changes in Phosphoprotein Abundance during Hyphal Morphogenesis. Appl. Microbiol. Biotechnol. 2018, 102, 9731–9743. [Google Scholar] [CrossRef] [PubMed]
- Kamthan, M.; Mukhopadhyay, G.; Chakraborty, N.; Chakraborty, S.; Datta, A. Quantitative Proteomics and Metabolomics Approaches to Demonstrate N-Acetyl-d-Glucosamine Inducible Amino Acid Deprivation Response as Morphological Switch in Candida Albicans. Fungal Genet. Biol. 2012, 49, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Hanumantha Rao, K.; Paul, S.; Ghosh, S. N-Acetylglucosamine Signaling: Transcriptional Dynamics of a Novel Sugar Sensing Cascade in a Model Pathogenic Yeast, Candida Albicans. JoF 2021, 7, 65. [Google Scholar] [CrossRef] [PubMed]
- Vesely, E.M.; Williams, R.B.; Konopka, J.B.; Lorenz, M.C. N-Acetylglucosamine Metabolism Promotes Survival of Candida Albicans in the Phagosome. mSphere 2017, 2, e00357-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamthan, M.; Kamthan, A.; Ruhela, D.; Maiti, P.; Bhavesh, N.S.; Datta, A. Upregulation of Galactose Metabolic Pathway by N-Acetylglucosamine Induced Endogenous Synthesis of Galactose in Candida Albicans. Fungal Genet. Biol. 2013, 54, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Shen, A.; Kamp, H.D.; Gründling, A.; Higgins, D.E. A Bifunctional O-GlcNAc Transferase Governs Flagellar Motility through Anti-Repression. Genes Dev. 2006, 20, 3283–3295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grigorian, A.; Araujo, L.; Naidu, N.N.; Place, D.J.; Choudhury, B.; Demetriou, M. N-Acetylglucosamine Inhibits T-Helper 1 (Th1)/T-Helper 17 (Th17) Cell Responses and Treats Experimental Autoimmune Encephalomyelitis. J. Biol. Chem. 2011, 286, 40133–40141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stokes, M.J.; Güther, M.L.S.; Turnock, D.C.; Prescott, A.R.; Martin, K.L.; Alphey, M.S.; Ferguson, M.A.J. The Synthesis of UDP-N-Acetylglucosamine Is Essential for Bloodstream Form Trypanosoma Brucei in Vitro and in Vivo and UDP-N-Acetylglucosamine Starvation Reveals a Hierarchy in Parasite Protein Glycosylation. J. Biol. Chem. 2008, 283, 16147–16161. [Google Scholar] [CrossRef] [Green Version]
- Soni, V.; Upadhayay, S.; Suryadevara, P.; Samla, G.; Singh, A.; Yogeeswari, P.; Sriram, D.; Nandicoori, V.K. Depletion of M. Tuberculosis GlmU from Infected Murine Lungs Effects the Clearance of the Pathogen. PLoS Pathog. 2015, 11, e1005235. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Xin, Y.; Sha, S.; Ma, Y. Kinetic Properties of Mycobacterium Tuberculosis Bifunctional GlmU. Arch Microbiol. 2011, 193, 751–757. [Google Scholar] [CrossRef]
- Bhatt, D.N.; Ansari, S.; Kumar, A.; Ghosh, S.; Narula, A.; Datta, A. Magnaporthe Oryzae MoNdt80 Is a Transcriptional Regulator of GlcNAc Catabolic Pathway Involved in Pathogenesis. Microbiol. Res. 2020, 239, 126550. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Ghosh, S.; Bhatt, D.N.; Narula, A.; Datta, A. M Agnaporthe Oryzae Aminosugar Metabolism Is Essential for Successful Host Colonization: N-Acetylglucosamine Metabolism by Rice Blast Fungus. Environ. Microbiol. 2016, 18, 1063–1077. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, F.M.; Mendoza-Hernandez, G.; Castañeda-Bueno, M.; Aparicio, R.; Fischer, H.; Calcagno, M.L.; Oliva, G. Structural Analysis of N-Acetylglucosamine-6-Phosphate Deacetylase Apoenzyme from Escherichia Coli. J. Mol. Biol. 2006, 359, 308–321. [Google Scholar] [CrossRef] [PubMed]
- Ishibashi, A.; Saga, K.; Hisatomi, Y.; Li, Y.; Kaneda, Y.; Nimura, K. A Simple Method Using CRISPR-Cas9 to Knock-out Genes in Murine Cancerous Cell Lines. Sci. Rep. 2020, 10, 22345. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, A.; Puri, M.; Datta, A. Induction of N-Acetylglucosamine Kinase in Yeast. Biochem. J. 1974, 141, 593–595. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Datta, A. Induction of N-Acetylglucosamine-Catabolic Pathway in Spheroplasts of Candida Albicans. Biochem. J. 1979, 178, 427–431. [Google Scholar] [CrossRef] [Green Version]
- Kumar, M.J.; Jamaluddin, Md. S.; Natarajan, K.; Kaur, D.; Datta, A. The Inducible N-Acetylglucosamine Catabolic Pathway Gene Cluster in Candida Albicans: Discrete N-Acetylglucosamine-Inducible Factors Interact at the Promoter of NAG1. Proc. Natl. Acad. Sci. USA 2000, 97, 14218–14223. [Google Scholar] [CrossRef] [Green Version]
- Moye, Z.D.; Burne, R.A.; Zeng, L. Uptake and Metabolism of N-Acetylglucosamine and Glucosamine by Streptococcus Mutans. Appl. Environ. Microbiol. 2014, 80, 5053–5067. [Google Scholar] [CrossRef] [Green Version]
- Spoel, S.H.; Dong, X. How Do Plants Achieve Immunity? Defence without Specialized Immune Cells. Nat. Rev. Immunol. 2012, 12, 89–100. [Google Scholar] [CrossRef]
- Choi, H.W.; Klessig, D.F. DAMPs, MAMPs, and NAMPs in Plant Innate Immunity. BMC Plant Biol. 2016, 16, 232. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Zhou, J.-M. Plant Immunity Triggered by Microbial Molecular Signatures. Mol. Plant 2010, 3, 783–793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, J.; Zhang, X.-C.; Stacey, G. Chitin Signaling and Plant Disease Resistance. Plant Signal. Behav. 2008, 3, 831–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desaki, Y.; Miyata, K.; Suzuki, M.; Shibuya, N.; Kaku, H. Plant Immunity and Symbiosis Signaling Mediated by LysM Receptors. Innate Immun. 2018, 24, 92–100. [Google Scholar] [CrossRef] [PubMed]
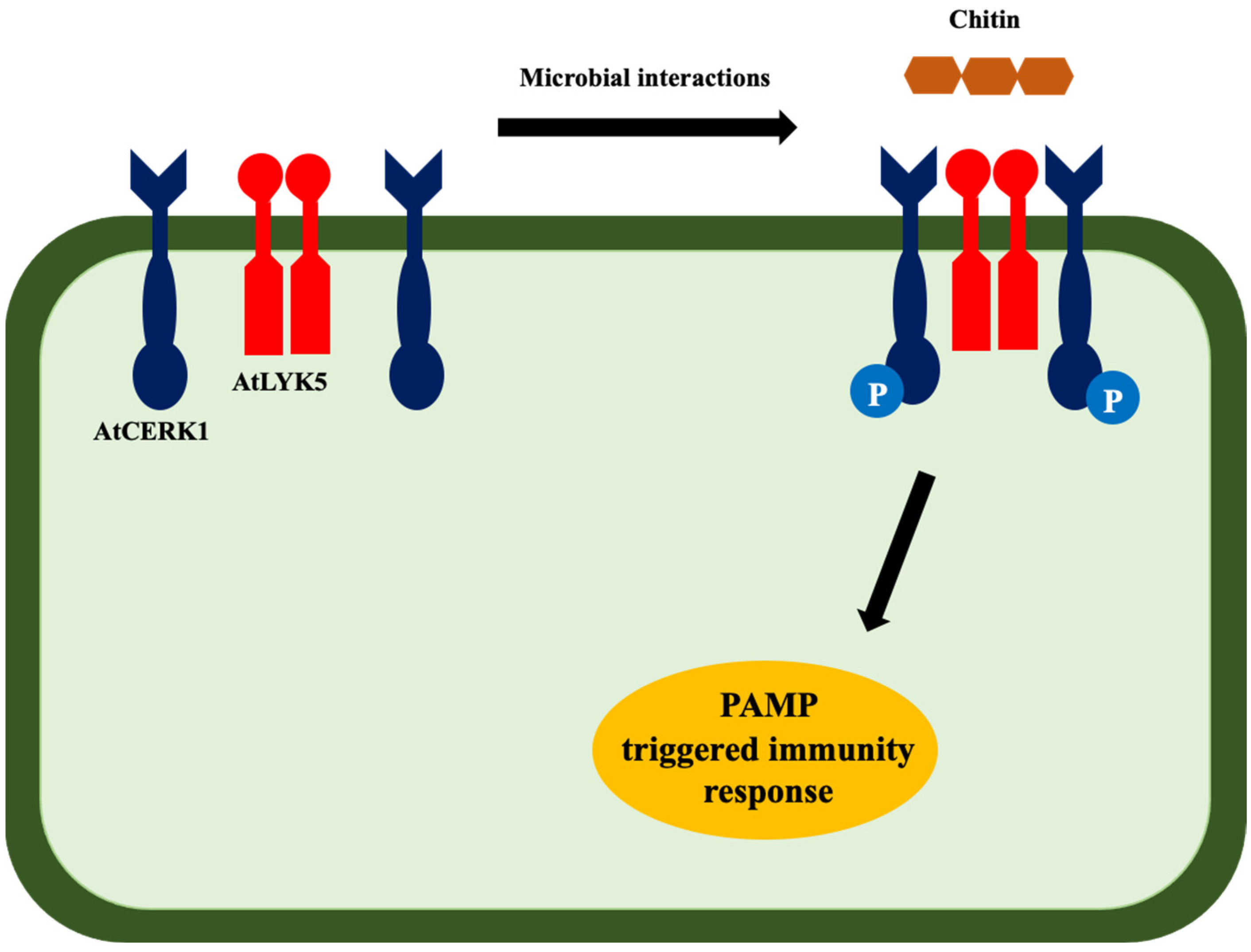
- Huang, C.; Yan, Y.; Zhao, H.; Ye, Y.; Cao, Y. Arabidopsis CPK5 Phosphorylates the Chitin Receptor LYK5 to Regulate Plant Innate Immunity. Front. Plant Sci. 2020, 11, 702. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Liang, Y.; Tanaka, K.; Nguyen, C.T.; Jedrzejczak, R.P.; Joachimiak, A.; Stacey, G. The Kinase LYK5 Is a Major Chitin Receptor in Arabidopsis and Forms a Chitin-Induced Complex with Related Kinase CERK1. eLife 2014, 3, e03766. [Google Scholar] [CrossRef] [PubMed]
- Kashulin, P.A.; Merzlyak, M.N.; Zhiboedov, P.M.; Zhirov, V.K. Extremely Rapid Effects of Polyunsaturated Fatty Acids and N-Acetylglucosamine on Free-Radical Metabolism in Cultured Potato Plant Cells. Biochem. Soc. Trans. 2000, 28, 865–867. [Google Scholar] [CrossRef]
- Raikhel, N.V.; Lee, H.I.; Broekaert, W.F. Structure and Function of Chitin-Binding Proteins. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1993, 44, 591–615. [Google Scholar] [CrossRef]
- Asensio, J.L.; Cañada, F.J.; Siebert, H.C.; Laynez, J.; Poveda, A.; Nieto, P.M.; Soedjanaamadja, U.M.; Gabius, H.J.; Jiménez-Barbero, J. Structural Basis for Chitin Recognition by Defense Proteins: GlcNAc Residues Are Bound in a Multivalent Fashion by Extended Binding Sites in Hevein Domains. Chem. Biol. 2000, 7, 529–543. [Google Scholar] [CrossRef] [Green Version]
- Kooman-Gersmann, M.; Vogelsang, R.; Vossen, P.; van den Hooven, H.W.; Mahé, E.; Honée, G.; de Wit, P.J. Correlation between Binding Affinity and Necrosis-Inducing Activity of Mutant AVR9 Peptide Elicitors. Plant Physiol. 1998, 117, 609–618. [Google Scholar] [CrossRef] [Green Version]
- Kourelis, J.; van der Hoorn, R.A.L. Defended to the Nines: 25 Years of Resistance Gene Cloning Identifies Nine Mechanisms for R Protein Function. Plant Cell 2018, 30, 285–299. [Google Scholar] [CrossRef] [Green Version]
- van den Burg, H.A.; Spronk, C.A.E.M.; Boeren, S.; Kennedy, M.A.; Vissers, J.P.C.; Vuister, G.W.; de Wit, P.J.G.M.; Vervoort, J. Binding of the AVR4 Elicitor of Cladosporium Fulvum to Chitotriose Units Is Facilitated by Positive Allosteric Protein-Protein Interactions: The Chitin-Binding Site of AVR4 Represents a Novel Binding Site on the Folding Scaffold Shared between the Invertebrate and the Plant Chitin-Binding Domain. J. Biol. Chem. 2004, 279, 16786–16796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van den Burg, H.A.; Harrison, S.J.; Joosten, M.H.A.J.; Vervoort, J.; de Wit, P.J.G.M. Cladosporium Fulvum Avr4 Protects Fungal Cell Walls against Hydrolysis by Plant Chitinases Accumulating during Infection. Mol. Plant Microbe Interact. 2006, 19, 1420–1430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Esse, H.P.; Bolton, M.D.; Stergiopoulos, I.; de Wit, P.J.G.M.; Thomma, B.P.H.J. The Chitin-Binding Cladosporium Fulvum Effector Protein Avr4 Is a Virulence Factor. Mol. Plant Microbe Interact. 2007, 20, 1092–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martín-Urdíroz, M.; Roncero, M.I.G.; González-Reyes, J.A.; Ruiz-Roldán, C. ChsVb, a Class VII Chitin Synthase Involved in Septation, Is Critical for Pathogenicity in Fusarium Oxysporum. Eukaryot. Cell 2008, 7, 112–121. [Google Scholar] [CrossRef] [Green Version]
- Madrid, M.P.; Di Pietro, A.; Roncero, M.I.G. Class V Chitin Synthase Determines Pathogenesis in the Vascular Wilt Fungus Fusarium Oxysporum and Mediates Resistance to Plant Defence Compounds. Mol. Microbiol. 2003, 47, 257–266. [Google Scholar] [CrossRef]
- Jaroszuk-Ściseł, J.; Kurek, E. Hydrolysis of Fungal and Plant Cell Walls by Enzymatic Complexes from Cultures of Fusarium Isolates with Different Aggressiveness to Rye (Secale Cereale). Arch Microbiol. 2012, 194, 653–665. [Google Scholar] [CrossRef]
- Jaroszuk-Scisel, J.; Kurek, E.; Slomka, A.; Janczarek, M.; Rodzik, B. Activities of Cell Wall Degrading Enzymes in Autolyzing Cultures of Three Fusarium Culmorum Isolates: Growth-Promoting, Deleterious and Pathogenic to Rye (Secale Cereale). Mycologia 2011, 103, 929–945. [Google Scholar] [CrossRef]
- Rey, T.; Nars, A.; Bonhomme, M.; Bottin, A.; Huguet, S.; Balzergue, S.; Jardinaud, M.-F.; Bono, J.-J.; Cullimore, J.; Dumas, B.; et al. NFP, a LysM Protein Controlling Nod Factor Perception, Also Intervenes in Medicago Truncatula Resistance to Pathogens. New Phytol. 2013, 198, 875–886. [Google Scholar] [CrossRef]
- Mesnage, S.; Dellarole, M.; Baxter, N.J.; Rouget, J.-B.; Dimitrov, J.D.; Wang, N.; Fujimoto, Y.; Hounslow, A.M.; Lacroix-Desmazes, S.; Fukase, K.; et al. Molecular Basis for Bacterial Peptidoglycan Recognition by LysM Domains. Nat. Commun. 2014, 5, 4269. [Google Scholar] [CrossRef]
- Nars, A.; Rey, T.; Lafitte, C.; Vergnes, S.; Amatya, S.; Jacquet, C.; Dumas, B.; Thibaudeau, C.; Heux, L.; Bottin, A.; et al. An Experimental System to Study Responses of Medicago Truncatula Roots to Chitin Oligomers of High Degree of Polymerization and Other Microbial Elicitors. Plant Cell Rep. 2013, 32, 489–502. [Google Scholar] [CrossRef]
- Nars, A.; Lafitte, C.; Chabaud, M.; Drouillard, S.; Mélida, H.; Danoun, S.; Le Costaouëc, T.; Rey, T.; Benedetti, J.; Bulone, V.; et al. Aphanomyces Euteiches Cell Wall Fractions Containing Novel Glucan-Chitosaccharides Induce Defense Genes and Nuclear Calcium Oscillations in the Plant Host Medicago Truncatula. PLoS ONE 2013, 8, e75039. [Google Scholar] [CrossRef] [Green Version]
- Cletus, J.; Balasubramanian, V.; Vashisht, D.; Sakthivel, N. Transgenic Expression of Plant Chitinases to Enhance Disease Resistance. Biotechnol. Lett. 2013, 35, 1719–1732. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanian, V.; Vashisht, D.; Cletus, J.; Sakthivel, N. Plant β-1,3-Glucanases: Their Biological Functions and Transgenic Expression against Phytopathogenic Fungi. Biotechnol. Lett. 2012, 34, 1983–1990. [Google Scholar] [CrossRef] [PubMed]
- Punja, Z.K.; Zhang, Y.Y. Plant Chitinases and Their Roles in Resistance to Fungal Diseases. J. Nematol. 1993, 25, 526–540. [Google Scholar]
- Langner, T.; Göhre, V. Fungal Chitinases: Function, Regulation, and Potential Roles in Plant/Pathogen Interactions. Curr. Genet. 2016, 62, 243–254. [Google Scholar] [CrossRef]
- Chen, C.-S.; Chen, C.-Y.; Ravinath, D.M.; Bungahot, A.; Cheng, C.-P.; You, R.-I. Functional Characterization of Chitin-Binding Lectin from Solanum Integrifolium Containing Anti-Fungal and Insecticidal Activities. BMC Plant Biol. 2018, 18, 3. [Google Scholar] [CrossRef]
- Krieger de Moraes, C.; Schrank, A.; Vainstein, M.H. Regulation of Extracellular Chitinases and Proteases in the Entomopathogen and Acaricide Metarhizium Anisopliae. Curr. Microbiol. 2003, 46, 205–210. [Google Scholar] [CrossRef]
- Barreto, C.C.; Staats, C.C.; Schrank, A.; Vainstein, M.H. Distribution of Chitinases in the Entomopathogen Metarhizium Anisopliae and Effect of N-Acetylglucosamine in Protein Secretion. Curr. Microbiol. 2004, 48, 102–107. [Google Scholar] [CrossRef]
- Dupoiron, S.; Zischek, C.; Ligat, L.; Carbonne, J.; Boulanger, A.; Dugé de Bernonville, T.; Lautier, M.; Rival, P.; Arlat, M.; Jamet, E.; et al. The N-Glycan Cluster from Xanthomonas Campestris Pv. Campestris: A Toolbox for Sequential Plant N-Glycan Processing. J. Biol. Chem. 2015, 290, 6022–6036. [Google Scholar] [CrossRef] [Green Version]
- Boulanger, A.; Zischek, C.; Lautier, M.; Jamet, S.; Rival, P.; Carrère, S.; Arlat, M.; Lauber, E. The Plant Pathogen Xanthomonas Campestris Pv. Campestris Exploits N-Acetylglucosamine during Infection. mBio 2014, 5, e01527-14. [Google Scholar] [CrossRef] [Green Version]
- Boulanger, A.; Déjean, G.; Lautier, M.; Glories, M.; Zischek, C.; Arlat, M.; Lauber, E. Identification and Regulation of the N-Acetylglucosamine Utilization Pathway of the Plant Pathogenic Bacterium Xanthomonas Campestris Pv. Campestris. J. Bacteriol. 2010, 192, 1487–1497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Fernández, L.; Ruiz-Roldán, C.; Pareja-Jaime, Y.; Prieto, A.; Khraiwesh, H.; Roncero, M.I.G. The Fusarium Oxysporum Gnt2, Encoding a Putative N-Acetylglucosamine Transferase, Is Involved in Cell Wall Architecture and Virulence. PLoS ONE 2013, 8, e84690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Fernandez, L.; Roncero, M.I.G.; Prieto, A.; Ruiz-Roldan, C. Comparative Proteomic Analyses Reveal That Gnt2-Mediated N-Glycosylation Affects Cell Wall Glycans and Protein Content in Fusarium Oxysporum. J. Proteom. 2015, 128, 189–202. [Google Scholar] [CrossRef] [PubMed]
- Barghahn, S.; Arnal, G.; Jain, N.; Petutschnig, E.; Brumer, H.; Lipka, V. Mixed Linkage β-1,3/1,4-Glucan Oligosaccharides Induce Defense Responses in Hordeum Vulgare and Arabidopsis Thaliana. Front. Plant Sci. 2021, 12, 682439. [Google Scholar] [CrossRef]
- Del Hierro, I.; Mélida, H.; Broyart, C.; Santiago, J.; Molina, A. Computational Prediction Method to Decipher Receptor-Glycoligand Interactions in Plant Immunity. Plant J. 2021, 105, 1710–1726. [Google Scholar] [CrossRef]
- Rebaque, D.; Del Hierro, I.; López, G.; Bacete, L.; Vilaplana, F.; Dallabernardina, P.; Pfrengle, F.; Jordá, L.; Sánchez-Vallet, A.; Pérez, R.; et al. Cell Wall-Derived Mixed-Linked β-1,3/1,4-Glucans Trigger Immune Responses and Disease Resistance in Plants. Plant J. 2021, 106, 601–615. [Google Scholar] [CrossRef]
- Mélida, H.; Sopeña-Torres, S.; Bacete, L.; Garrido-Arandia, M.; Jordá, L.; López, G.; Muñoz-Barrios, A.; Pacios, L.F.; Molina, A. Non-Branched β-1,3-Glucan Oligosaccharides Trigger Immune Responses in Arabidopsis. Plant J. 2018, 93, 34–49. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microbes | Cell Surface Protein | Organ/Tissue | Disease | References |
|---|---|---|---|---|
| Helicobacter pylori | Mucin 5AC (MUC5AC) | Gastric mucosa | Peptic ulcers | [36] |
| Vibrio cholerae | N-acetylneuraminic acid (Neu5Ac) and N-acetylglucosamine (GlcNAc) | Small intestine | Cholera | [37] |
| Salmonella enterica | MUC1 | Intestinal epithelial cells | Typhoid | [38] |
| Leishmania species | Neuraminidase 1 (NEU 1) | Skin, spleen, and liver | Leishmaniasis | [39] |
| Toxoplasma gondii | Spleen, lung, etc. | Toxoplasmosis | [40,41] | |
| Enterotoxigenic Escherichia coli | MUC2 | Large intestine | Noninflammatory Diarrheas | [42] |
| Candida albicans | Msb2 | Mouth, throat, gut, and vagina | Candidiasis | [43,44] |
| Akkermansia muciniphila | Gastrointestinal mucin | Gastrointestinal tract | - | [45,46] |
| Clostridioides difficile | O-glycan mucin | Gastrointestinal tract | Diarrhea | [47] |
| Staphylococcus aureus | Nasal mucin | Brain, heart, and lung | Pneumonia and Meningitis | [48,49] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ansari, S.; Kumar, V.; Bhatt, D.N.; Irfan, M.; Datta, A. N-Acetylglucosamine Sensing and Metabolic Engineering for Attenuating Human and Plant Pathogens. Bioengineering 2022, 9, 64. https://doi.org/10.3390/bioengineering9020064
Ansari S, Kumar V, Bhatt DN, Irfan M, Datta A. N-Acetylglucosamine Sensing and Metabolic Engineering for Attenuating Human and Plant Pathogens. Bioengineering. 2022; 9(2):64. https://doi.org/10.3390/bioengineering9020064
Chicago/Turabian StyleAnsari, Sekhu, Vinay Kumar, Dharmendra Nath Bhatt, Mohammad Irfan, and Asis Datta. 2022. "N-Acetylglucosamine Sensing and Metabolic Engineering for Attenuating Human and Plant Pathogens" Bioengineering 9, no. 2: 64. https://doi.org/10.3390/bioengineering9020064
APA StyleAnsari, S., Kumar, V., Bhatt, D. N., Irfan, M., & Datta, A. (2022). N-Acetylglucosamine Sensing and Metabolic Engineering for Attenuating Human and Plant Pathogens. Bioengineering, 9(2), 64. https://doi.org/10.3390/bioengineering9020064