
Meniscus Matrix Structural and Biomechanical Evaluation: Age-Dependent Properties in a Swine Model

 , , , , , and
, , , , , and 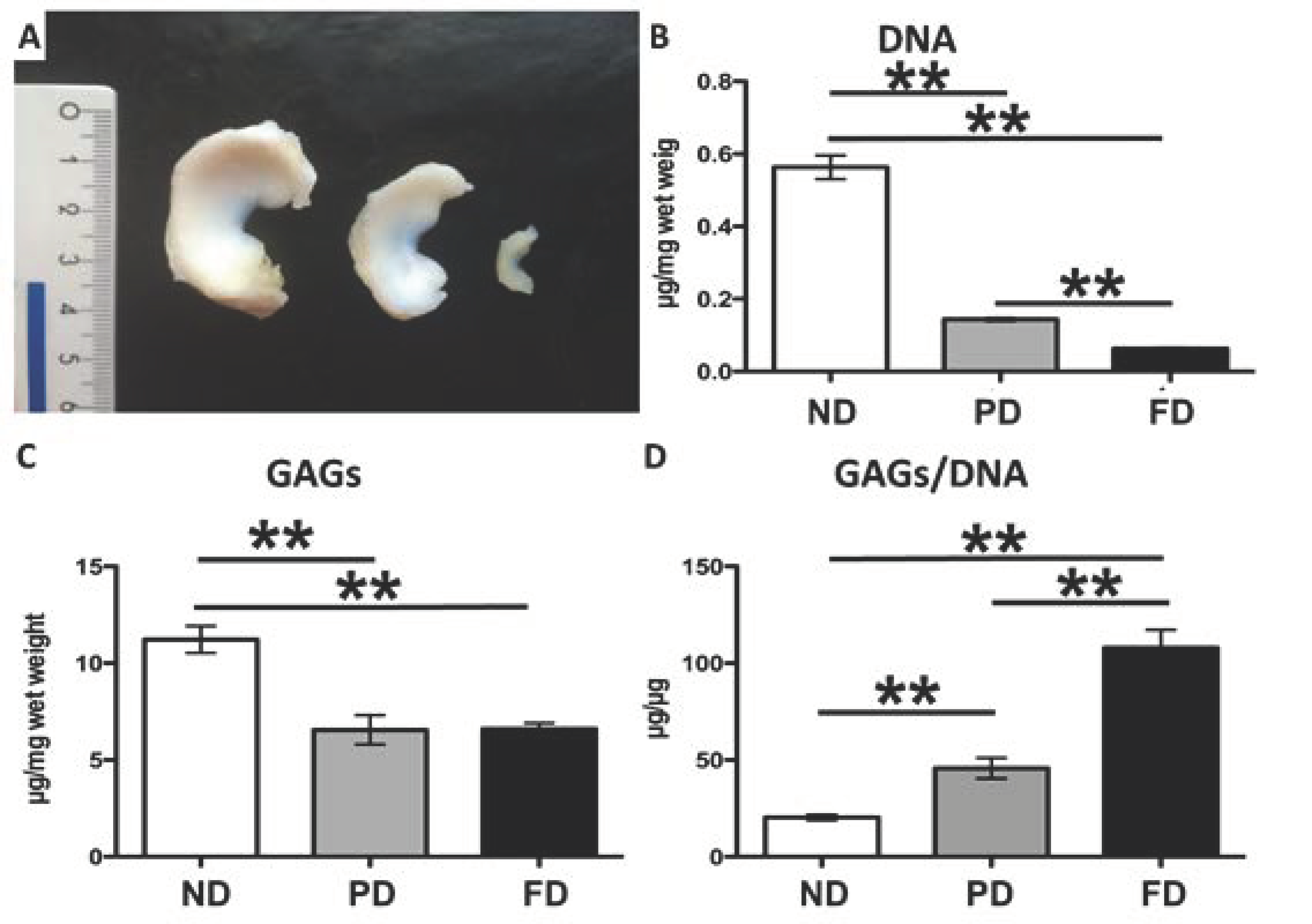{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biochemical Analyses
2.2. Protein Extraction and Western Blot
2.3. Micro Anatomical Analysis: Immunohistochemistry
2.4. Biomechanical Analysis
2.5. Statistical Analyses
3. Results
3.1. Biochemical Analyses
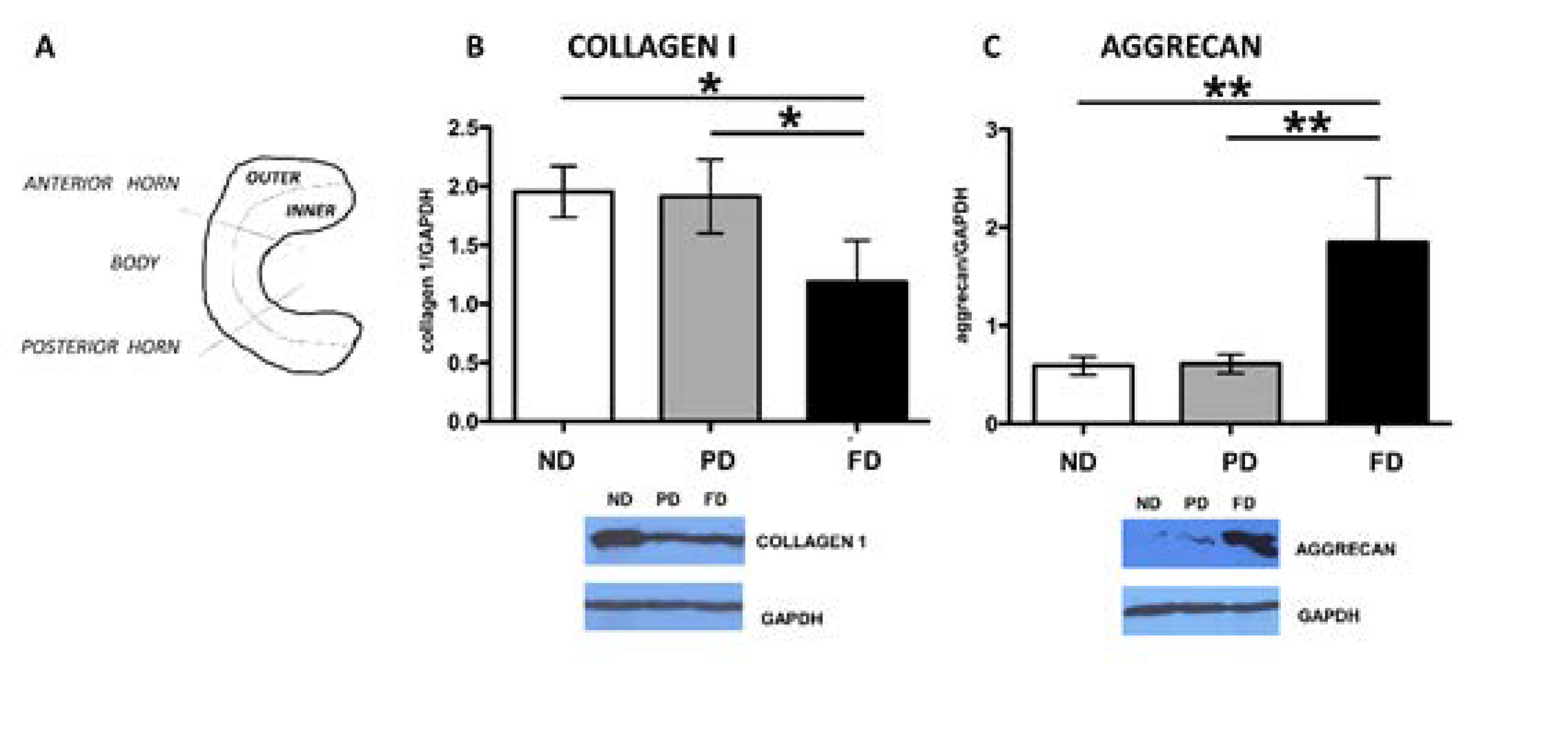
3.2. Western Blot
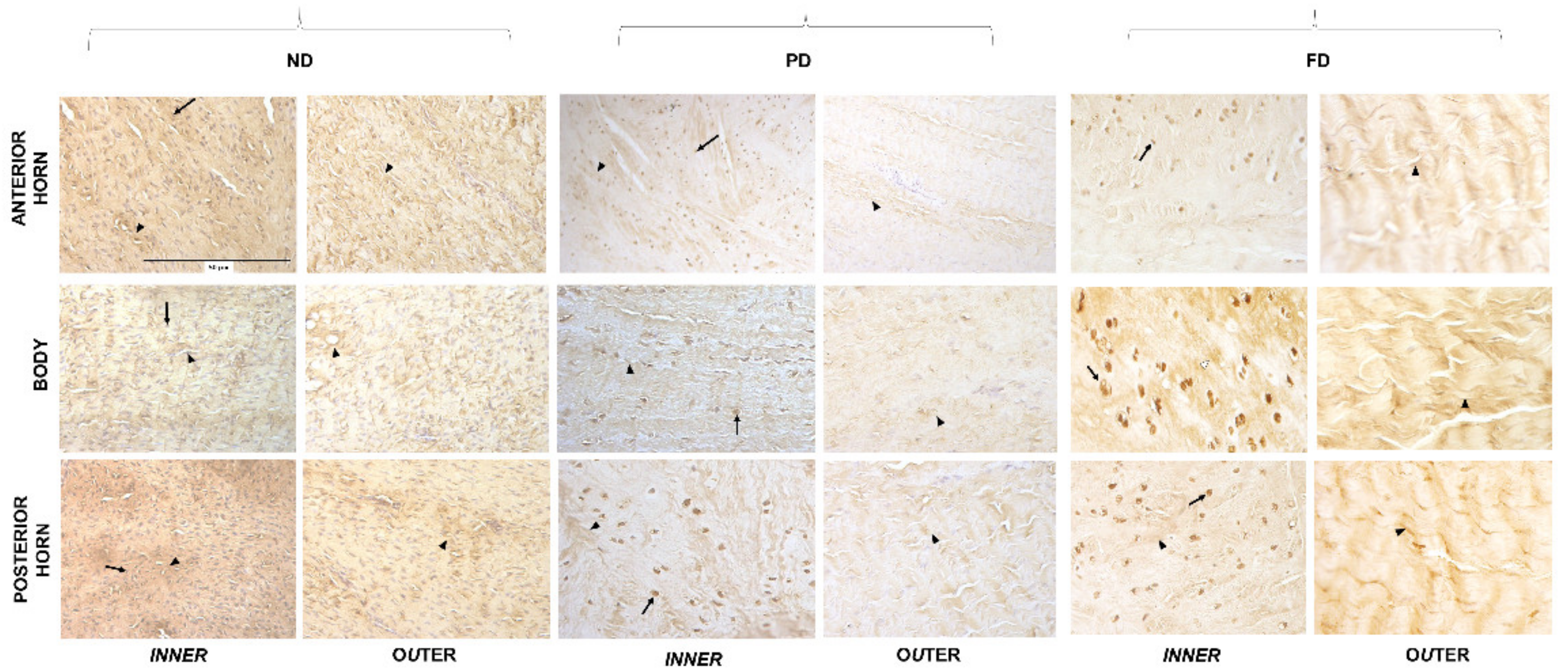
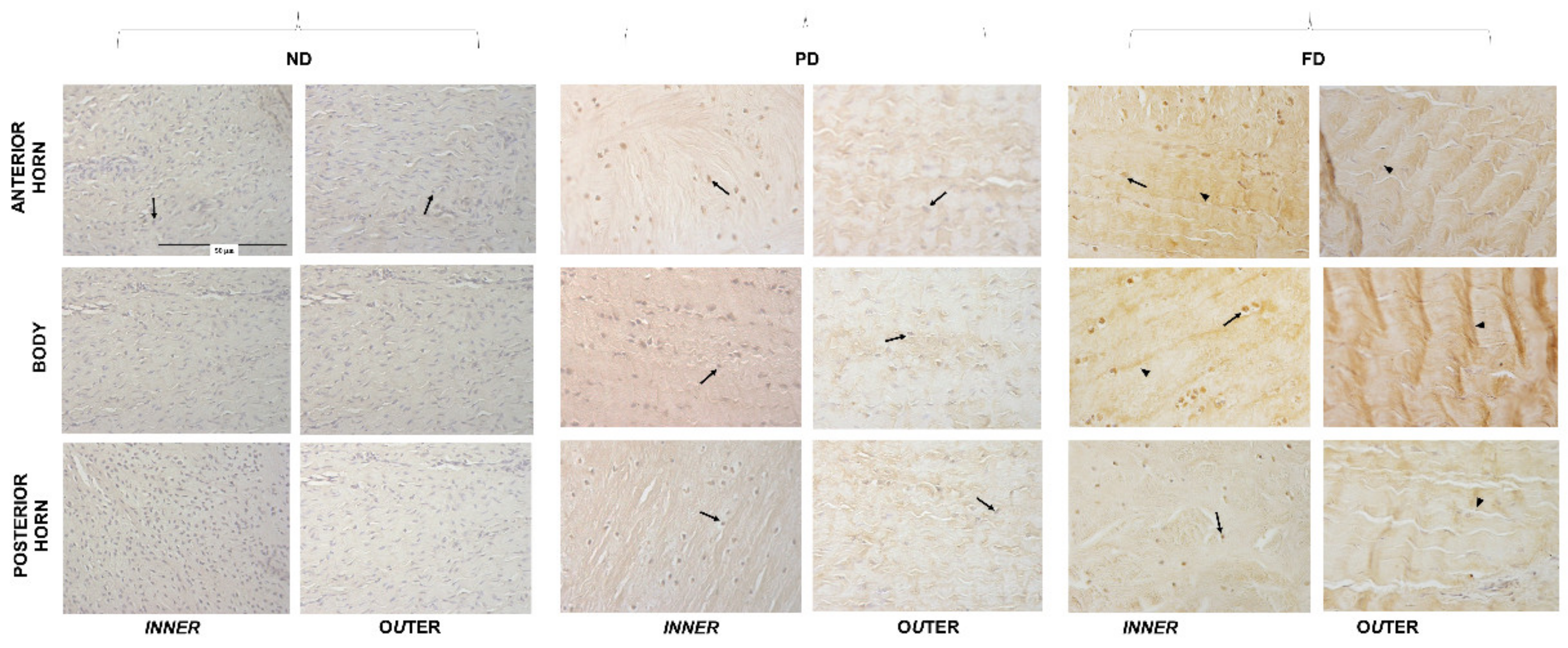
3.3. Microanatomical Analysis: Immunohistochemistry
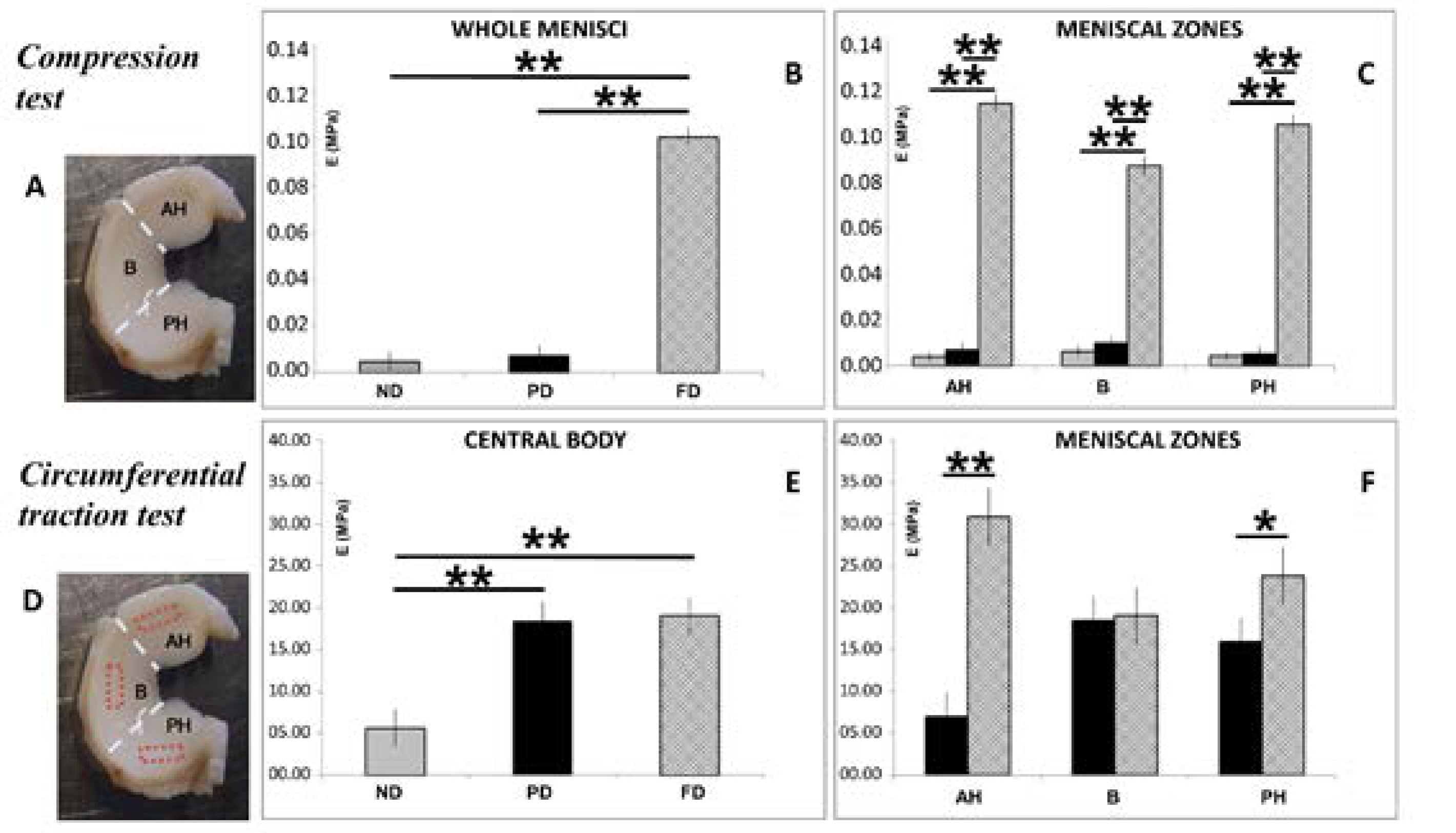
3.4. Biomechanical Analysis: Compression and Traction Tests
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Proffen, B.L.; McElfresh, M.; Fleming, B.C.; Murray, M.M. A comparative anatomical study of the human knee and six animal species. Knee 2012, 19, 493–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshioka, S.; Nagano, A.; Himeno, R.; Fukashiro, S. Computation of the Kinematics and the Minimum Peak Joint Moments of Sit-to-Stand Movements. Biomed. Eng. Online 2007, 6, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fox, A.J.S.; Bedi, A.; Rodeo, S.A. The Basic Science of Human Knee Menisci. Sports Health 2012, 4, 340–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howell, R.; Kumar, N.S.; Patel, N.; Tom, J. Degenerative Meniscus: Pathogenesis, Diagnosis, and Treatment Options. World J. Orthop. 2014, 5, 597–602. [Google Scholar] [CrossRef]
- Peretti, G.M.; Tessaro, I.; Montanari, L.; Polito, U.; Di Giancamillo, A.; Di Giancamillo, M.; Marmotti, A.; Montaruli, A.; Roveda, E.; Mangiavini, L. Histological Changes of the Meniscus Following an Osteochondral Lesion. J. Biol. Regul. Homeost. Agents 2017, 31, 129–134. [Google Scholar]
- Di Giancamillo, A.; Mangiavini, L.; Tessaro, I.; Marmotti, A.; Scurati, R.; Peretti, G.M. The Meniscus Vascularization: The Direct Correlation with Tissue Composition for Tissue Engineering Purposes. J. Biol. Regul. Homeost. Agents 2016, 30, 85–90. [Google Scholar]
- McNulty, A.L.; Guilak, F. Mechanobiology of the Meniscus. J. Biomech. 2015, 48, 1469–1478. [Google Scholar] [CrossRef]
- Abraham, A.C.; Edwards, C.R.; Odegard, G.M.; Haut Donahue, T.L. Regional and Fiber Orientation Dependent Shear Properties and Anisotropy of Bovine Meniscus. J. Mech. Behav. Biomed. Mater. 2011, 4, 2024–2030. [Google Scholar] [CrossRef] [Green Version]
- Valiyaveettil, M.; Mort, J.S.; McDevitt, C.A. The Concentration, Gene Expression, and Spatial Distribution of Aggrecan in Canine Articular Cartilage, Meniscus, and Anterior and Posterior Cruciate Ligaments: A New Molecular Distinction between Hyaline Cartilage and Fibrocartilage in the Knee Joint. Connect. Tissue Res. 2005, 46, 83–91. [Google Scholar] [CrossRef]
- Roughley, P.J.; Mort, J.S. The Role of Aggrecan in Normal and Osteoarthritic Cartilage. J. Exp. Orthop. 2014, 1, 8. [Google Scholar] [CrossRef] [Green Version]
- Melrose, J.; Smith, S.; Cake, M.; Read, R.; Whitelock, J. Comparative Spatial and Temporal Localisation of Perlecan, Aggrecan and Type I, II and IV Collagen in the Ovine Meniscus: An Ageing Study. Histochem. Cell Biol. 2005, 124, 225–235. [Google Scholar] [CrossRef]
- Shrive, N.G.; O’connor, J.J.; Goodfellow, J.W. Load-Bearing in the Knee Joint. Clin. Orthop. Relat. Res. 1978, 131, 279–287. [Google Scholar] [CrossRef]
- Arnoczky, S.P.; Warren, R.F. The Microvasculature of the Meniscus and Its Response to Injury: An Experimental Study in the Dog. Am. J. Sports Med. 1983, 11, 131–141. [Google Scholar] [CrossRef]
- Pufe, T.; Petersen, W.J.; Miosge, N.; Goldring, M.B.; Mentlein, R.; Varoga, D.J.; Tillmann, B.N. Endostatin/Collagen XVIII--an Inhibitor of Angiogenesis--Is Expressed in Cartilage and Fibrocartilage. Matrix Biol. 2004, 23, 267–276. [Google Scholar] [CrossRef]
- Smith, S.M.; Shu, C.; Melrose, J. Comparative Immunolocalisation of Perlecan with Collagen II and Aggrecan in Human Foetal, Newborn and Adult Ovine Joint Tissues Demonstrates Perlecan as an Early Developmental Chondrogenic Marker. Histochem. Cell. Biol. 2010, 134, 251–263. [Google Scholar] [CrossRef]
- Di Giancamillo, A.; Deponti, D.; Modina, S.; Tessaro, I.; Domeneghini, C.; Peretti, G.M. Age-Related Modulation of Angiogenesis-Regulating Factors in the Swine Meniscus. J. Cell Mol. Med. 2017, 21, 3066–3075. [Google Scholar] [CrossRef]
- Deponti, D.; Di Giancamillo, A.; Scotti, C.; Peretti, G.M.; Martin, I. Animal Models for Meniscus Repair and Regeneration. J. Tissue Eng. Regen. Med. 2015, 9, 512–527. [Google Scholar] [CrossRef]
- Yu, H.; Adesida, A.B.; Jomha, N.M. Meniscus Repair Using Mesenchymal Stem Cells—A Comprehensive Review. Stem Cell Res. Ther. 2015, 6, 86. [Google Scholar] [CrossRef] [Green Version]
- Ding, Z.; Huang, H. Mesenchymal Stem Cells in Rabbit Meniscus and Bone Marrow Exhibit a Similar Feature but a Heterogeneous Multi-Differentiation Potential: Superiority of Meniscus as a Cell Source for Meniscus Repair. BMC Musculoskelet. Disord. 2015, 16, 65. [Google Scholar] [CrossRef] [Green Version]
- Wei, L.-C.; Gao, S.-G.; Xu, M.; Jiang, W.; Tian, J.; Lei, G.-H. A Novel Hypothesis: The Application of Platelet-Rich Plasma Can Promote the Clinical Healing of White-White Meniscal Tears. Med. Sci. Monit. 2012, 18, HY47–HY50. [Google Scholar] [CrossRef] [Green Version]
- Cerejo, R.; Dunlop, D.D.; Cahue, S.; Channin, D.; Song, J.; Sharma, L. The Influence of Alignment on Risk of Knee Osteoarthritis Progression According to Baseline Stage of Disease. Arthritis Rheum. 2002, 46, 2632–2636. [Google Scholar] [CrossRef]
- Ferroni, M.; Belgio, B.; Peretti, G.M.; Di Giancamillo, A.; Boschetti, F. Evolution of Meniscal Biomechanical Properties with Growth: An Experimental and Numerical Study. Bioengineering 2021, 8, 70. [Google Scholar] [CrossRef]
- Gunja, N.J.; Athanasiou, K.A. Passage and Reversal Effects on Gene Expression of Bovine Meniscal Fibrochondrocytes. Arthritis Res. Ther. 2007, 9, R93. [Google Scholar] [CrossRef] [Green Version]
- Bland, Y.S.; Ashhurst, D.E. Changes in the Content of the Fibrillar Collagens and the Expression of Their MRNAs in the Menisci of the Rabbit Knee Joint during Development and Ageing. Histochem. J. 1996, 28, 265–274. [Google Scholar] [CrossRef]
- Kiani, C.; Chen, L.; Wu, Y.J.; Yee, A.J.; Yang, B.B. Structure and Function of Aggrecan. Cell Res. 2002, 12, 19–32. [Google Scholar] [CrossRef] [Green Version]
- Giorgi, M.; Carriero, A.; Shefelbine, S.J.; Nowlan, N.C. Mechanobiological Simulations of Prenatal Joint Morphogenesis. J. Biomech. 2014, 47, 989–995. [Google Scholar] [CrossRef] [Green Version]
- Sosio, C.; Di Giancamillo, A.; Deponti, D.; Gervaso, F.; Scalera, F.; Melato, M.; Campagnol, M.; Boschetti, F.; Nonis, A.; Domeneghini, C.; et al. Osteochondral Repair by a Novel Interconnecting Collagen–Hydroxyapatite Substitute: A Large-Animal Study. Tissue Eng. Part. A 2015, 21, 704–715. [Google Scholar] [CrossRef]
- Deponti, D.; Di Giancamillo, A.; Mangiavini, L.; Pozzi, A.; Fraschini, G.; Sosio, C.; Domeneghini, C.; Peretti, G.M. Fibrin-Based Model for Cartilage Regeneration: Tissue Maturation from In Vitro to In Vivo. Tissue Eng. Part. A 2012, 18, 1109–1122. [Google Scholar] [CrossRef] [PubMed]
- Cheung, H.S. Distribution of Type I, II, III and V in the Pepsin Solubilized Collagens in Bovine Menisci. Connect. Tissue Res. 1987, 16, 343–356. [Google Scholar] [CrossRef]
- Di Giancamillo, A.; Deponti, D.; Addis, A.; Domeneghini, C.; Peretti, G.M. Meniscus Maturation in the Swine Model: Changes Occurring along with Anterior to Posterior and Medial to Lateral Aspect during Growth. J. Cell Mol. Med. 2014, 18, 1964–1974. [Google Scholar] [CrossRef]
- McAlinden, A.; Dudhia, J.; Bolton, M.C.; Lorenzo, P.; Heinegård, D.; Bayliss, M.T. Age-Related Changes in the Synthesis and MRNA Expression of Decorin and Aggrecan in Human Meniscus and Articular Cartilage. Osteoarthr. Cartil. 2001, 9, 33–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hellio Le Graverand, M.P.; Ou, Y.; Schield-Yee, T.; Barclay, L.; Hart, D.; Natsume, T.; Rattner, J.B. The Cells of the Rabbit Meniscus: Their Arrangement, Interrelationship, Morphological Variations and Cytoarchitecture. J. Anat. 2001, 198, 525–535. [Google Scholar] [CrossRef] [PubMed]
- Sizeland, K.H.; Wells, H.C.; Kirby, N.M.; Hawley, A.; Mudie, S.T.; Ryan, T.M.; Haverkamp, R.G. Bovine Meniscus Middle Zone Tissue: Measurement of Collagen Fibril Behavior During Compression. IJN 2020, 15, 5289–5298. [Google Scholar] [CrossRef] [PubMed]
- Gee, S.M.; Posner, M. Meniscus Anatomy and Basic Science. Sports Med. Arthrosc. Rev. 2021, 29, e18. [Google Scholar] [CrossRef]
- Scott, W.N. Insall & Scott Surgery of the Knee E-Book; Elsevier Health Sciences: Philadelphia, PA, USA, 2011; ISBN 978-1-4557-2789-6. [Google Scholar]
- Sweigart, M.A.; Athanasiou, K.A. Biomechanical Characteristics of the Normal Medial and Lateral Porcine Knee Menisci. Proc. Inst. Mech. Eng. H J. Eng. Med. 2005, 219, 53–62. [Google Scholar] [CrossRef]
- Guilak, F.; Cohen, D.M.; Estes, B.T.; Gimble, J.M.; Liedtke, W.; Chen, C.S. Control of Stem Cell Fate by Physical Interactions with the Extracellular Matrix. Cell Stem Cell 2009, 5, 17–26. [Google Scholar] [CrossRef] [Green Version]
- Boudreau, N.J.; Jones, P.L. Extracellular Matrix and Integrin Signalling: The Shape of Things to Come. Biochem. J. 1999, 339, 481–488. [Google Scholar] [CrossRef]
- Streuli, C. Extracellular Matrix Remodelling and Cellular Differentiation. Curr. Opin. Cell Biol. 1999, 11, 634–640. [Google Scholar] [CrossRef]
- Bissel, M.J.; Barcellos-Hoff, M.H. The Influence of Extracellular Matrix on Gene Expression: Is Structure the Message? J. Cell Sci. 1987, 1987, 327–343. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.Q.; Bissell, M.J. Multi-Faceted Regulation of Cell Differentiation by Extracellular Matrix. FASEB J. 1993, 7, 737–743. [Google Scholar] [CrossRef]
- Tan, G.-K.; Cooper-White, J.J. Interactions of Meniscal Cells with Extracellular Matrix Molecules: Towards the Generation of Tissue Engineered Menisci. Cell Adh. Migr. 2011, 5, 220–226. [Google Scholar] [CrossRef] [Green Version]
- Mauck, R.L.; Burdick, J.A. From Repair to Regeneration: Biomaterials to Reprogram the Meniscus Wound Microenvironment. Ann. Biomed. Eng. 2015, 43, 529–542. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aidos, L.; Modina, S.C.; Millar, V.R.H.; Peretti, G.M.; Mangiavini, L.; Ferroni, M.; Boschetti, F.; Di Giancamillo, A. Meniscus Matrix Structural and Biomechanical Evaluation: Age-Dependent Properties in a Swine Model. Bioengineering 2022, 9, 117. https://doi.org/10.3390/bioengineering9030117
Aidos L, Modina SC, Millar VRH, Peretti GM, Mangiavini L, Ferroni M, Boschetti F, Di Giancamillo A. Meniscus Matrix Structural and Biomechanical Evaluation: Age-Dependent Properties in a Swine Model. Bioengineering. 2022; 9(3):117. https://doi.org/10.3390/bioengineering9030117
Chicago/Turabian StyleAidos, Lucia, Silvia Clotilde Modina, Valentina Rafaela Herrera Millar, Giuseppe Maria Peretti, Laura Mangiavini, Marco Ferroni, Federica Boschetti, and Alessia Di Giancamillo. 2022. "Meniscus Matrix Structural and Biomechanical Evaluation: Age-Dependent Properties in a Swine Model" Bioengineering 9, no. 3: 117. https://doi.org/10.3390/bioengineering9030117
APA StyleAidos, L., Modina, S. C., Millar, V. R. H., Peretti, G. M., Mangiavini, L., Ferroni, M., Boschetti, F., & Di Giancamillo, A. (2022). Meniscus Matrix Structural and Biomechanical Evaluation: Age-Dependent Properties in a Swine Model. Bioengineering, 9(3), 117. https://doi.org/10.3390/bioengineering9030117