1. Introduction
Fermented foods and beverages have become a major feature of a consumer trend known as the “wellness lifestyle”, whereby individuals make dietary choices with the intention of pursuing or maintaining a health goal [
1]. This is particularly true of non-alcoholic fermented products that contain, or are perceived to contain, live beneficial microorganisms at the point of retail sale. Kombucha, a fermented tea-based beverage, falls into this category and is generally considered to be a functional food [
2].
Research into the functionality of kombucha has explored a range of perceived health benefits. While there is a lack of supporting clinical data [
3], in vitro studies have demonstrated that tea polyphenols are potent antioxidants [
4], fermented kombucha inhibits the growth of foodborne pathogens [
5], and kombucha extracts exhibit antiproliferative effects on cancer cells [
6], along with an array of other bioactivities. In addition, some recent animal studies have indicated that the consumption of kombucha can alter the gut microbiome and alleviate disease states, results that align with the generalized concept that fermented foods containing live microbes exert their impacts via direct interactions with the gastrointestinal tract and its microbiota [
7,
8]. Nevertheless, the potential for a fermented food to provide gut- and general-health benefits does not equate to probiotic status, which is reserved for systems where a known dosage of specific microbes with demonstrated health benefits can be assured [
9]. Are the terms “live”, “contains probiotics”, and “probiotic beverage”, often used to describe kombucha [
2], warranted based on how it is produced?
The traditional process of kombucha production involves the inoculation of sweetened tea with a cellulosic pellicle harboring an undefined microbial community known as the SCOBY (Symbiotic Culture of Bacteria and Yeast) [
10,
11,
12]. Upon the completion of fermentation, these microbes remain part of the tea, which is adjusted for preferred organoleptic properties by the addition of flavorants and non-caloric sweeteners [
11] and then carbonated and bottled for commercial sale. Although microbial taxa within the kombucha pellicle may vary between producers, the most prevalent belong to the acetic acid bacteria (AAB)
Komagataeibacter and the fermentative yeast
Brettanomyces [
13,
14]. Neither genus has been associated with physiological impacts on ingestion or health benefits. While few studies have profiled the microbiota of retail-available kombuchas, Andreson et al. [
15] found that
Brettanomyces was the dominant yeast genus in 16 European samples. In the same study,
Komagataeibacter was often displaced by
Bacilli and
Lactobacilli, possibly reflecting that some producers inoculate before packaging with known probiotic strains of
Bacillus coagulans,
B. subtilis,
Lacticaseibacillus rhamnosus, and
S. cerevisae var
boulardii [
2].
“Hard”, or alcoholic, kombucha is a relatively new beverage category, and its production practices are not described in the scientific literature. The process of the hard kombucha production workflow and how it compares to the soft kombucha production workflow is postulated in
Figure 1. It is likely that some hard kombuchas on the market reflect a minimal extension of the traditional process of kombucha production, whereby an anaerobic phase without refrigeration is incorporated to allow ethanol to accumulate beyond 0.5-percent alcohol by volume (ABV). This process essentially takes advantage of a known problem in the “soft” (non-alcoholic) kombucha sector: post-packaging cold-chain failures that result in retail-available products that exceed the maximum allowed ABV [
7]. It is also likely that hard kombucha is produced by the intentional inoculation of soft kombucha with the pre-eminent fermentation yeast species,
S. cerevisiae [
16]. To date, no studies have examined the chemical or microbial composition of finished retail-available hard kombucha.
Consumers seeking to embrace a “wellness lifestyle” are faced with a growing array of kombucha-based products, including “hard” styles containing alcohol. In this study, we surveyed kombucha beverages available at retail outlets in Oregon, USA, profiling their chemical and microbial compositions to address two hypotheses. First, we asked whether the chemical and microbial contents of hard kombucha differed from those found in soft kombucha. Then, we asked whether package claims of “live” or “probiotic”, or the specific mention of probiotic species and/or strains in the package ingredient lists, were predictive of the microbial content. Our results highlight the variability between retail-available kombucha products in terms of their composition and associated package claims, providing data that can guide further research focused on the nutritional and health outcomes of kombucha consumption.
2. Materials and Methods
2.1. Chemicals, Reagents, and Microbiological Media
Unless stated otherwise, all chemicals and reagents were obtained from Sigma-Aldrich (St. Louis, MO, USA). The standard yeast differential medium WLN (Millipore Burlington, MA, USA) agar was prepared by dissolving 77 g/L in deionized water. M13 agar for the cultivation of acetic acid bacteria contained 25 g/L d-mannitol, 5 g/L yeast extract, and 3 g/L peptone in deionized water, with the addition of 15 g/L agar (Bioplus, Altamonte Springs, FL, USA). Post-autoclave, M13 agar was supplemented with 2.0% (volume by volume) acetic acid and 2.0% (v/v) ethanol (Oregon State University Chemistry Store, Corvallis, OR, USA) prior to pouring into Petri dishes. Nutrient agar (NA; Research Products International, Mount Prospect, IL, USA) for the cultivation of spore-forming Bacilli was prepared by dissolving 23 g/L in deionized water with 15 g/L agar. MRS broth (Research Products International, Mount Prospect, IL, USA) for the cultivation of lactic acid bacteria was prepared by dissolving 55 g/L in deionized water. All media were sterilized by autoclaving at 121 °C for 15 min.
2.2. Kombucha Sampling and Processing
All kombucha products were purchased from retail stores in the Willamette Valley (Oregon, USA) in January 2021. A preliminary analysis of the ingredient lists of all kombucha products available at one online retailer (
wholefoodsmarket.com, accessed on 11 March 2023) was used to define the prevalent flavor categories: “berry/fruity”, “ginger/citrus”, and “herbal/tea”. Brands available in the marketplace in Oregon were representatively sampled by purchasing products for each brand from within the prevalent flavor categories. Overall, 39 unique kombucha products from 21 brands were obtained (
Table 1). One hard kombucha product and two soft kombucha products were repeat-sampled from different packages, providing an additional 7 samples for the evaluation of packaged product consistency. On-package claims of “live” or “probiotic” and the specific mention of probiotic cultures (species and strain, CFU count) in the ingredient lists were recorded as metadata (
Table S1).
Each kombucha product was immediately transported in a cooler to Oregon State University following its purchase, where it was refrigerated at 4 °C until opened for sampling. Packaged products were agitated to homogenize contents and then allowed to rest for 2 min prior to being opened aseptically in a laminar-flow hood. Then, 50 mL was transferred into sterile gas-exchange “bioreactor” tubes (Tubespin, TPP Techno Plastic Products AG, Trasadingen, Switzerland) and centrifuged at 3900 RPM for 15 min to remove carbon dioxide. Degassed samples were vortexed and subsampled for chemical and microbiological analyses, and then the remainder of each was stored at 4 °C for resampling if required.
Subsamples were used immediately for dilution and standard plate counting. Additional subsamples for 1H NMR spectroscopy and DNA extraction were centrifuged at 15,000 RPM for 5 min. Supernatants were transferred to fresh tubes, and then supernatants and pellets were stored at −20 °C until use.
2.3. Enumeration of Culturable Yeast and Bacteria
For enumeration on Wallerstein (WLN), de Man Rogosa and Sharpe (MRS), Mannitol Agar (M13), and Nutrient Agar (NA) media, 39 unique samples and an additional 7 repeat samples were serially diluted in 0.1% sterile peptone water, and 100 µL of the appropriate dilution was spread onto the surface of each plate to generate 25–250 colony-forming units (CFU). For enumeration on MRS, samples were centrifuged, resuspended and serially diluted in MRS broth, and then 1 mL of the appropriate dilution was transferred to the surface of Aerobic Plate Count (APC) Petrifilm (3M, St. Paul, MN, USA) to generate <300 CFU. WLN and M13 plates were incubated aerobically at 27 °C for 7 days, and MRS-loaded APC Petrifilms were incubated anaerobically at 27 °C for 7 days in a BD GasPak EZ Container with EZ Gaskpak indicator sachets (Becton Dickinson and Company, Franklin Lakes, NJ, USA). NA plates were incubated aerobically at 27 °C for 2 days. All samples that did not generate a CFU count, or where CFU counts were out of range, were immediately repeat-plated from degassed reserve samples to confirm the lack of culturable cells or obtain counts within the target range.
2.4. 1H NMR Spectroscopy Analysis of Chemical Composition
2.4.1. Acquisition of Chemical Composition by 1H NMR Spectroscopy
The frozen supernatants of 39 unique samples and 7 additional repeat samples were thawed and incubated at room temperature. Samples were normalized to pH 7 using phosphate buffer (1.88 mM NaHPO•HO, 5.8 mM NaHPO, pH 7.1). Then, 540 µL of each sample was transferred to a 5 mm NMR tube (Norell, Morganton, NC, USA) with 60 µL of 0.25 mM 4,4-dimethyl-4-silapentane-1-sulfonic acid (DSS) in 90% HO/10% deuterium oxide (Chenomx Inc., Edmonton, Alberta, Canada). All spectra were acquired at 300 K using a Bruker Avance 800 MHz NMR spectrometer (Bruker BioSpin, Rheinstetten, Germany) operating at 800 MHz. NMR data were acquired with 128 scans at 32 K data points, a 1H spectral window of 9600 Hz, a 12.1 ppm spectral width, and a dwell time interval of 52 s between data point collections and a recovery (D1) delay of 5 s between acquisitions.
2.4.2. Analysis of 1H NMR Spectra
NMR spectra were Fourier transformed and automatically phased using TOPSPIN 3.7 software (Leyton, London, UK). Spectra were corrected for using Chenomx NMR suite 8.2 software (Edmonton, AB, Canada) with the DSS internal standard. Automatic phase correction, line broadening, water deletion, and baseline correction were performed on 39 samples according to the chemical form indicator. Some manual adjustments to the phase were used where necessary. Analytes were identified and quantified by fitting the 1D 1H spectral patterns, chemical shifts, and spectral intensities to references from the Chenomx spectral database for 800 MHz NMR spectrometers.
Principal component analysis (PCA) was applied to NMR analytes matched to reference spectra using “prcomp” from the package “stats” in R (Boston, MA, USA) [
17]. The Wilcoxon rank-sum test, Kruskal–Wallis test, and multivariate analysis of variance (MANOVA) were employed for testing differences between “Types”, “Claim”, and “Organism listed” groups of kombucha. Ethanol gave a triplet signal at 1.20 ppm and a multiplet signal at 3.66 without noise from other analytes [
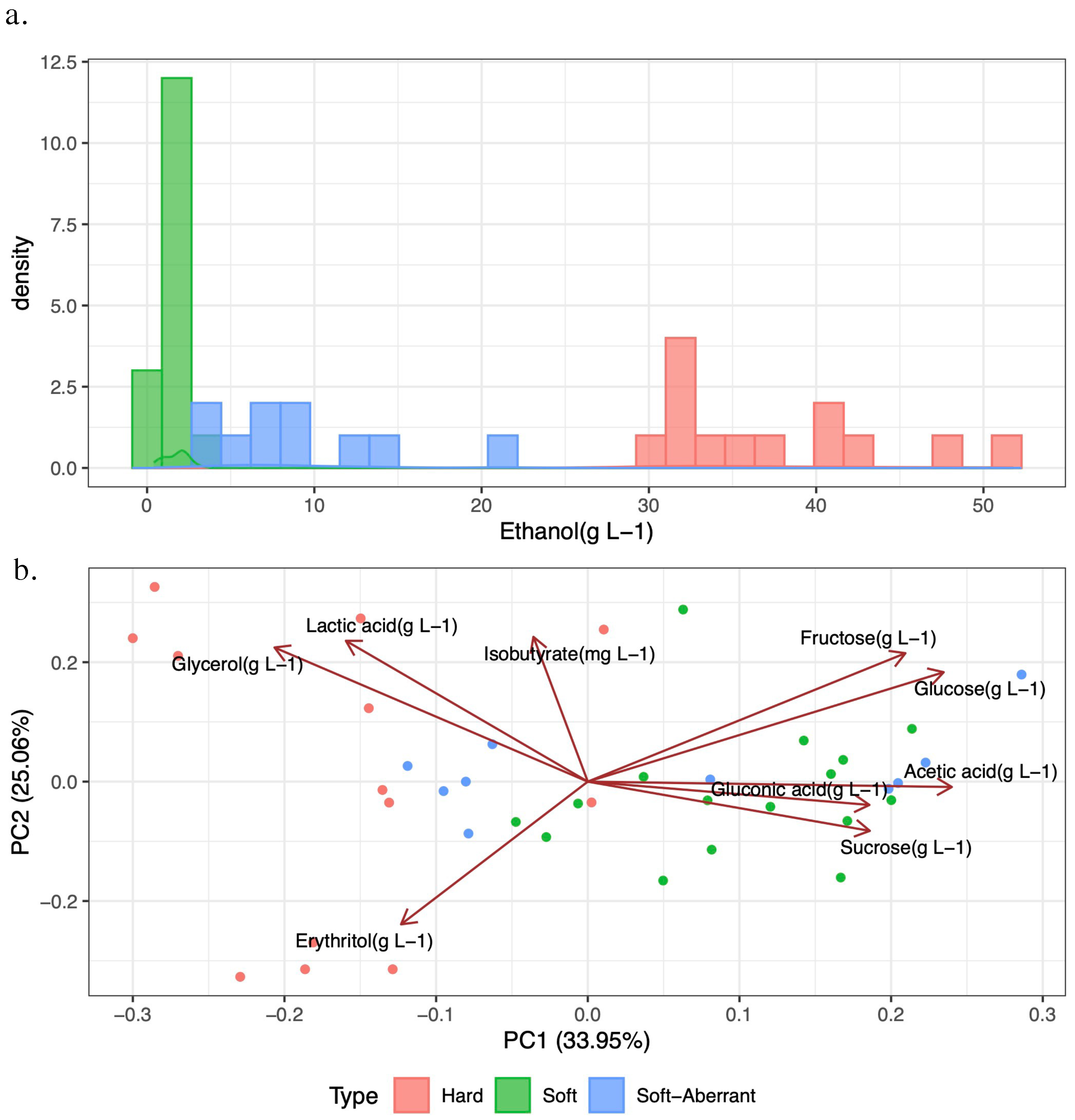
18]. The ABV for ethanol was calculated from g/L, and 0.5 ± 0.05% was used as the threshold for defining soft and soft-aberrant kombucha types. The cutoff used for the inclusion of organic acids in this study was 0.1 mg/L. Organic acids that fell below this threshold were excluded from analysis.
2.5. DNA Extraction for Microbiome Analysis
Centrifuged pellets for 39 unique samples and 7 additional repeat samples (see
Section 2.2) were resuspended in molecular-grade water, and DNA was extracted using the DNeasy Power Foods Microbial kit (QIAGEN, Hilden, Germany) according to the manufacturer’s instructions with minor modifications. In place of vortexing, the Omni Bead Ruptor 24 (Omni International, Inc., Kennesaw, GA, USA) was used with 15 s pulses at 8.0 m/s with a 55 s pause and 10 cycles. All DNA extracts were quantified using the SpectraMax Quant Accuclear Nano dsDNA assay kit (Molecular Devices, San Jose, CA, USA) on the SpectraMax M2 (Molecular Devices, San Jose, CA, USA) with excitation and emission at 468 and 507 nm, respectively. The ZymoBIOMICS Microbial Community Standard (Zymo Research, Irvine, CA, USA) was included as a positive control for DNA extraction. DNA extracts were stored at
°C until further use.
2.6. Metabarcoding Sequencing and Data Analysis
2.6.1. Metabarcoding Sequence Processing
Libraries for metabarcoding analyses were prepared as described by Comeau et al. [
19], with minor modifications. For the analysis of fungal communities, the fungal ITS2 region was amplified using BITS-F and B58S3-R primers [
20]. For the analysis of bacterial communities, the bacterial 16S V4–V5 domain was amplified using F515 and R926 primers [
21]. Forward and reverse fusion primers were designed using the scheme recommended by Comeau et al. [
19] for Illumina Nextera XT v2 indices and Nextera adapters (
Table S2).
Each DNA extract was amplified twice (reactions using 1× and 1/10× DNA concentration) using 12.5 µL of Platinum Hot Start 2× master mix (Thermo Scientific, Waltham, MA, USA), 1 µM of each primer, 2 µL of template DNA, and molecular-biology-grade water (Biotium, Hayward, NJ, USA) in a final volume of 25 µL. Successful amplification of DNA templates was verified by visualizing products on 1.5% agarose gels using 6× GelRed Nucleic Acid Stain (Biotium, Hayward, NJ, USA). Samples not successfully amplified were excluded from downstream processing (n = 2). Duplicate PCR products of 37 samples were combined, purified, and normalized using SequalPrep Normalization Plates (Thermo Scientific, Waltham, MA, USA) according to the manufacturer’s instructions. Pooled libraries were quantified using the Qubit dsDNA HS assay (Thermo Scientific, Waltham, MA, USA), and the median fragment size was determined on the Bioanalyzer 2100 high-sensitivity DNA assay (Agilent, Santa Clara, CA, USA). The pooled fungal ITS2 and bacterial 16S libraries were sequenced separately using MiSeq 2 × 300 bp v3 chemistry by the Oregon State University Center for Quantitative Life Sciences (CQLS, Corvallis, OR, USA) with a 5% PhiX spike-in, according to standard Illumina protocols.
2.6.2. Post-Sequencing Processing
Paired-end reads were trimmed and merged, and chimeric sequences were removed using the DADA2 plugin in QIIME2 (qiime dada2 denoise-paired) [
22]. 16S OTUs were clustered at 100% sequence similarity (amplicon sequence variants) using DADA2 and classified using the QIIME2 pre-trained naïve Bayes classifier trained on the Greengenes database (13_8) [
23]. ITS OTUs were classified using the classifier trained on UNITE (ver8) with q2-feature-classifier [
24].
2.7. Metabarcoding Sequence Analysis
Metabarcoding data visualization was performed in R v4.2.1 following the Phyloseq and Microviz tutorials (
https://vaulot.github.io/tutorials/Phyloseq_tutorial.html and
https://david-barnett.github.io/microViz/articles/shao19-analyses.html, accessed on 11 March 2023 [
25,
26]. The function filter_taxa was used to remove taxa not seen more than 5 times in at least 25% of the samples. Two samples were excluded for having fewer than 500 reads [
27]. To determine species-level assignments for 16S feature IDs, a BLAST search using the command blastn and NIH nt.fsa database was used [
28]. Species richness (Chao1), alpha-diversity (Shannon), and beta-diversity metrics were grouped by facets of “Type” (“Soft”, “Soft-aberrant”, or “Hard”) and “Claims” (“Live”, “Probiotic”, or “No claim”) to compare compositions by kombucha type and label claim. Beta diversity was calculated using Bray–Curtis distance and visualized using MicroViz [
29]. The samples in the NMDS plot are sorted based on their pairwise similarities, with closer samples representing more similar microbial communities. The ordering is determined by minimizing the stress value, which measures the discrepancy between the pairwise distances in the plot and the original distances between the samples. Generalized UniFrac and weighted UniFrac scores were calculated using the phyloseq distance function and were subsequently used to determine the dissimilarity between samples according to their bacterial and fungal communities. For beta diversity, overall and pairwise comparisons were made with permutational multivariate analyses of variance (PERMANOVA) and were performed using adonis in vegan [
30]. For “type” comparisons, samples without NMR data were excluded from analyses.
4. Discussion
Kombucha has rapidly become a mainstream beverage over the past decade, as the emergent public awareness of gut health as a function of the gut microbiome has become a driver for consumers to make choices consistent with the “wellness lifestyle”. Central to the place of kombucha within this paradigm is the perception that it contains live, potentially probiotic microbes and that non-alcoholic styles are indeed non-alcoholic.
Within our survey, approximately one-third of retail-available non-alcoholic kombucha products exceeded the TTB’s threshold of 0.5% ABV. This is consistent with the proportion of non-compliant samples (>1% ABV) observed in a comprehensive (n = 684) sampling of kombucha products from different points of sale in British Columbia, Canada [
32]. We termed these samples “soft-aberrant” and assume that excess ethanol was due to in-package refermentation, possibly reflecting failures in cold-chain logistics [
33].
Distinct chemical profiles of hard kombucha samples suggest that products available at retail in the Pacific Northwest of the United States are intentionally alcoholic, utilizing defined production systems, rather than representing aberrant batches of soft kombucha. We found that hard kombucha contained more glycerol, and it is notable that the most prevalent fungal species associated with soft kombucha production,
Brettanomyces bruxellensis [
10,
11], produces negligible glycerol during alcoholic fermentation under anaerobiosis in minimal culture medium (5% glucose, 0.2% (NH
4)
2SO
4, 0.5% KH
2PO
4, 0.04% MgSO
4·7H
2O, and 0.1% yeast extract) [
34]. Saccharomyces cerevisiae, on the other hand, is known to balance its redox state during fermentation and manage osmotic stress by making more glycerol [
35]. While no studies have directly assessed whether these yeast species exhibit similar behavior under conditions of kombucha fermentation, we speculate that hard kombucha producers inoculate a base kombucha tea with starter cultures of
S. cerevisiae, given its widespread application in other alcoholic fermentations [
16]. The detection of erythritol only in hard kombucha samples in our survey, on the other hand, is less readily explainable. As a non-caloric sweetener, erythritol addition likely reflects attempts to appeal to health-conscious consumers [
2,
36]. Alternatively, hard kombucha producers may be taking advantage of the relative microbial stability conferred by ethanol, acidity, and low residual sugars and therefore using erythritol for organoleptic balance without a risk of continued in-package fermentation, even when stored at non-refrigeration temperatures.
We found that across soft, soft-aberrant, and hard kombucha samples, the main organic acids were acetic acid, gluconic acid, and lactic acid. We did not detect glucuronic acid, a finding in line with recent studies, which may have implications for some health claims associated with consuming kombucha [
15,
37]. However, it is important to note that our study only analyzed a limited number of commercially available kombucha brands.
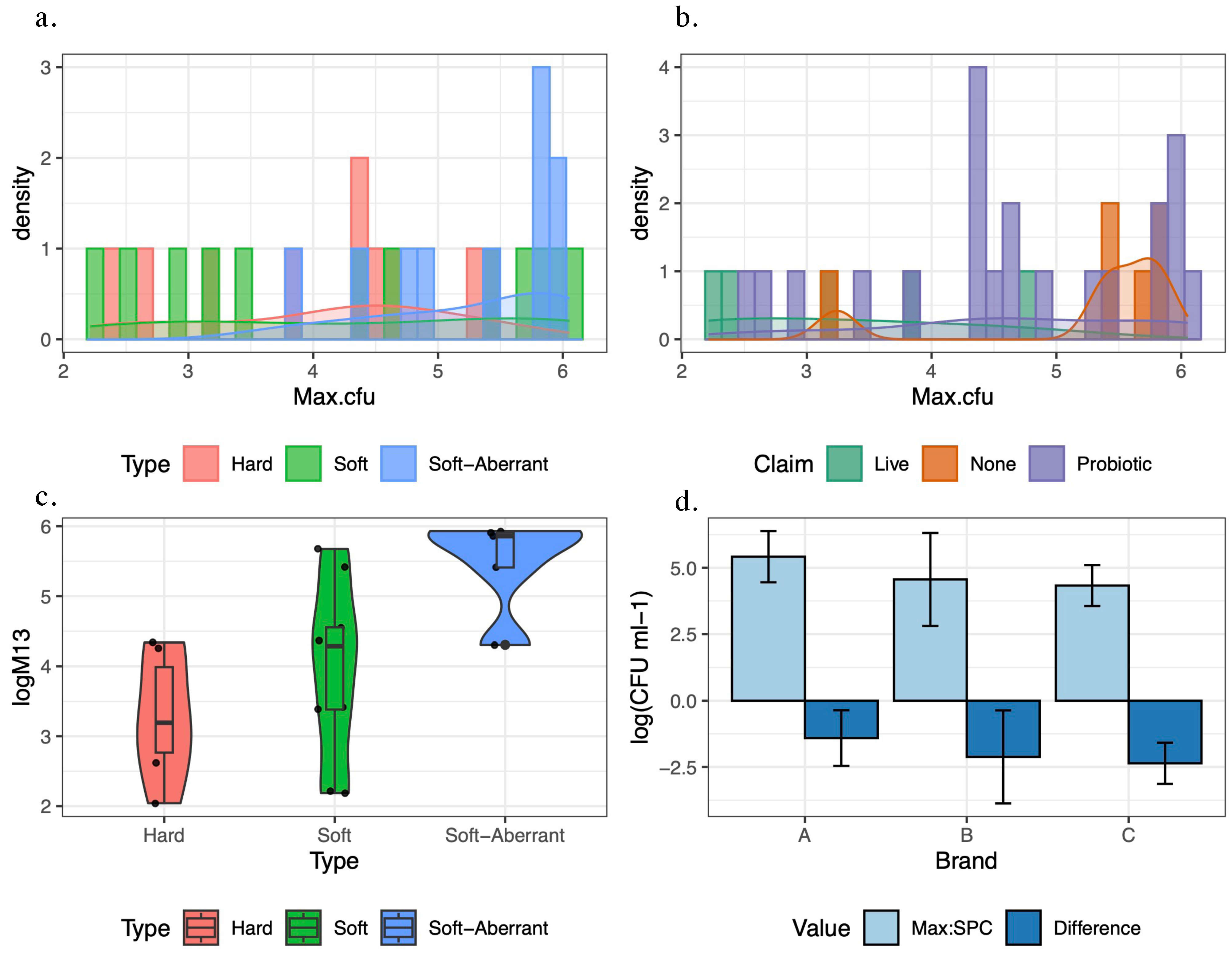
Based upon our dataset, the risk of kombucha continuing alcoholic fermentation in-package is considerable, because across all three kombucha types, the majority of products sampled contained culturable microbes. We observed a wide range of maximum CFU counts, from
to
CFU/mL, but did not observe a relationship with alcohol (kombucha type). Live microbial population sizes in our survey were lower than the
–
CFU/mL quantified using qPCR across 16 retail kombucha samples in a recent study [
15]. Our results are, however, in line with culturable microbial populations in the liquid phase during kombucha fermentation [
37,
38]. Compared with other “live” fermented foods, retail-available kombucha CFU counts span a similar range to traditional Asian fermented foods, such as miso, tempeh, and fish sauce, all of which contain fewer CFU than yogurt or other cultured dairy foods [
39].
In contrast to the lack of differences between kombucha types with regard to culturable microbial population sizes, the structures of retail-available hard, soft, and soft-aberrant kombucha microbial communities diverged. The major difference between fungal communities was that hard kombucha exhibited decreased species richness, likely due to the inoculation of
S. cerevisiae, which was most abundant in the read counts of hard kombucha. Niche engineering by
S. cerevisiae through the production of ethanol is known to cause a collapse in fungal diversity of wine fermentations, and the dominance of
S. cerevisiae in hard kombucha likely reflects its capacity to outcompete
B. bruxellensis and
B. anomalus [
40]. Bacterial communities differed according to the relative abundance of several genera, including the two most abundant:
Gluconobacter and
Bacillus. It was interesting to note that
Gluconobacter comprised more sequencing reads than
Komagataeibacter in retail-available soft kombucha, whereas
Komagataeibacter is the consensus dominant bacterial genus in the kombucha pellicle as well as in the liquid phase at the end of kombucha fermentation [
13,
14,
41]. These observations indicate that different production practices during hard kombucha fermentation are responsible for the differences in the abundance of the main taxa between hard, soft, and soft-aberrant kombucha. Therefore, there is no evidence to suggest that soft-aberrant kombucha shows an intermediate abundance of the main fungal or bacterial taxa between hard and soft types.
A large proportion of brands surveyed made a probiotic claim on their product packaging, and a subset of these listed probiotic strains within their ingredient lists with accompanying CFU numbers “at packaging”. Our data show that for three brands, represented by multiple individual products, the maximum CFU counts of retail-available samples were, on average, 2–4 log lower than the probiotic CFU counts stated on their labels. The maintenance of minimum effective CFU levels during the product’s shelf-life is a recommended criterion for the consideration of food as probiotic [
42]. It is also essential that the stated species/strains are detectable in retail-available products. Metabarcoding analyses revealed abundant reads associated with
Bacillus, particularly in soft kombucha, supporting the presence of
B. coagulans in products listing strains of this species in ingredient lists. This was not the case for probiotic
Lactobacilli or
S. cerevisiae var.
boulardii. The lack of an association between
Lactobacilli in ingredient lists with
Lactobacillus reads may reflect the poor survivability of some lactic acid bacteria in kombucha, although it should be noted that some LAB may not have poor survivability, especially those strains that are more adapted to kombucha or those that are naturally present in the pellicle. Fu et al. [
43] observed that during refrigerated storage, inoculated
Lactiplantibacillus plantarum populations decreased in size by at least 4-log. The practice of supplementing kombucha with known probiotic species/strains is one solution to meet consumer expectations of healthfulness, but shelf-life stability appears to be a critical gap that requires further research [
2]. Encapsulation strategies that protect culture viability in kombucha may be a path forward [
39,
40]. Regardless, it is worth considering whether this practice is necessary or beneficial for consumers if the purpose is solely to support probiotic labeling.
Kombucha has a long cultural history of perceived health benefits that, as already discussed, require more supporting clinical evidence. Nevertheless, this association exists and has been established for kombucha beverages containing their own microbial communities; supplementation with known probiotics may disrupt these communities and inadvertently negate associated gut-health benefits. Emerging views of fermented foods and health acknowledge that their functionality may be cumulative as part of a diet comprising other fermented products, collectively delivering transient microbes to the gut [
8]. In this context, the labeling terminology “contains live and active cultures” would be most applicable to traditionally made kombucha [
44]. Given the acidic pH range (2.5–4.0) of kombucha, it is reasonable to speculate that it contains microbial communities with greater potential to survive the gastric environment, though, to date, there is no evidence that kombucha microbes become resident in the gut microbiome. Moreover, there is a lack of evidence that food is a more effective form of probiotic delivery than supplements, though some research has focused upon the acid adaptation of probiotic strains for this purpose [
45]. Regardless, it is well established that live or even dead microorganisms, through their cellular constituents or metabolites, can stimulate the gut microbiome, modulating its composition with associated impacts on the host’s health [
46]. Therefore, further research is needed to understand the extent to which kombucha consumption meets consumer expectations, and strategies should be developed to maximize the native microbial content of finished kombucha while addressing stability issues.
Setting aside kombucha supplemented with known probiotic strains of
B. coagulans, our metabarcoding analyses imply that most kombucha products in the marketplace in the Pacific Northwest of the United States do not contain populations of genera corresponding to demonstrated species/strains with probiotic capacity, many of which belong to the lactic acid bacteria [
47]. It is possible that the two hard and two soft kombuchas exhibiting relatively high abundances of
Streptococcus reads represent products to which a known probiotic has been added but not listed with the ingredients.
Streptococcus thermophilus is a well-known starter culture used in dairy foods that has demonstrated probiotic capacity [
48] but does not feature in the kombucha SCOBY microbiome [
13]. On the other hand, six kombuchas harbored bacterial communities that were
Lactobacilli-dominant, exhibiting sequence features assigned to
L. nagelli. This species was the most abundant lactic acid bacterium detected in kombucha SCOBY, albeit a relatively minor component compared to other genera [
10]. Preliminary work on a non-kombucha isolate of this species suggests that it exhibits properties found in probiotic
Lactobacilli [
49].
As the first large survey of commercial retail-available soft and hard kombucha in the United States that evaluated both chemical and microbial compositions, our study provides a snapshot of this increasingly popular beverage. Our data point toward a known but continued challenge for the soft kombucha industry—aberrant alcohol levels that exceed regulatory requirements. Strategies to mitigate the risk of post-packaging alcohol formation, such as pasteurization, conflict with the ideology surrounding kombucha as a “live” product with potential probiotic value and are currently not widely practiced [
2], but the results of our study suggest that native kombucha microbial communities do not contain effective doses of known probiotics. Supplementing kombucha with probiotic strains and listing them as ingredients is one way to address this; indeed, we were able to detect added
B. coagulans in retail-available kombucha. The extent to which this meets consumer expectations is worthy of further research, along with strategies to maximize the native microbial content of finished kombucha while addressing stability issues, which constitutes quite a challenge.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}