1. Introduction
Amylolytic enzymes are among the most important enzymes used to manufacture food and beverages. They produce a variety of monosaccharides and oligosaccharides by acting on the O-glycosidic linkages of starch molecules [
1]. The hydrolysed products obtained from starch are extensively present in various food processes such as ethanol production, baking, and brewing, as well as in the pharmaceutical and textile industries [
2]. Although alkaline and acidic treatments, microwaving, heating, and extrusion are ways to fragment the starch glycosidic bonds to form different oligomers and monomers, enzymatic hydrolysis is the most extensively utilised methodology due to the low energy consumption and excellent product yield. Acid hydrolysis, for example, can produce undesirable colour and bitter taste, as well as highly salted solutions due to the need for pH neutralisation post-reaction [
1,
2]. Among different starch debranching enzymes, amyloglucosidase, an exo-amylase, has an extra commercial interest due to the complete conversion of starch and other related polysaccharides into D-glucose monomers, as it specifically hydrolyses α-(1 → 4) and α-(1 → 6) glycosidic linkages of starch into D-glucose [
1,
2]. Moreover, this enzyme is of great interest for use and study because of its varied food and beverage applications, such as the preparation of sweeteners; the liquefaction of insoluble starch granules present in fruit juice, especially evident in unripe fruits; the production of alcoholic beverage by increasing the fermentability and filterability of wort, facilitating the development of low carbohydrate beers and boosting alcohol yield through distilling processes; the saccharification of complex carbohydrate available in plant-based beverages, mitigating potential sensory degradation caused by heat treatment; the production of fermentable carbohydrates, thereby fostering the production of fermented plant-based beverages; and the treatment of starch processing wastewater [
3,
4,
5,
6,
7].
The biotransformation of starch-based materials into simple sugars is a valuable approach for generating cost-effective and sustainable industrial processes. It also enhances the digestibility of starchy foods and drinks, improves the texture and mouthfeel, and prolongs the shelf-life by reducing the presence of starch, which can lead to spoilage. In this context, biocatalytic compounds with exceptional properties, including high catalytic activity and selectivity, emerge as viable options for various practical applications. However, some difficulties have appeared as significant obstacles in their industrial usage, such as their low thermal and chemical stabilities, narrow pH range, catalytic activity limited under certain conditions, the necessity of stopping the reaction by high temperature, and challenges in recovery and reusability of the bioactive compounds [
8]. Meanwhile, immobilisation is a practical approach to minimise these issues by facilitating control and stopping hydrolysis reactions, improving enzyme stability, and reusing. This technique physically limits or localises enzymes in a specific region while keeping their catalytic capabilities. Furthermore, it allows the utilisation of enzymes in optimal micro-environments and conditions and improves their properties by tailoring and modifying a variety of catalytic features: activity specificity, selectivity, and stability under a variety of pH and temperature conditions, recyclability over multiple catalytic cycles, and kinetic properties [
9,
10]. Moreover, stable immobilised biocatalyst systems simplify enzyme separation from the reaction medium, diminish or completely circumvent potential product contamination, and enable continuous operations and the application of enzymes in various reactors [
11,
12,
13].
Due to these factors, extensive research efforts have been dedicated to immobilising amyloglucosidase through various techniques and a wide range of matrices. These methods include adsorption onto activated charcoal [
14], nonporous polystyrene/poly (sodium styrene sulfonate) microspheres [
15], and bone powder [
16]. The covalent attachment has also been explored using glutaraldehyde-activated chitosan beads [
8], glutaraldehyde-activated poly(o-toluidine) support [
13], glutaraldehyde-activated active carbon [
5], and glutaraldehyde-activated magnetic nanoparticles [
17,
18]. Other approaches involved entrapping the enzyme into beads made of alginate [
8], agar–agar [
1], or agarose [
2] or creating a crosslinked enzyme aggregate by treating amyloglucosidase with glutaraldehyde [
19].
Among many supports for enzyme immobilisation, chitosan is highly appealing due to its favourable properties. This organic biopolymer has been extensively utilised in various food applications in recent years, and its unique biological and chemical characteristics have sparked interest in the field of immobilisation. Chitosan is a linear polysaccharide made by the deacetylation of chitin. It is a cationic, inert, non-toxic, biodegradable, and versatile low-cost biocompatible polymer that forms gels in various geometrical configurations, including beads, membranes, nanoparticles, powder, and fibres [
8,
20,
21]. It has two modifiable groups (amino and hydroxyl) in an aqueous acidic solution that chemical treatments can easily amend. In addition, the high porosity, high hydrophilicity, broad adhesion area, and low mass transfer resistance to the enzyme are all factors that contribute to chitosan’s appeal as an enzyme immobilisation carrier material. It has also been demonstrated to be adequate support for immobilising various enzymes, including glycosidases, proteases, glucose oxidases, and others [
20,
22].
Despite the advantageous characteristics that make chitosan considerable and beneficial for the immobilisation of proteins and cells, its extensive application and excellent quality were studied and explored mainly as support for adsorption and covalent attachment systems of enzymes [
23,
24,
25,
26]. In addition, the commercial use of chitosan hydrogel beads formed by alkaline precipitation is limited due to their low mechanical strength and low acid stability. This low acidic stability could result in poor functionality of the gels to efficiently entrap enzymes that operate in acidic environments, such as the amyloglucosidase under study here. Therefore, a promising approach for producing more resilient and robust chitosan hydrogel beads is using sodium dodecyl sulfate (SDS) [
27,
28]. Studies have demonstrated that chitosan and SDS can interact to form chitosan-SDS hydrogel capsules with different properties, making them an attractive alternative for drug delivery, cosmetics, and food applications [
29]. These capsules have also shown potential for wastewater treatment and have been used particularly in the adsorption of coloured compounds from textile dyeing wastewater [
28,
30].
The literature suggests that chitosan can interact with anionic surfactants such as SDS to form coacervates with various structures and properties via a simple coacervation phenomenon [
29,
31]. A spontaneous liquid/liquid phase separation forms a dense coacervate phase and a dilute equilibrium phase. The coacervates can be created even at surfactant concentrations below the critical micelle concentration and may be stabilised by electrostatic attraction between oppositely charged molecules [
31,
32,
33]. Henceforth, a negatively charged surfactant, sodium dodecyl sulfate (SDS), was utilised to form strong bonds with cationic chitosan, developing insoluble coacervates. Previous research indicates that the application of anionic surfactant to create chitosan hydrogel capsules for enzyme immobilisation has yet to be documented. Therefore, this study aimed to develop a novel chitosan-SDS hydrogel to encapsulate amyloglucosidase. The morphological characteristic of the capsules was evaluated in terms of morphology and size. The enzymatic activity of free and immobilised amyloglucosidase was investigated through the glucose production performance by gelatinised corn starch and maltose hydrolysis. In addition, studies of immobilisation yield, enzyme efficiency, storage stability, feedback inhibition and kinetic properties were performed.
3. Results and Discussion
3.1. Morphological Characteristics
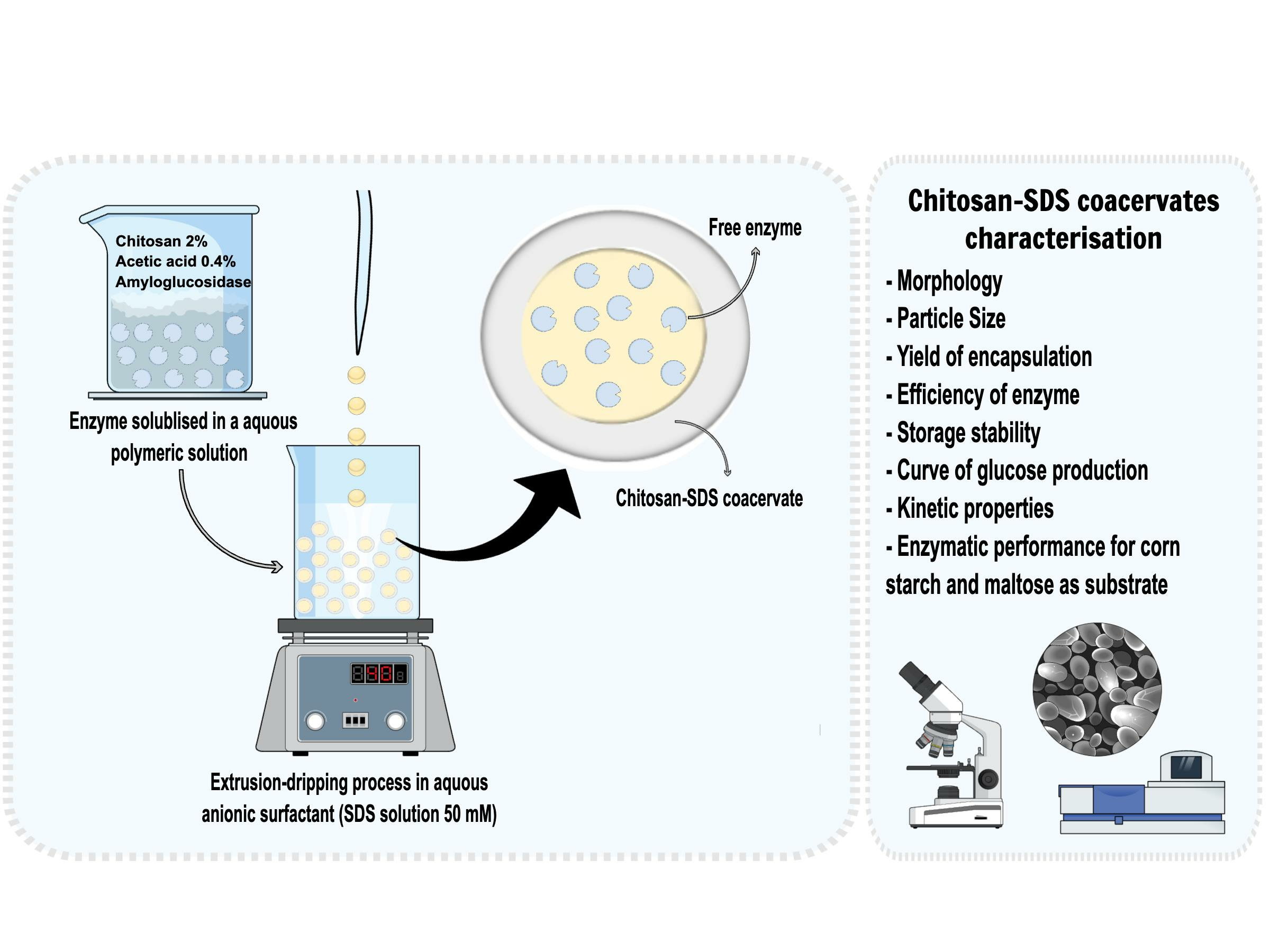
An image of the chitosan-SDS beads containing amyloglucosidase is shown in
Figure 1. The capsule-like beads are characterised by a thick outer layer membrane and a more transparent core in the centre. Upon further observation, the capsules were physically cut open with the aid of a scalpel and were found to be made of a liquid core surrounded by a gel-like membrane.
The average diameter of the capsule and core, as well as the membrane size, were evaluated. The capsule diameters were 3.17 ± 0.03 mm, with a membrane size of 0.55 ± 0.02 mm and a core diameter of 2.62 ± 0.05 mm.
The large mean particle size results from the orifice through which the chitosan solution was extruded, a 200 μL pipette tip, which resulted in large chitosan pre-gel solution drops (drop weight of 19 mg ± 0.04 corresponding to drop diameter sizes of 3.31 ± 0.01 mm). According to Takka and Gurel [
37], the polyethene tubing nozzle is an important parameter leading to the formation of large particle sizes. In addition, beads with a diameter greater than 1 mm are usually prepared using a syringe with a needle or a pipette tip.
The process of forming these beads involved dropwise extrusion of 2% chitosan into a neutral 50 mM SDS solution. As chitosan is a positively charged polymer soluble in an aqueous solution at a pH lower than 6.5, it readily interacted with the anionic surfactant SDS through electrostatic attraction. The interaction between both compounds resulted in the formation of an insoluble coacervate with self-supporting structures and spherical reservoir-type capsules.
The large size of beads can be considered an advantage regarding recycling enzymes and controlling enzymatic reactions. Due to their size, a simple filtration process can effectively remove the insoluble enzyme from the environment, facilitating enzyme recovery and reaction termination.
3.2. Curve of Glucose Production for Gelatinised Corn Starch
The curve of glucose production during the hydrolysis of gelatinised corn starch solution (5 g/L) at 60 °C was plotted against time, as shown in
Figure 2. The study was performed for 300 min using free and encapsulated enzymes with an overall concentration of 240 mg enzyme per litre of substrate solution. As observed, the glucose production curve in the case of the free enzyme was composed of an initial phase where glucose production increased linearly with hydrolysis time, followed by a plateau where the glucose production remained constant. In the case of the encapsulated enzyme, a lag phase is also seen in the first 10 min of the reaction, where the glucose production is prolonged and only reaches a maximum between 30 and 90 min. The free enzyme displayed a steep slope at the beginning of the hydrolysis, indicating a high initial reaction velocity (V
0). This high initial reaction velocity leads to the near-final conversion of starch into glucose within the first 5 min of the reaction, in line with previously published data by Selmi et al. [
38]. On the other hand, the encapsulated enzyme displayed a much slower slope, suggesting that the encapsulation process has vastly reduced the enzymatic reaction velocity.
The reduction in the catalytic performance was also observed in another study in that of immobilised amyloglucosidase by entrapment in an agar–agar matrix [
1]. The authors attributed the reduction to the amyloglucosidase being confined within the microenvironment of agar–agar, which also hindered the penetration of soluble potato starch into the matrix and created diffusional resistance or steric hindrance, further contributing to the lower enzymatic activity.
Figure 2 also revealed different conversion rates of corn starch into glucose between the free and encapsulated enzymes. The amyloglucosidase in free form achieved a plateau of glucose production at 4068 mg/L from an initial starch concentration of 5000 mg/L or a conversion rate of 81 ± 0.43% of the total glucose. In contrast, the encapsulated enzyme achieved a conversion rate of 67 ± 3.7%. The lower-than-expected conversion rate for free enzymes (amyloglucosidases are known as amylases that completely hydrolyse starch and other related polysaccharides into D-glucose monomers) suggested a potential end-product inhibition for both free and encapsulated enzymes, possibly due to excess product in the reaction medium. The lower substrate conversion rate for encapsulated enzymes instigated the necessity of understanding the immobilisation yield and enzyme efficiency of the encapsulated amyloglucosidase. Therefore, studies were performed to investigate feedback inhibition, immobilisation yield and enzyme efficiency.
The lag phase, observed in
Figure 2, at the beginning of the catalytic reaction has also raised the hypothesis that the gel-like structure or the chitosan-SDS coacervate restricted the diffusion of the large corn starch substrate molecules into the capsule and slowed the release of glucose product back into the solution. In contrast to the free enzyme, which exhibited a rapid increase in the glucose concentration from the beginning of the hydrolysis, gelatinised corn starch showed minimal glucose production in the first 10 min of catalytic activity, followed by a sharp increase that eventually reached a linear trend. Therefore, to address this concern, a hydrolysis study was conducted using a smaller substrate, such as maltose, while maintaining the same immobilisation system. Additionally, the kinetic properties, including the Michaelis–Menten constant and the maximum reaction velocity, were evaluated to have further information about the enzymatic performance of chitosan-SDS coacervates containing amyloglucosidase using different substrates.
3.3. Yield of Immobilisation and Enzyme Efficiency
The effectiveness of the encapsulation process was evaluated through the immobilisation yield and enzyme efficiency. The method introduced by Sheldon and van Pelt [
36] for immobilisation yield was adopted. It involved measuring the encapsulated enzyme’s activity (through the total enzymatic activity that was mechanically released from the capsules) compared to the original activity of the free enzyme before immobilisation (see rate of Glucose production in both cases in
Supplementary Materials Figure S1). In order to assess enzyme efficiency, the total glucose produced using the encapsulated enzyme was compared to the total glucose produced in its free form. The immobilisation yield was determined to be 71.3 ± 2.6%, indicating the percentage of successfully encapsulated enzymes’ activity, and the enzyme efficiency of 83.2 ± 0.75%, illustrating the capability of the immobilised enzyme to convert starch into glucose when compared to its free form.
The chitosan-SDS capsules containing amyloglucosidase demonstrated a high immobilisation yield of 71.3 ± 2.6%, meaning efficacious preservation of enzyme activity through the simple coacervation process. This method successfully surrounded numerous active enzymes with a protective wall material due to favourable immobilisation conditions that facilitated the formation of self-supported and stable chitosan-SDS capsules. The achieved immobilisation yield for amyloglucosidase using this approach was consistent with the results reported in the literature. For instance, Pervez, Aman and Ul Qader [
8] achieved a 66% immobilisation yield when they entrapped amyloglucosidase in alginate beads, while amyloglucosidase immobilised into agarose (40 g/L) reached a 78% yield in another study [
2].
Regarding enzyme efficiency, the encapsulated enzymes’ ability to convert corn starch into glucose to an extent similar to the free enzyme was determined to be 83.2 ± 0.75%. This result can be attributed to the gentle encapsulation process, which did not require harsh conditions such as a prolonged curation period, extreme pH or high temperatures that could denature or inactivate the enzymes. The encapsulation process involved mixing the amyloglucosidase (with an optimum pH of around 4.8) with a chitosan solution at pH 5.87 and then dripping it into an SDS solution at pH 7.0, resulting in the immediate formation of the chitosan-SDS membrane encapsulating the enzyme. The curation process took only 30 min, a significantly shorter period compared to studies using crosslinking agents, such as genipin or glutaraldehyde, which require hours and even days of curation to obtain firm and stable beads [
8,
20]. The curation period and pH condition did not contribute to denaturing or inactivating the biocatalytic compound. The chitosan-SDS coacervate formed a membrane that preserved enzymatic activity and allowed substrate catalysis.
Furthermore, the glucose production curve for the released encapsulated enzyme exhibited similar behaviour to the curve obtained for the free enzyme (shown in the
Supplementary Material in Figure S1). During the initial phase, a linear correlation between glucose produced and hydrolysis time was seen, with the slope of V
0 determined to be 706 ± 18 mg L
−1 min
−1 glucose for the free enzyme and 503.8 ± 31 mg L
−1 min
−1 glucose for the released enzyme. These results suggest that the lower initial reaction velocity observed for the released enzyme, compared to the free form, may be attributed to the fraction of enzyme that could not be released successfully, probably the enzyme located in the coacervate gel for which the activity was impossible to measure. However, the significantly high V
0 and enzymatic activity after the release of the encapsulated enzyme imply that most of the encapsulated enzyme (71%) was in a free and active form inside the liquid core of the capsules, allowing for preserved mobility and regular enzymatic activity.
3.4. Study of Feedback Inhibition
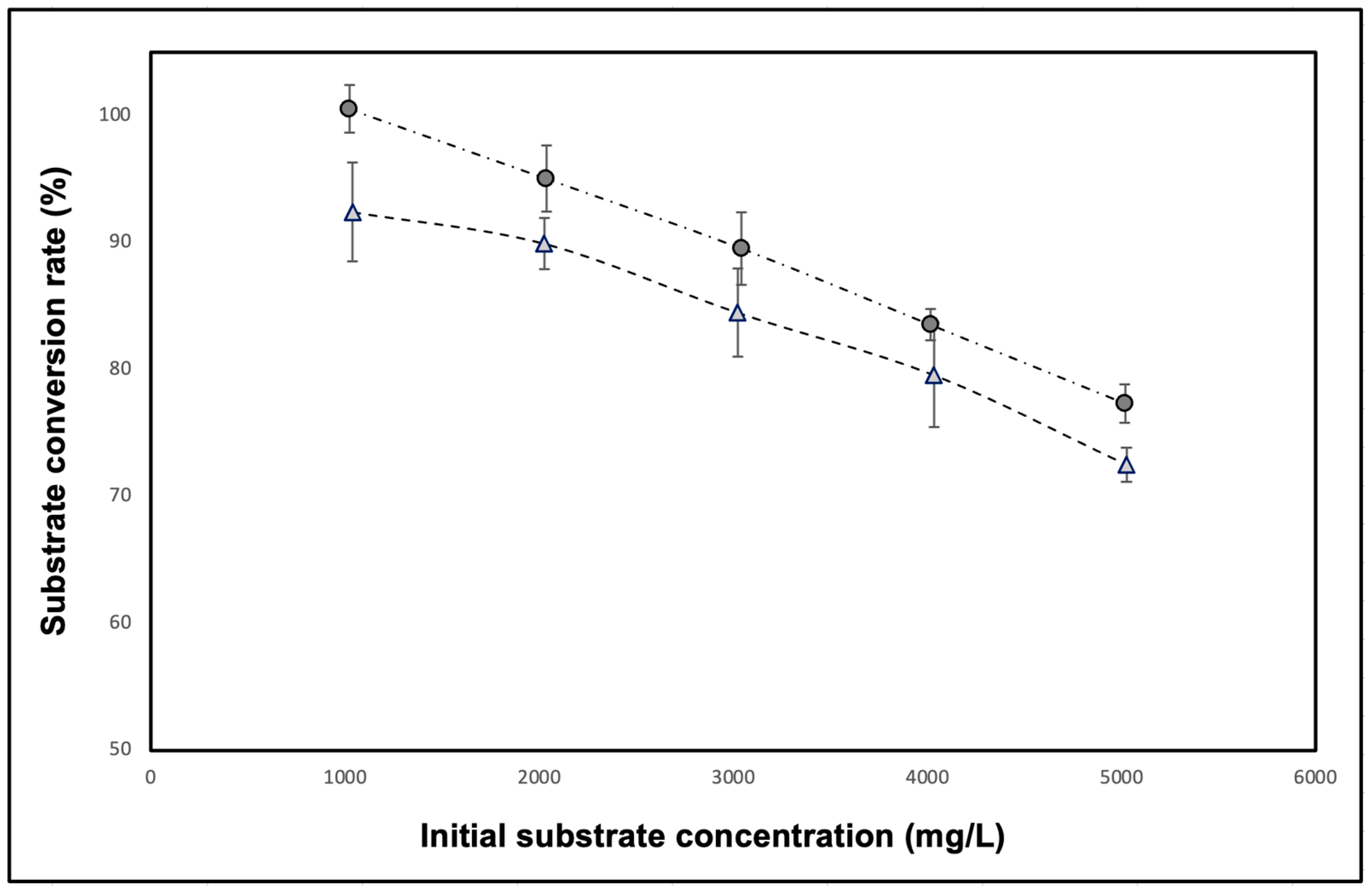
The feedback inhibition plot for free and encapsulated amyloglucosidase is displayed in
Figure 3. The graph illustrates the conversion rate of corn starch into glucose at various initial corn starch concentrations in the reaction medium. Both free and encapsulated enzymes showed consistently decreasing substrate conversion rates as the initial substrate concentration increased. When using a concentration of 1 g/L corn starch, 100% substrate was converted by the free enzyme, whereas the encapsulated enzyme hydrolysed only 92% of the substrate. At initial gelatinised corn starch concentrations of 2, 3, 4, and 5 g/L, the conversion rates for free enzyme were 95%, 91%, 82%, and 79%, respectively, while encapsulated enzyme achieved 90%, 84%, 80%, and 73%, respectively. Notably, the encapsulated amyloglucosidase exhibited a lower conversion rate than the free enzyme, roughly 10% or less, at the same concentrations. This small but significant difference in substrate conversion rate between free and encapsulated enzymes confirms that the encapsulation process reduces the enzyme’s performance marginally in terms of the total amount of glucose produced.
Despite the disparity and significant difference in conversion rates between the free and encapsulated enzymes, the near linear decrease in the plot indicates the presence and impact of feedback inhibition in both scenarios, suggesting that feedback inhibition remains a robust regulatory mechanism for amyloglucosidase, regardless of encapsulation. The feedback inhibition mechanism involves binding glucose, a product of the enzyme’s hydrolysis of starch, to the enzyme at an allosteric site, reducing enzyme activity [
39]. Then, the accumulation of glucose as a reaction product likely inhibited the enzyme from binding to a new substrate and generating more product, thus contributing to the observed conversion rates in both cases. The results also reveal that the substrate has not been fully consumed, and products have not been completely formed, particularly with increasing substrate concentration, due to end-products build-up in the reaction medium. These observations imply that the chitosan-SDS encapsulation system did not prevent the regulatory mechanism of amyloglucosidase.
Although the reduced conversion rate and maintained feedback inhibition, the encapsulation of amyloglucosidase offers potential advantages such as enzyme stability, protection, and controlled product release. It also circumvents potential enzyme product contamination. These make it a valuable approach for specific applications in the food and beverage industry. The feedback inhibition outcomes suggest that this enzyme system could be conveniently applied in a continuous reaction treatment, which operates based on a constant flow of substrate and product, where the encapsulated enzyme works within an enzymatic reactor with no interruption or need for enzyme removal. Such continuous flow applications promise to enhance process efficiency and scalability in various industries and eliminate issues regarding regulatory mechanisms, such as feedback inhibition.
3.5. Comparison between Maltose and Gelatinised Corn Starch as Substrates on Glucose Production and Enzyme Performance
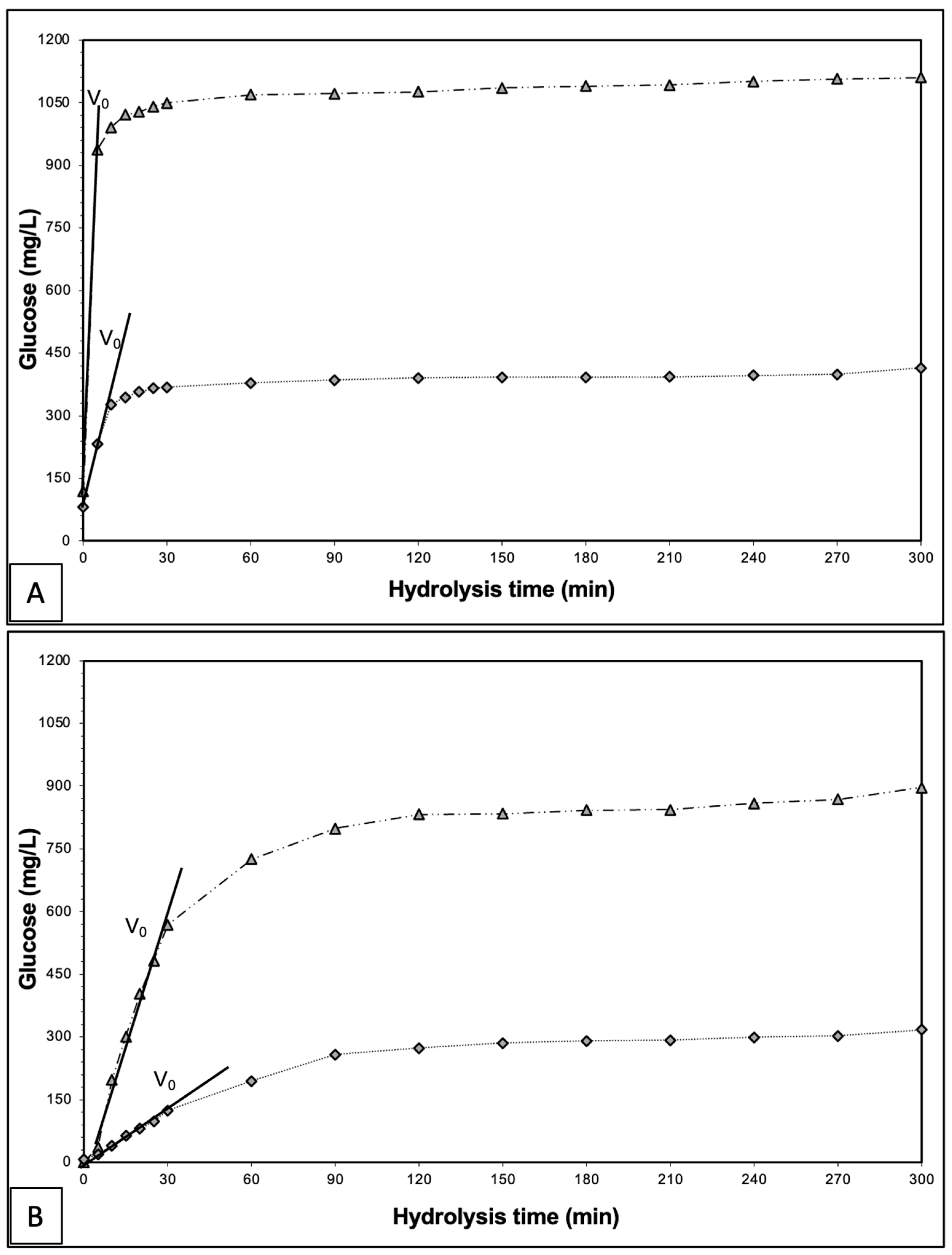
A comparison between glucose production curves resulting from the 300 min of hydrolysis of maltose and gelatinised corn starch solutions, each at an initial concentration of 1 g/L and using free and encapsulated enzymes, each at an overall concentration of 240 mg/L, is shown in
Figure 4. The investigation focused on understanding the detected lag phase in glucose production when encapsulated enzymes were used (
Figure 2). Corroborating with previous results, as observed in
Figure 4A, the free enzymes led to an initial hydrolysis phase with a steep slope for both substrates, reaching a plateau within 30 min. In the case of the encapsulated enzyme (
Figure 4B), a more gradual increase in glucose production over time was seen for both substrates. However, maltose hydrolysis displayed a linear curve from the start, while gelatinised corn starch exhibited a lag phase with minimal glucose production in the first 5 min, followed by a sharp increase, eventually reaching a plateau for both substrates. The free enzyme displayed a steep slope at the beginning of the hydrolysis, indicating a high initial reaction velocity (V
0) of 567 ± 23 and 24.5 ± 2 mg L
−1 min
−1 glucose for corn starch and maltose catalysis, respectively. In contrast, the encapsulated enzyme displayed a much slower V
0 slope of 19.2 ± 1 and 4.8 ± 0.8 mg L
−1 min
−1 glucose for gelatinised corn starch and maltose hydrolysis, respectively. These results confirmed that the encapsulation had substantially reduced the enzymatic reaction rate, much more so in the case of starch than in the case of maltose. In addition, the impact of encapsulation was also observed in the enzyme’s capability to convert substrate into glucose. The encapsulated enzyme presented better efficiency in converting corn starch than maltose, with enzyme efficiency equal to 81.4 ± 2% and 76.5 ± 1%, respectively.
The lag phase seen in
Figure 4B, exclusively during the hydrolysis of gelatinised corn starch, may indicate that the chitosan-SDS membrane, which has proven highly effective in stopping the enzyme from leaching out yet is allowing diffusion of substrate and products in and out of the beads, significantly hindered corn starch substrate diffusion and glucose release from the capsule. It resulted in an apparent lag phase in glucose production, which could be attributed to corn starch’s large molecule size and structure. As demonstrated in
Figure 1, the chitosan-SDS coacervate formed a protective barrier around the enzyme, and although chitosan has high porosity, the corn starch might have faced difficulties entering the capsules, consequently accessing the enzyme’s active site within the beads. On the other hand, this diffusional hindrance was comparatively less pronounced for maltose, as evidenced by the linear curve observed right from the beginning of the enzymatic reaction. This implies that maltose encountered fewer diffusion restrictions than gelatinised corn starch, likely due to its lower molecular weight and smaller size. However, despite maltose having a smaller molecular weight benefiting its diffusional characteristics, its initial reaction velocity and enzyme efficiency were lower than those of gelatinised corn starch, with significant differences confirmed by an independent
t-test at a significance level of 0.05. The reasons behind these findings could be multifaceted, including the enzymatic preference for corn starch over maltose, the inhibitory effect of maltose as a reducing sugar on enzymatic activity due to the previously mentioned regulatory mechanism, and the limited range of glucose production.
Thus, the comparison of glucose production curves during the hydrolysis of maltose and gelatinised corn starch using free and encapsulated enzymes confirmed that free enzymes exhibited a rapid initial hydrolysis phase with a shorter time to reach plateaus, whereas encapsulated enzymes led to a gradual increase in glucose production over time, regardless of the substrate. The results revealed that the chitosan-SDS membrane somewhat hindered the diffusion of substrate and glucose release for corn starch hydrolysis, confirming that a simpler substrate, such as the maltose, presented less diffusional difficulties. In addition, the findings suggested an amyloglucosidase substrate preference for corn starch over that of maltose, although maltose molecular structure and size resulted in favourable diffusional aspects. These outcomes emphasise the importance of considering enzyme form and substrate characteristics in biocatalyst applications, with implications for optimising enzyme delivery systems in various industries.
3.6. Determination of Kinetic Properties (KM and Vmax) Using Gelatinised Corn Starch and Maltose as Substrate Solutions
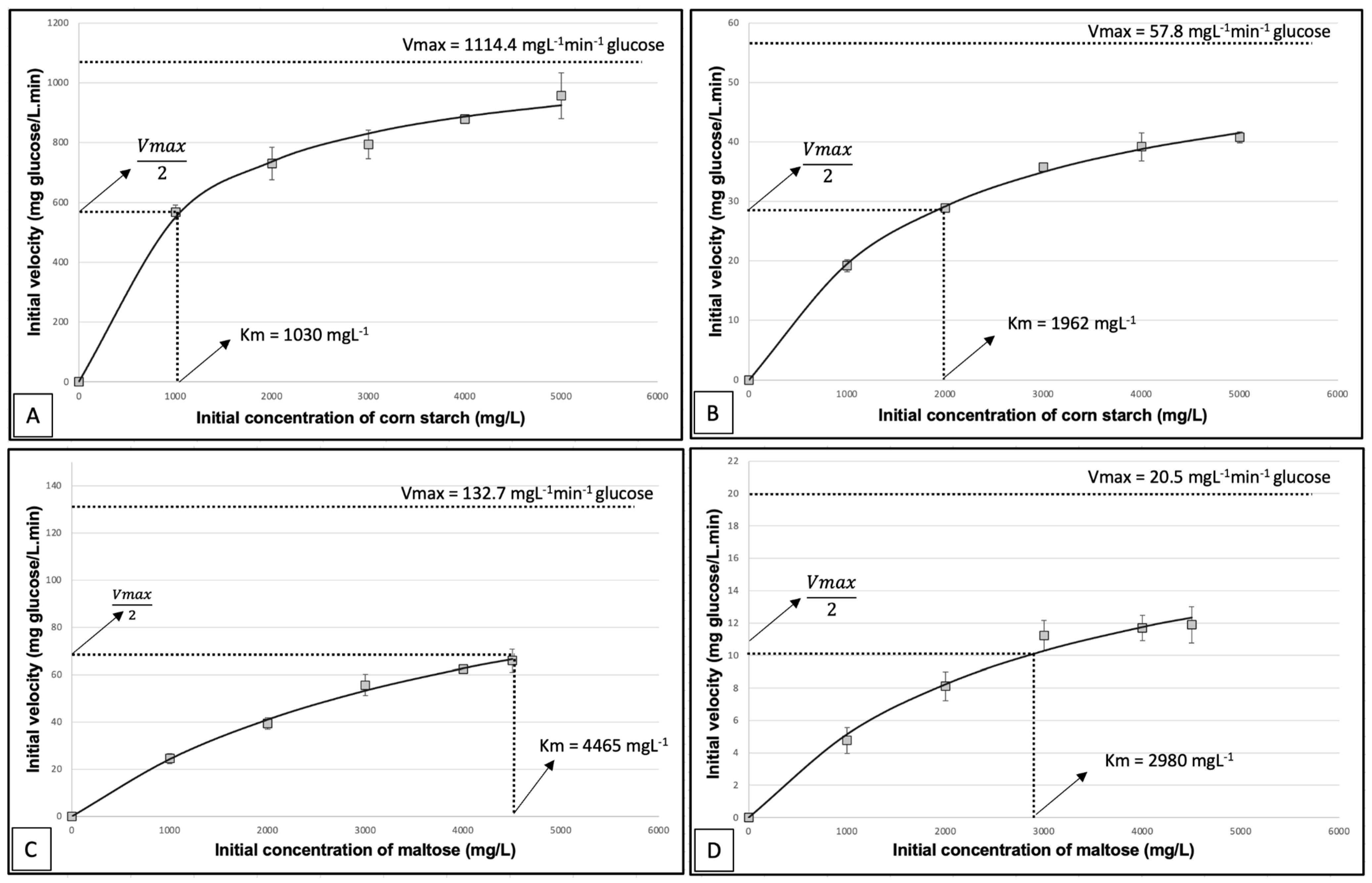
Michaelis–Menten plots, displayed in
Figure 5, exhibit the initial enzymatic velocity (V
0) of free and encapsulated amyloglucosidase across a range of initial substrate concentrations involving gelatinised corn starch and maltose. The curves were plotted, fitted, and adjusted through a non-linear fit by Solver. The Michaelis–Menten equation allowed direct determination of the crucial kinetic parameters, such as V
max (maximum reaction velocity) and K
M (Michaelis–Menten constant), in contrast to the double reciprocal method (Lineweaver-Burk plot), which necessitates additional steps such as extrapolating lines or calculating slopes. Such other steps make the double reciprocal method more susceptible to errors and uncertainties. Moreover, the double reciprocal method requires that substrate concentrations be equally spaced and data points be evenly distributed for accurate and precise interpretation [
40]. In the case of gelatinised corn starch as the substrate, the free enzyme exhibited a V
max of 1114 mg/L min
−1 glucose, whereas the encapsulated enzyme displayed a notably lower V
max of 57.8 mg/L min
−1 glucose. For maltose, the V
max for the free enzyme was 132.7 mg/L min
−1 glucose, while the encapsulated enzyme showed a V
max of 20.5 mg/L min
−1 glucose. Remarkably, encapsulation led to an increase in the K
M value from 1030 mg/L to 1962 mg/L in the case of gelatinised corn starch. Conversely, the K
M value decreased from 4465 mg/L to 2980 g/L in the case of maltose.
The observed reduction in the maximum velocity (V
max) for both substrates and the decrease in enzyme-substrate affinity (K
M) for corn starch suggests that encapsulation limited substrate accessibility to the enzyme’s active site. The creation of a rigid and compact structure during encapsulation acts as a steric barrier, causing mass transfer resistance and impeding substrate diffusion to the immobilised enzyme’s active sites, leading to lower catalytic activity compared to the free enzyme [
41]. The interaction between chitosan and SDS likely formed a membrane, challenging corn starch diffusion through the capsule and restricting its access to the enzyme’s active site, as observed through the lag phase in corn starch hydrolysis (
Figure 4B). In addition, although the overall concentration of the enzyme utilised in the hydrolysis processes was the same (240 mg/L), in the case of the encapsulated enzyme, this concentration was available in a small volume of capsules, which represented a nominal concentration of (24,000 mg/L). It might have affected the enzymatic performance and its kinetic properties. The high concentration of enzyme molecules present within the capsule volume might have interfered with the diffusional effect of both substrate and product [
42]. The heightened enzyme collision rate could also lead to undesirable intermolecular interactions and steric hindrance, potentially inhibiting enzymatic activity and reducing reaction rate [
43]. These findings align with previous literature that also observed a reduction in V
max and an increase in K
M after entrapping different types of enzymes, which might be caused by substrate mass transfer resistance and increased relative concentration of enzymes within the beads [
44,
45,
46,
47].
Looking at the effect of enzymatic encapsulation on the hydrolysis parameters for each substrate, maltose seems to be less affected than the substantially larger starch molecules. This is because maltose can diffuse more easily through the capsule membrane and reach the enzymatic active site compared to corn starch (with no lag phase apparent, as demonstrated in
Figure 4B). Encapsulation should also contribute to high local substrate concentrations around the enzyme, enhancing enzyme-substrate interaction and augmenting affinity for maltose. Similar trends were observed by de Oliveira et al. [
48] and Jadhav and Singhal [
49] when they entrapped pectinase, α-amylase and glucosidase into alginate beads. They also noticed a decrease in the Michaelis–Menten constant values. They attribute this decrease to the internal diffusion of the substrate within the alginate beads, which could have limited the substrate concentration around the gel due to the large substrate molecules having difficulty diffusing rapidly into the calcium alginate matrix. Interestingly, corn starch still exhibits superior kinetic parameters compared to maltose (higher V
max and lower K
M values for corn starch than maltose for both free and encapsulated enzymes). This difference can be ascribed to amyloglucosidase preference for corn starch, as its structural composition provides multiple binding sites [
50] specifically adapted for efficient enzyme activity. Additionally, the lower reaction velocity observed with maltose can be attributed to its simpler structure than starch. Amyloglucosidase enzymes are optimised to efficiently process complex molecules such as starch, resulting in lower reaction rates with simpler substrates such as maltose.
Comparing the kinetic parameters by hydrolysing gelatinised corn starch using free and chitosan-SDS amyloglucosidase obtained in this study, it was observed strong correlation with existing literature. Studies by Rani et al. [
51] and Rani, Das and Satyanarayana [
14] characterised amyloglucosidase bound to chitin and adsorbed on activated charcoal, respectively. They reported an increase in the Michaelis–Menten constant (K
M) after the immobilisation process, confirming the reduction in the affinity between the enzyme and substrate upon immobilisation of amyloglucosidase on the surfaces of chitin or activated charcoal supports. However, in this study, the maximum reaction velocity (V
max) was much smaller than their findings, suggesting that substrate preparation, temperature, and pH significantly influence enzyme performance and the overall reaction rate. Similarly, Aslan, Sharif and Sahin [
5], on maltodextrin hydrolysis, observed a reduction in K
M values after the immobilisation process, in line with results obtained for maltose hydrolysis.
3.7. Storage Stability
Table 1 presents the comprehensive results from a 28-day storage stability study on amyloglucosidase-chitosan-SDS coacervates. Stored at 5 °C in a dilute glucose solution of 1 g/L, samples were analysed at regular intervals (7, 14, 21, and 28 days) to assess their enzymatic activity, capsule weight, and shrinkage. The coacervates of chitosan-SDS demonstrated preservation of 76.2 ± 2.7% of the initial amyloglucosidase activity after the 28-day storage period. During the first 7 days of storage, catalytic activity significantly declined from 100% to 85.3 ± 3.9%, with a substantial mass loss of 9.7% (
Table 1). Statistical analysis using one-way repeated ANOVA and the Bonferroni post hoc test revealed a notable difference in residual catalytic activity within the initial week of storage (
p < 0.05). Subsequently, no significant variations were observed in enzymatic activity or capsule weight. These findings suggest that the storage period primarily impacted enzymatic activity during the initial week of storage. The observed decrease of 11.1% in capsule weight and the decline in hydrolytic activity over the entire 28-day storage period may be attributed to the potential release of trapped enzymes within the chitosan-SDS wall or the leakage of enzymes and chitosan solution, which were physically enclosed by the chitosan-SDS membrane. These effects could be attributed to the high porosity of chitosan and the absence of a crosslinking agent that irreversibly binds the enzyme and capsule together.
Pervez, Aman and Ul Qader [
8] also observed a leaching of amyloglucosidase during the storage of alginate-amyloglucosidase beads at 4 °C. Entrapped amyloglucosidase retained only 30% of amyloglucosidase enzymatic activity after 90 days of immobilisation. In contrast, the authors evaluated the storage stability of the enzyme immobilised onto chitosan beads using glutaraldehyde as a crosslinking agent. They noticed enzymatic activity retention of 70% after the same storage period. They reported that chitosan-glutaraldehyde-based matrices effectively preserved the enzyme’s tertiary structure, preventing undesired conformational changes and enzyme release from the support. In addition, when monitoring the leaching of amyloglucosidase for both systems for 30 days, no enzyme leakage was observed for the chitosan-glutaraldehyde beads [
8]. Immobilised amyloglucosidase in an agar–agar matrix through entrapment retained around 57% of its initial activity after 60 days of storage at 4 °C [
1]. Although the study also indicated a leaching of the enzyme, it demonstrated that immobilised amyloglucosidase is more stable than the soluble enzyme during storage in the same conditions.
Therefore, the storage stability study of amyloglucosidase-chitosan-SDS coacervates demonstrated their effectiveness in preserving enzymatic activity over a 28-day storage period. Although a significant decrease in activity was observed within the first 7 days, subsequent periods showed no further variations. The observed capsule weight decreases and decline in enzymatic activity over time may be attributed to the release of trapped enzymes or chitosan solution due to the porous nature of chitosan and the lack of a crosslinking agent. These findings highlight the potential of chitosan-based encapsulation strategies for enzyme preservation during storage, offering promising applications in various industries. Nonetheless, further optimisation in encapsulation techniques would enhance long-term stability for practical use.
4. Conclusions
This study successfully encapsulated amyloglucosidase within chitosan-SDS coacervates, preserving the enzyme’s mobility and activity with an immobilisation yield of approximately 71%. The enzyme was found to be mainly free inside the liquid core of the capsule, with little or no evidence of an entrapped enzyme inside the coacervate gel. The encapsulated enzyme effectively converted gelatinised corn starch into glucose across different concentrations (ranging from 1 g/L to 5 g/L), with varying conversion rates of 92.42 ± 3.90% to 72.51 ± 1.32% from lower to higher concentrations, respectively. The encapsulation process did alter the kinetic properties of the enzyme, leading to a reduction in the maximum reaction velocity and an increase in the Michaelis–Menten constant. In addition, it resulted in a lag phase in the glucose production curve attributed to limitations imposed by the chitosan-SDS membrane due to steric hindrance that limited the diffusion of corn starch into the capsule. However, these changes can benefit this enzyme’s food and technological applicability. As a high-catalytic enzyme, slowing down the reaction velocity can offer better control, enabling the possibility of stopping the reaction at the desired glucose quantity. This feature enhances the applicability of amyloglucosidase and facilitates stopping the enzymatic reaction by a simple separation step. Furthermore, the feedback inhibition study indicated that the encapsulated amyloglucosidase might be suitable for continuous reaction treatment applications, as it supports prolonged catalytic reactions without being hindered by high concentrations of the reaction product in the environment, which makes the immobilisation system an interesting alternative for beverage production processes. Overall, this study provides valuable insights into the encapsulation of amyloglucosidase using chitosan-SDS coacervates, advancing enzyme immobilisation techniques with potential applications in beverage production, preferably for small molecular weight substrates such as maltose or lactose, for example. The mild and food-grade chitosan and SDS encapsulation system present an attractive and promising alternative for drug delivery, cosmetics, and food-related applications. However, further improvements and optimisations are still necessary for their use in enzyme immobilisation systems. Additionally, exploring the amyloglucosidase-chitosan-SDS capsule application in continuous operational processes could yield valuable knowledge for practical use.











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}