A Collection of 13 Archaeal and 46 Bacterial Genomes Reconstructed from Marine Metagenomes Derived from the North Sea


Abstract
:1. Summary
2. Data Description
2.1. Archaeal Metagenome-Assembled Genomes
2.2. Bacterial Metagenome-Assembled Genomes
3. Methods
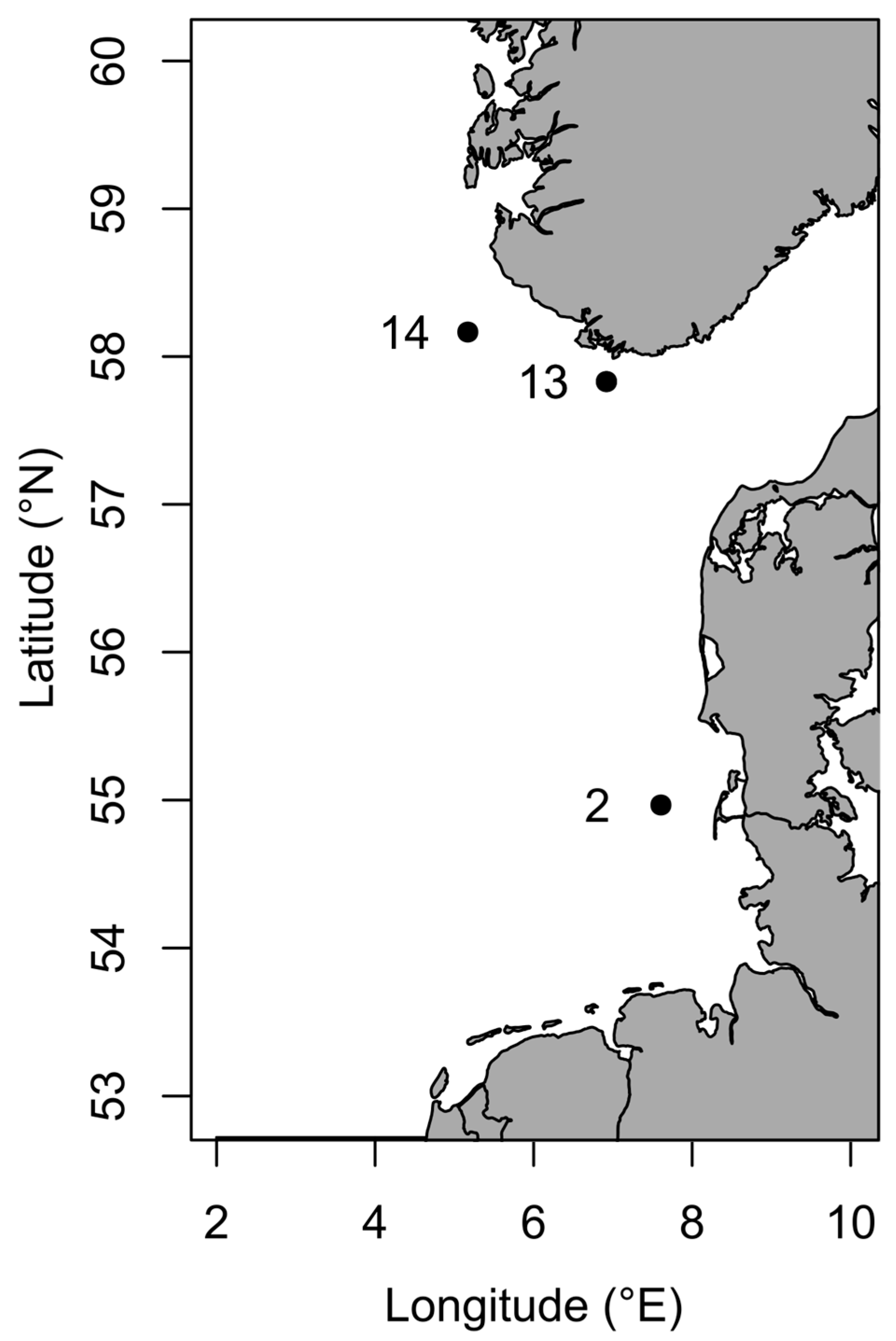
3.1. Sampling and Sample Preparation
3.2. DNA Extraction and Sequencing
3.3. Assembly and Genome Reconstruction
Supplementary Materials
Funding
Acknowledgments
Conflicts of Interest
References
- Wiltshire, K.H.; Kraberg, A.; Bartsch, I.; Boersma, M.; Franke, H.-D.; Freund, J.; Gebühr, C.; Gerdts, G.; Stockmann, K.; Wichels, A. Helgoland Roads, North Sea: 45 Years of Change. Estuaries Coasts 2010, 33, 295–310. [Google Scholar] [CrossRef] [Green Version]
- McQuatters-Gollop, A.; Raitsos, D.E.; Edwards, M.; Pradhan, Y.; Mee, L.D.; Lavender, S.J.; Attrill, M.J. A long-term chlorophyll data set reveals regime shift in North Sea phytoplankton biomass unconnected to nutrient trends. Limnol. Oceanogr. 2007, 52, 635. [Google Scholar] [CrossRef]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Brownrigg, R. Maps, 3.3.0. 2018. Available online: https://cran.r-project.org/web/packages/maps/ (accessed on 1 January 2020).
- Brownrigg, R. MapData, 2.3.0. CRAN, 2018. Available online: https://cran.r-project.org/web/packages/mapdata/index.html (accessed on 1 January 2020).
- Chaumeil, P.-A.; Mussig, A.J.; Hugenholtz, P.; Parks, D.H. GTDB-Tk: A toolkit to classify genomes with the Genome Taxonomy Database. Bioinformatics 2019. [Google Scholar] [CrossRef] [PubMed]
- Pernthaler, A.; Preston, C.M.; Pernthaler, J.; DeLong, E.F.; Amann, R. Comparison of Fluorescently Labeled Oligonucleotide and Polynucleotide Probes for the Detection of Pelagic Marine Bacteria and Archaea. Appl. Environ. Microbiol. 2002, 68, 661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wemheuer, B.; Wemheuer, F.; Daniel, R. RNA-Based Assessment of Diversity and Composition of Active Archaeal Communities in the German Bight. Archaea 2012, 2012, 695826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.L.; Xie, W.; Martin-Cuadrado, A.-B.; Rodriguez-Valera, F. Marine Group II Archaea, potentially important players in the global ocean carbon cycle. Front. Microbiol. 2015, 6, 1108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giebel, H.A.; Brinkhoff, T.; Zwisler, W.; Selje, N.; Simon, M. Distribution of Roseobacter RCA and SAR11 lineages and distinct bacterial communities from the subtropics to the Southern Ocean. Environ. Microbiol. 2009, 11, 2164–2178. [Google Scholar] [CrossRef]
- Wemheuer, B.; Güllert, S.; Billerbeck, S.; Giebel, H.-A.; Voget, S.; Simon, M.; Daniel, R. Impact of a phytoplankton bloom on the diversity of the active bacterial community in the southern North Sea as revealed by metatranscriptomic approaches. Fems Microbiol. Ecol. 2014, 87, 378–389. [Google Scholar] [CrossRef] [Green Version]
- Giebel, H.A.; Kalhoefer, D.; Lemke, A.; Thole, S.; Gahl-Janssen, R.; Simon, M.; Brinkhoff, T. Distribution of Roseobacter RCA and SAR11 lineages in the North Sea and characteristics of an abundant RCA isolate. ISME J. 2011, 5, 8–19. [Google Scholar] [CrossRef]
- Wemheuer, B.; Wemheuer, F.; Hollensteiner, J.; Meyer, F.-D.; Voget, S.; Daniel, R. The green impact: Bacterioplankton response towards a phytoplankton spring bloom in the southern North Sea assessed by comparative metagenomic and metatranscriptomic approaches. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Voget, S.; Wemheuer, B.; Brinkhoff, T.; Vollmers, J.; Dietrich, S.; Giebel, H.-A.; Beardsley, C.; Sardemann, C.; Bakenhus, I.; Billerbeck, S.; et al. Adaptation of an abundant Roseobacter RCA organism to pelagic systems revealed by genomic and transcriptomic analyses. ISME J. 2015, 9, 371–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucas, J.; Wichels, A.; Teeling, H.; Chafee, M.; Scharfe, M.; Gerdts, G. Annual dynamics of North Sea bacterioplankton: Seasonal variability superimposes short-term variation. Fems Microbiol. Ecol. 2015, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wemheuer, B.; Wemheuer, F.; Meier, D.; Billerbeck, S.; Giebel, H.-A.; Simon, M.; Scherber, C.; Daniel, R. Linking Compositional and Functional Predictions to Decipher the Biogeochemical Significance in DFAA Turnover of Abundant Bacterioplankton Lineages in the North Sea. Microorganisms 2017, 5, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, S.; Fuchs, B.M.; Lenk, S.; Harder, J.; Wulf, J.; Jiao, N.Z.; Amann, R. Biogeography and phylogeny of the NOR5/OM60 clade of Gammaproteobacteria. Syst. Appl. Microbiol. 2009, 32, 124–139. [Google Scholar] [CrossRef] [PubMed]
- Stingl, U.; Desiderio, R.A.; Cho, J.C.; Vergin, K.L.; Giovannoni, S.J. The SAR92 clade: An abundant coastal clade of culturable marine bacteria possessing proteorhodopsin. Appl. Environ. Microbiol. 2007, 73, 2290–2296. [Google Scholar] [CrossRef] [Green Version]
- Billerbeck, S.; Wemheuer, B.; Voget, S.; Poehlein, A.; Giebel, H.-A.; Brinkhoff, T.; Gram, L.; Jeffrey, W.H.; Daniel, R.; Meinhard, S. Biogeography and environmental genomics of the Roseobacter group affiliated pelagic CHAB-I-5 lineage. Nat. Microbiol. 2016, 1, 16063. [Google Scholar] [CrossRef]
- Tsementzi, D.; Wu, J.; Deutsch, S.; Nath, S.; Rodriguez-R, L.M.; Burns, A.S.; Ranjan, P.; Sarode, N.; Malmstrom, R.R.; Padilla, C.C.; et al. SAR11 bacteria linked to ocean anoxia and nitrogen loss. Nature 2016, 536, 179–183. [Google Scholar] [CrossRef] [Green Version]
- Osterholz, H.; Singer, G.; Wemheuer, B.; Daniel, R.; Simon, M.; Niggemann, J.; Dittmar, T. Deciphering associations between dissolved organic molecules and bacterial communities in a pelagic marine system. ISME J. 2016. [Google Scholar] [CrossRef]
- Weinbauer, M.G.; Fritz, I.; Wenderoth, D.F.; Höfle, M.G. Simultaneous extraction from bacterioplankton of total RNA and DNA suitable for quantitative structure and function analyses. Appl. Environ. Microbiol. 2002, 68, 1082–1087. [Google Scholar] [CrossRef] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Meth. 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, D.D.; Froula, J.; Egan, R.; Wang, Z. MetaBAT, an efficient tool for accurately reconstructing single genomes from complex microbial communities. Peer J. 2015, 3, e1165. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.-H.; Liao, Y.-C. Accurate binning of metagenomic contigs via automated clustering sequences using information of genomic signatures and marker genes. Sci. Rep. 2016, 6, 24175. [Google Scholar] [CrossRef]
- Song, W.-Z.; Thomas, T. Binning–refiner: Improving genome bins through the combination of different binning programs. Bioinformatics 2017, 33, 1873–1875. [Google Scholar] [CrossRef]
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef] [Green Version]
- Parks, D.H.; Chuvochina, M.; Chaumeil, P.-A.; Rinke, C.; Mussig, A.J.; Hugenholtz, P. Selection of representative genomes for 24,706 bacterial and archaeal species clusters provide a complete genome-based taxonomy. BioRxiv 2019, 771964. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wemheuer, B. A Collection of 13 Archaeal and 46 Bacterial Genomes Reconstructed from Marine Metagenomes Derived from the North Sea. Data 2020, 5, 15. https://doi.org/10.3390/data5010015
Wemheuer B. A Collection of 13 Archaeal and 46 Bacterial Genomes Reconstructed from Marine Metagenomes Derived from the North Sea. Data. 2020; 5(1):15. https://doi.org/10.3390/data5010015
Chicago/Turabian StyleWemheuer, Bernd. 2020. "A Collection of 13 Archaeal and 46 Bacterial Genomes Reconstructed from Marine Metagenomes Derived from the North Sea" Data 5, no. 1: 15. https://doi.org/10.3390/data5010015
APA StyleWemheuer, B. (2020). A Collection of 13 Archaeal and 46 Bacterial Genomes Reconstructed from Marine Metagenomes Derived from the North Sea. Data, 5(1), 15. https://doi.org/10.3390/data5010015