
The Effects of Respiratory Vaccine Type and Timing on Antibody Titers, Immunoglobulins, and Growth Performance in Pre- and Post-Weaned Beef Calves

 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Treatment and Vaccination Procedures
2.2. Animal Management
2.3. Blood Collection and Serology
2.4. Serum Immunoglobulins
2.5. Statistical Analysis
3. Results
3.1. Body Weight and Average Daily Gain
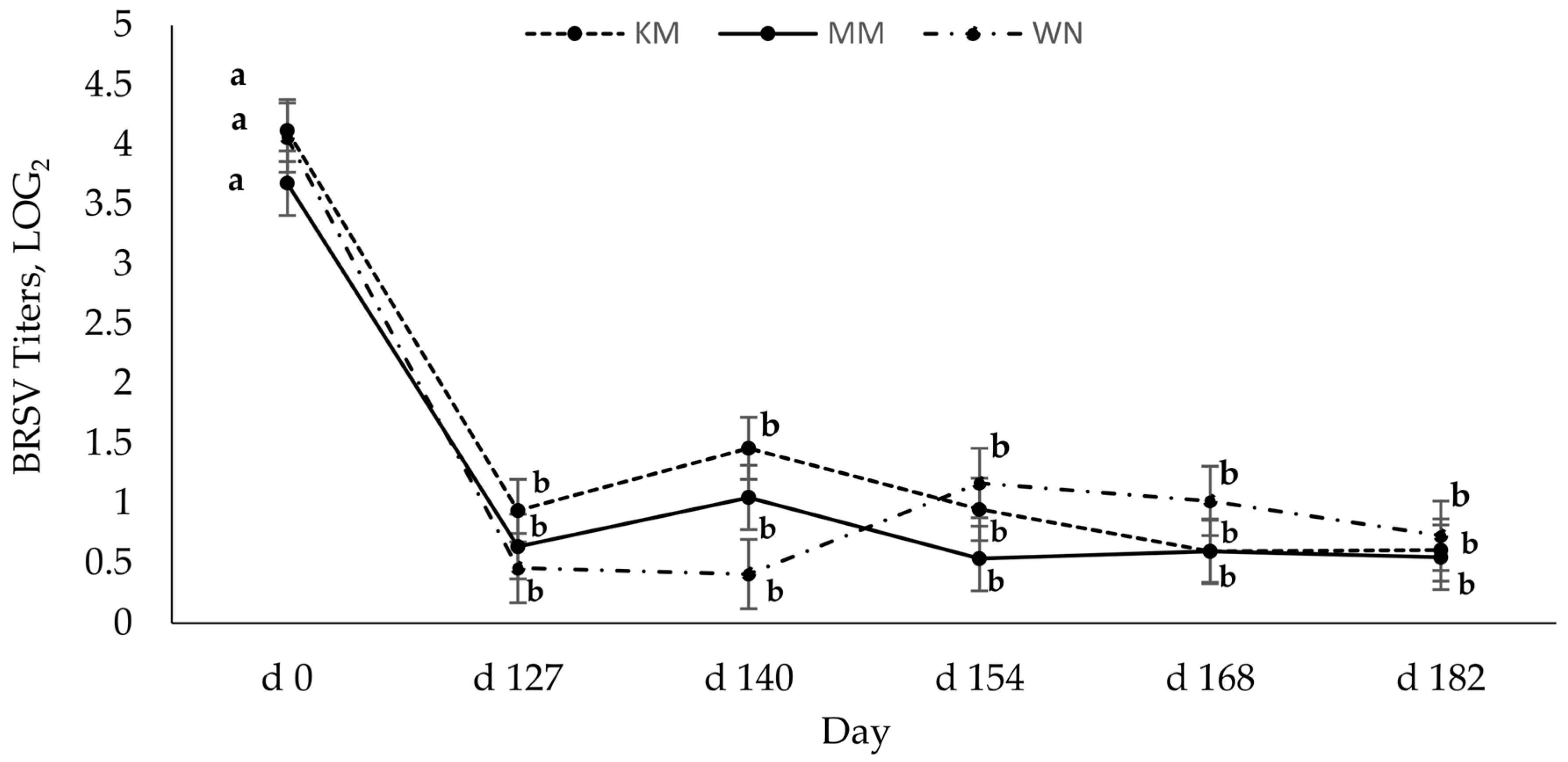
3.2. Bovine Respiratory Syncytial Virus SN Antibody Titers
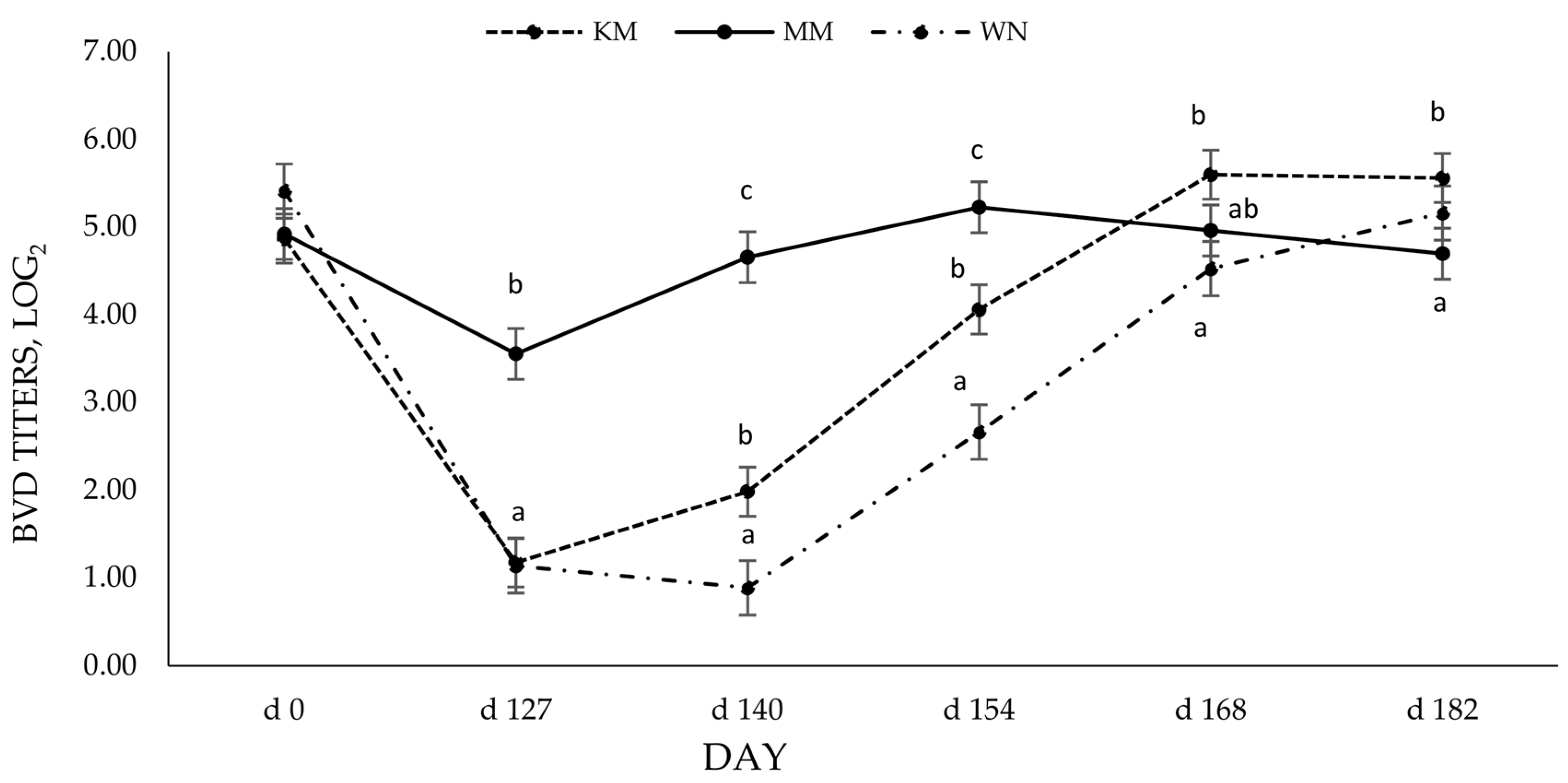
3.3. Bovine Viral Diarrhea Virus SN Antibody Titers
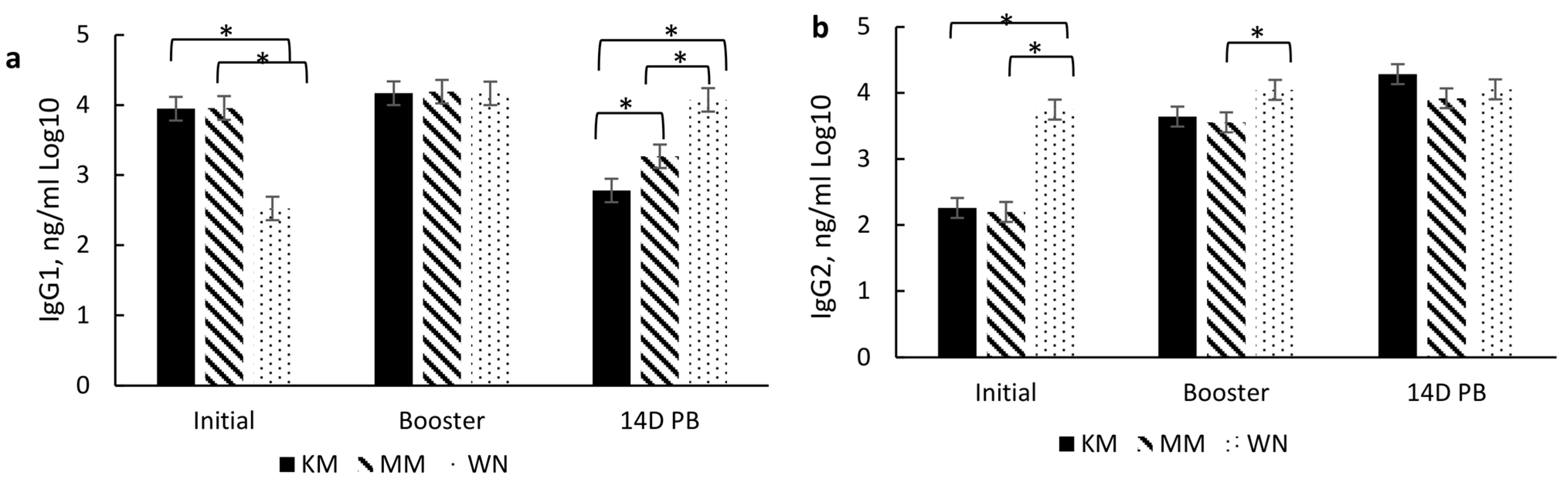
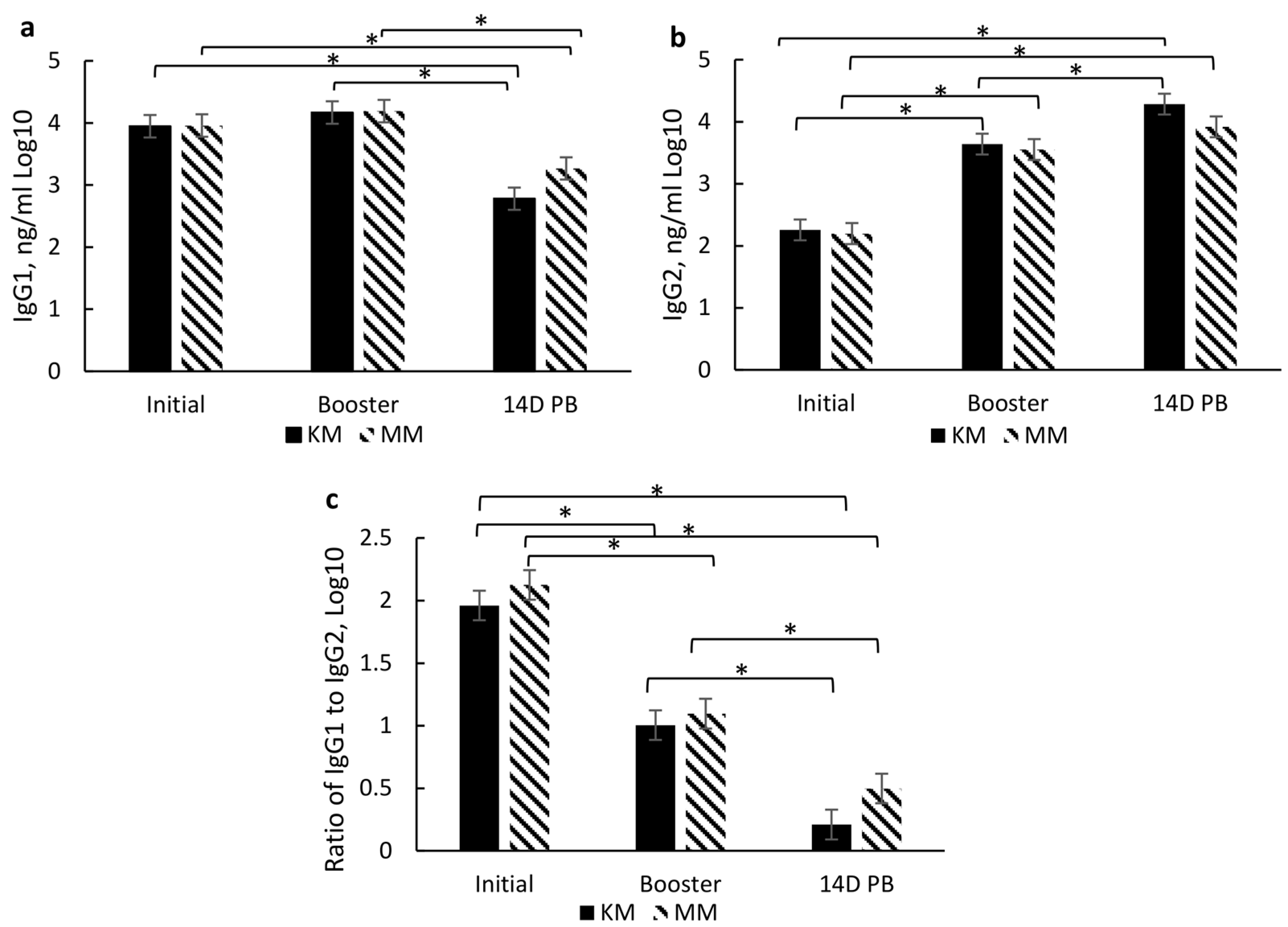
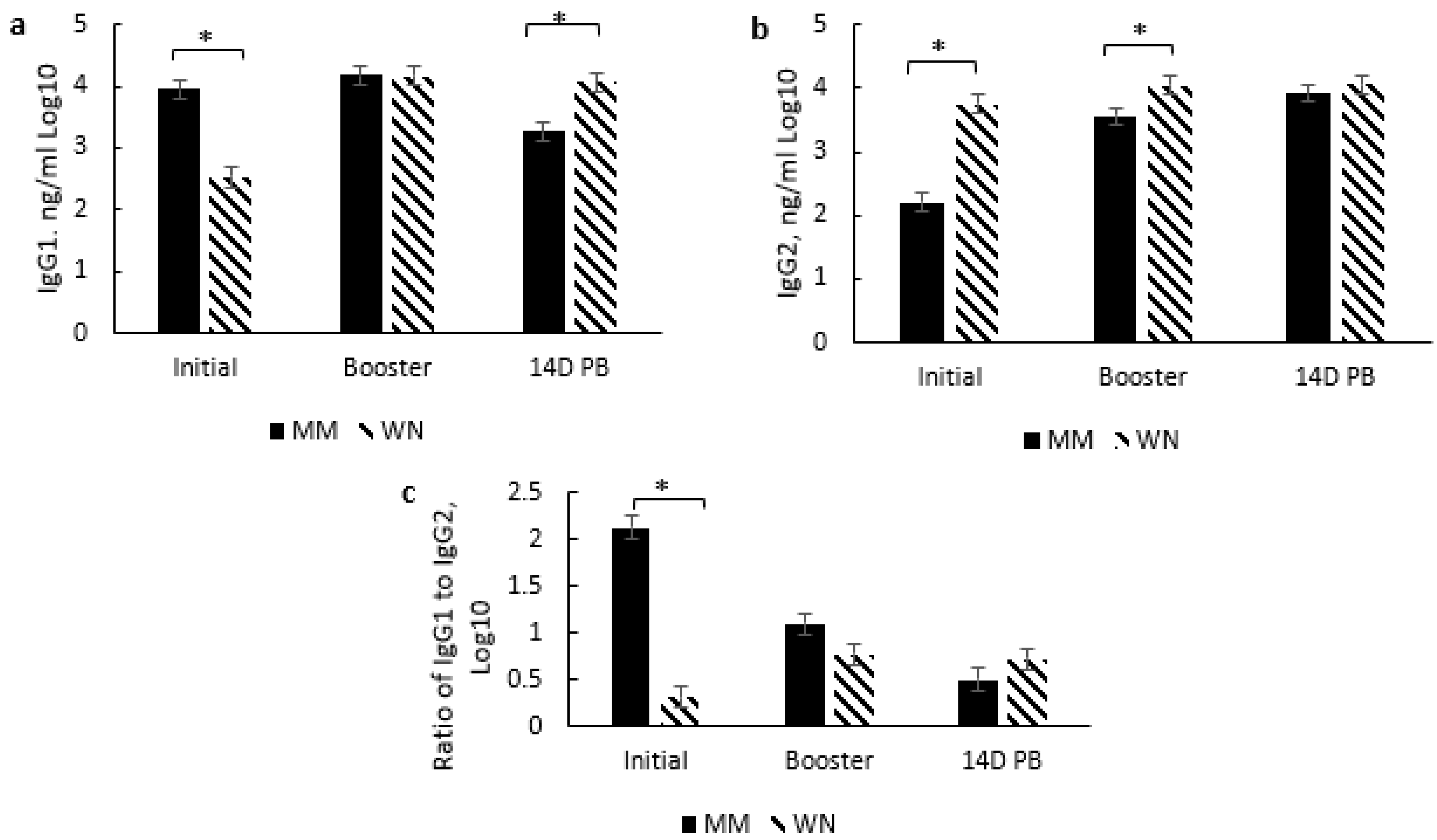
3.4. ImmunoglobulinG and Subsets Response to Type and Timing of Vaccines
3.4.1. Overall Effects of Treatment
3.4.2. Type of Vaccination and Timing of Vaccination
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hilton, W.M. Management of Preconditioned Calves and Impacts of Preconditioning. Vet. Clin. N. Am. Food Anim. Pract. 2015, 31, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Robe, J.; Biggs, R.; Beck, P.A. Oklahoma Quality Beef Network VAC-45 Program Requirements; Oklahoma State University Cooperative Extension (AFS-3305) (Fact Sheet); Oklahoma State University: Stillwater, OK, USA, 2022. [Google Scholar]
- Anglen, C.S.; Truckenmiller, M.E.; Schell, T.D.; Bonneau, R.H. The dual role of CD8+ T lymphocytes in the development of stress-induced herpes simplex encephalitis. J. Neuroimmunol. 2003, 140, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Kirkpatrick, J.G.; Step, D.L.; Payton, M.E.; Richards, J.B.; McTague, L.F.; Saliki, J.T.; Confer, A.W.; Cook, B.J.; Ingram, S.H.; Wright, J.C. Effect of age at the time of vaccination on antibody titers and feedlot performance in beef calves. J. Am. Vet. Med. Assoc. 2008, 233, 136–142. [Google Scholar] [CrossRef] [PubMed]
- West, K.; Petrie, L.; Konoby, C.; Haines, D.M.; Cortese, V.; Ellis, J.A. The efficacy of modified-live bovine respiratory syncytial virus vaccines in experimentally infected calves. Vaccine 1999, 18, 907–919. [Google Scholar] [CrossRef] [PubMed]
- Edwards, T.A. Control methods for bovine respiratory disease for feedlot cattle. Vet. Clin. N. Am. Food Anim. Pract. 2010, 26, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Endsley, J.J.; Roth, J.A.; Ridpath, J.; Neill, J. Maternal antibody blocks humoral but not T cell responses to BVDV. Biologicals 2003, 31, 123–125. [Google Scholar] [CrossRef] [Green Version]
- Chase, C.C.; Hurley, D.J.; Reber, A.J. Neonatal immune development in the calf and its impact on vaccine response. Vet. Clin. N. Am. Food Anim. Pract. 2008, 24, 87–104. [Google Scholar] [CrossRef]
- Richeson, J.; Kegley, E.; Gadberry, M.; Beck, P.; Powell, J.; Jones, C. Effects of on-arrival versus delayed clostridial or modified live respiratory vaccinations on health, performance, bovine viral diarrhea virus type I titers, and stress and immune measures of newly received beef calves. J. Anim. Sci. 2009, 87, 2409–2418. [Google Scholar] [CrossRef] [Green Version]
- Richeson, J.T.; Beck, P.A.; Gadberry, M.S.; Gunter, S.A.; Hess, T.W.; Hubbell, D.S., III; Jones, C. Effects of on-arrival versus delayed modified live virus vaccination on health, performance, and serum infectious bovine rhinotracheitis titers of newly received beef calves. J. Anim. Sci. 2008, 86, 999–1005. [Google Scholar] [CrossRef]
- Chamorro, M.F.; Walz, P.H.; Passler, T.; van Santen, E.; Gard, J.; Rodning, S.P.; Riddell, K.P.; Galik, P.K.; Zhang, Y. Efficacy of multivalent, modified- live virus (MLV) vaccines administered to early weaned beef calves subsequently challenged with virulent Bovine viral diarrhea virus type 2. BMC Vet. Res. 2015, 11, 29. [Google Scholar] [CrossRef]
- Grooms, D.L.; Coe, P. Neutralizing antibody responses in preconditioned calves following vaccination for respiratory viruses. Vet. Ther. Res. Appl. Vet. Med. 2002, 3, 119–127. [Google Scholar]
- BQA. Beef Quality Assurance National Manual. National Cattlemen’s Beef Association. Centennial, CO. 2019. Available online: https://www.bqa.org/Media/BQA/Docs/bqa_manual_final.pdf (accessed on 16 November 2022).
- Price, E.; Harris, J.; Borgwardt, R.; Sween, M.; Connor, J. Fenceline contact of beef calves with their dams at weaning reduces the negative effects of separation on behavior and growth rate. J. Anim. Sci. 2003, 81, 116–121. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.; Holland, B.; Step, D.; Payton, M.; Confer, A. Nasal isolation of Mannheimia haemolytica and Pasteurella multocida as predictors of respiratory disease in shipped calves. Res. Vet. Sci. 2015, 99, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Ridpath, J.E.; Neill, J.D.; Endsley, J.; Roth, J.A. Effect of passive immunity on the development of a protective immune response against bovine viral diarrhea virus in calves. Am. J. Vet. Res. 2003, 64, 65–69. [Google Scholar] [CrossRef]
- Downey-Slinker, E.D.; Ridpath, J.F.; Sawyer, J.E.; Skow, L.C.; Herring, A.D. Antibody titers to vaccination are not predictive of level of protection against a BVDV type 1b challenge in Bos indicus-Bos taurus steers. Vaccine 2016, 34, 5053–5059. [Google Scholar] [CrossRef] [Green Version]
- Fulton, R.W.; Confer, A.W.; Burge, L.J.; Perino, L.J.; d’Offay, J.M.; Payton, M.E.; Mock, R.E. Antibody responses by cattle after vaccination with commercial viral vaccines containing bovine herpesvirus-1, bovine viral diarrhea virus, parainfluenza-3 virus, and bovine respiratory syncytial virus immunogens and subsequent revaccination at day 140. Vaccine 1995, 13, 725–733. [Google Scholar] [CrossRef]
- Bolin, S.R.; Ridpath, J.F. Assessment of protection from systemic infection or disease afforded by low to intermediate titers of passively acquired neutralizing antibody against bovine viral diarrhea virus in calves. Am. J. Vet. Res. 1995, 56, 755–759. [Google Scholar]
- Peters, A.; Thevasagayam, S.; Wiseman, A.; Salt, J. Duration of immunity of a quadrivalent vaccine against respiratory diseases caused by BHV-1, PI3V, BVDV, and BRSV in experimentally infected calves. Prevent Vet. Med. 2004, 66, 63–77. [Google Scholar] [CrossRef]
- West, K.; Ellis, J. Functional analysis of antibody responses of feedlot cattle to bovine respiratory syncytial virus following vaccination with mixed vaccines. Can. J. Vet. Res. 1997, 61, 28–33. [Google Scholar]
- Van Donkersgoed, J.; Van den Hurk, J.V.; McCartney, D.; Harland, R.J. Comparative serological responses in calves to eight commercial vaccines against infectious bovine rhinotracheitis, parainfluenza-3, bovine respiratory syncytial, and bovine viral diarrhea viruses. Can. Vet. J. 1991, 32, 727–733. [Google Scholar]
- Kandel, A.; Masello, M.; Xiao, Z. CD4+ T Cell Responses to Pathogens in Cattle. In Bovine Science-Challenges and Advances; IntechOpen: London, UK, 2021. [Google Scholar] [CrossRef]
- Yang, K.; Whalen, B.J.; Tirabassi, R.S.; Selin, L.K.; Levchenko, T.S.; Torchilin, V.P.; Kislauskis, E.H.; Guberski, D.L. A DNA vaccine prime followed by a liposome-encapsulated protein boost confers enhanced mucosal immune responses and protection. J. Immunol. 2008, 180, 6159–6167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letellier, C.; Boxus, M.; Rosar, L.; Toussaint, J.F.; Walravens, K.; Roels, S.; Meyer, G.; Letesson, J.J.; Kerkhofs, P. Vaccination of calves using the BRSV nucleocapsid protein in a DNA prime-protein boost strategy stimulates cell-mediated immunity and protects the lungs against BRSV replication and pathology. Vaccine 2008, 26, 4840–4848. [Google Scholar] [CrossRef] [PubMed]
- Kalina, W.V.; Woolums, A.R.; Gershwin, L.J. Formalin-inactivated bovine RSV vaccine influences antibody levels in bronchoalveolar lavage fluid and disease outcome in experimentally infected calves. Vaccine 2005, 23, 4625–4630. [Google Scholar] [CrossRef] [PubMed]
- Kirkpatrick, J.; Fulton, R.W.; Burge, L.J.; DuBois, W.R.; Payton, M. Passively Transferred Immunity in Newborn Calves, Rate of Antibody Decay, and Effect on Subsequent Vaccination with Modified Live Virus Vaccine. Bov. Pract. 2019, 35, 47–55. [Google Scholar] [CrossRef]
- Duff, G.C.; Malcolm-Callis, K.J.; Walker, D.A.; Wiseman, M.W.; Galyean, M.L.; Perino, L.J. Effects of Intranasal Versus Intramuscular Modified Live Vaccines and Vaccine Timing on Health and Performance by Newly Received Beef Cattle. Bov. Pract. 2019, 34, 66–71. [Google Scholar] [CrossRef]
- Powell, J.G.; Richeson, J.T.; Kegley, E.B.; Coffey, K.P.; Erf, G.F.; Brown, A.H.; Downum, W.; Ensley, D.T. Immunologic, health, and growth responses of beef calves administered pentavalent modified-live virus respiratory vaccine in the presence of maternal antibody versus a traditional vaccination regimen. Bov. Pract. 2012, 46, 122–130. [Google Scholar] [CrossRef]
- Step, D.L.; Krehbiel, C.R.; Burciaga-Robles, L.O.; Holland, B.P.; Fulton, R.W.; Confer, A.W.; Bechtol, D.T.; Brister, D.L.; Hutcheson, J.P.; Newcomb, H.L. Comparison of single vaccination versus revaccination with a modified-live virus vaccine containing bovine herpesvirus-1, bovine viral diarrhea virus (types 1a and 2a), parainfluenza type 3 virus, and bovine respiratory syncytial virus in the prevention of bovine respiratory disease in cattle. J. Am. Vet. Med. Assoc. 2009, 235, 580–587. [Google Scholar] [CrossRef]
- O’Connor, A.; Hu, D.; Totton, S.; Scott, N.; Winder, C.; Wang, B.; Wang, C.; Glanville, J.; Wood, H.; White, B. A systematic review and network meta-analysis of bacterial and viral vaccines, administered at or near arrival at the feedlot, for control of bovine respiratory disease in beef cattle. Anim. Health Res. Rev. 2019, 20, 143–162. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | p Value | ||||||
|---|---|---|---|---|---|---|---|
| Item | KM 1 | MM 2 | WN 3 | SE | Sex | Trt | Sex × Trt |
| Body weight, kg | |||||||
| Day 0 | 103 | 103 | 103 | 2.8 | 0.03 | 0.99 | 0.82 |
| Day 127 | 244 | 244 | 244 | 8.4 | <0.01 | 0.99 | 0.39 |
| Day 140 | 250 | 251 | 252 | 9.2 | <0.01 | 0.94 | 0.68 |
| Day 154 | 259 | 263 | 257 | 10.3 | <0.01 | 0.70 | 0.61 |
| Day 168 | 262 | 267 | 264 | 10.0 | <0.01 | 0.74 | 0.74 |
| Day 182 | 276 | 276 | 275 | 7.4 | <0.01 | 0.99 | 0.56 |
| Average Daily Gain, kg | |||||||
| Day 0 to 127 | 1.11 | 1.11 | 1.11 | 0.036 | <0.01 | 0.97 | 0.20 |
| Day 127 to 140 | 0.48 | 0.58 | 0.62 | 0.356 | 0.94 | 0.63 | 0.16 |
| Day 140 to 154 | 0.73 | 0.81 | 0.43 | 0.144 | 0.52 | 0.08 | 0.19 |
| Day 154 to 168 | 0.19 | 0.33 | 0.48 | 0.133 | 0.50 | 0.19 | 0.13 |
| Day 168 to 182 | 0.96 | 0.60 | 0.78 | 0.233 | 0.06 | 0.09 | 0.39 |
| Day 127 to 168 | 0.59 | 0.58 | 0.58 | 0.057 | 0.81 | 0.97 | 0.88 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matty, J.M.; Reddout, C.; Adams, J.; Major, M.; Lalman, D.; Biggs, R.; Salak-Johnson, J.L.; Beck, P.A. The Effects of Respiratory Vaccine Type and Timing on Antibody Titers, Immunoglobulins, and Growth Performance in Pre- and Post-Weaned Beef Calves. Vet. Sci. 2023, 10, 37. https://doi.org/10.3390/vetsci10010037
Matty JM, Reddout C, Adams J, Major M, Lalman D, Biggs R, Salak-Johnson JL, Beck PA. The Effects of Respiratory Vaccine Type and Timing on Antibody Titers, Immunoglobulins, and Growth Performance in Pre- and Post-Weaned Beef Calves. Veterinary Sciences. 2023; 10(1):37. https://doi.org/10.3390/vetsci10010037
Chicago/Turabian StyleMatty, Jeff M., Cassidy Reddout, Jordan Adams, Mike Major, David Lalman, Rosslyn Biggs, Janeen L. Salak-Johnson, and Paul A. Beck. 2023. "The Effects of Respiratory Vaccine Type and Timing on Antibody Titers, Immunoglobulins, and Growth Performance in Pre- and Post-Weaned Beef Calves" Veterinary Sciences 10, no. 1: 37. https://doi.org/10.3390/vetsci10010037
APA StyleMatty, J. M., Reddout, C., Adams, J., Major, M., Lalman, D., Biggs, R., Salak-Johnson, J. L., & Beck, P. A. (2023). The Effects of Respiratory Vaccine Type and Timing on Antibody Titers, Immunoglobulins, and Growth Performance in Pre- and Post-Weaned Beef Calves. Veterinary Sciences, 10(1), 37. https://doi.org/10.3390/vetsci10010037