1. Introduction
Wild populations of rhesus monkeys (
Macaca mulatta) live in the southeastern parts of Asia, but this species can also be found in research facilities worldwide, as it is one of the most studied nonhuman primates [
1,
2]. They serve as animal models in toxicity studies, aid in unraveling the pathogenesis of various diseases and are key in the development of many vaccines against, e.g., Malaria, West Nile virus and H5N1 Influenza [
3,
4]. The genome of the rhesus monkey is 93.5% similar to that of humans [
5]. Consequently, the two species have many phenotypical similarities [
6].
Rhesus monkeys that are housed in zoos and primate centers often receive drugs, including anesthetics, by means of remote methodologies, such as darting. However, this intervention can cause trauma, ranging from minor lacerations to fractured bones. Another restraint technique relies on the use of a “restraint” or “squeeze” cage, in which the back is gently pulled forward to allow injections. This not only induces high stress levels but also can provoke mechanical muscle damage when the animal is not cooperative.
To ensure the safety of intramuscular (IM), subcutaneous (SC) and intravenous (IV) injections and related practices, anatomical knowledge is mandatory. In addition, it is essential to have a good knowledge of the anatomy of the rhesus monkey to diagnose and treat observed locomotor problems related to the lower limbs (membrum pelvinum, pelvic limb, hindlimb or hindleg). Unfortunately, the literature on the anatomy of the rhesus monkey is scarce. As the book by Hartman and Straus [
7] contains only line drawings, it fails to represent the authentic condition. The atlas by Berringer et al. [
8] contains only black-and-white pictures that lack clarity. In the recent book chapter by Casteleyn and Bakker [
9], the anatomy of the rhesus monkey is illustrated with many color images. Nevertheless, it may be more valuable to anatomists than to veterinary practitioners since the anatomy is described per system and not topographically. This could limit the use of the book chapter during surgical interventions, e.g., when wounds have to be tended to [
10].
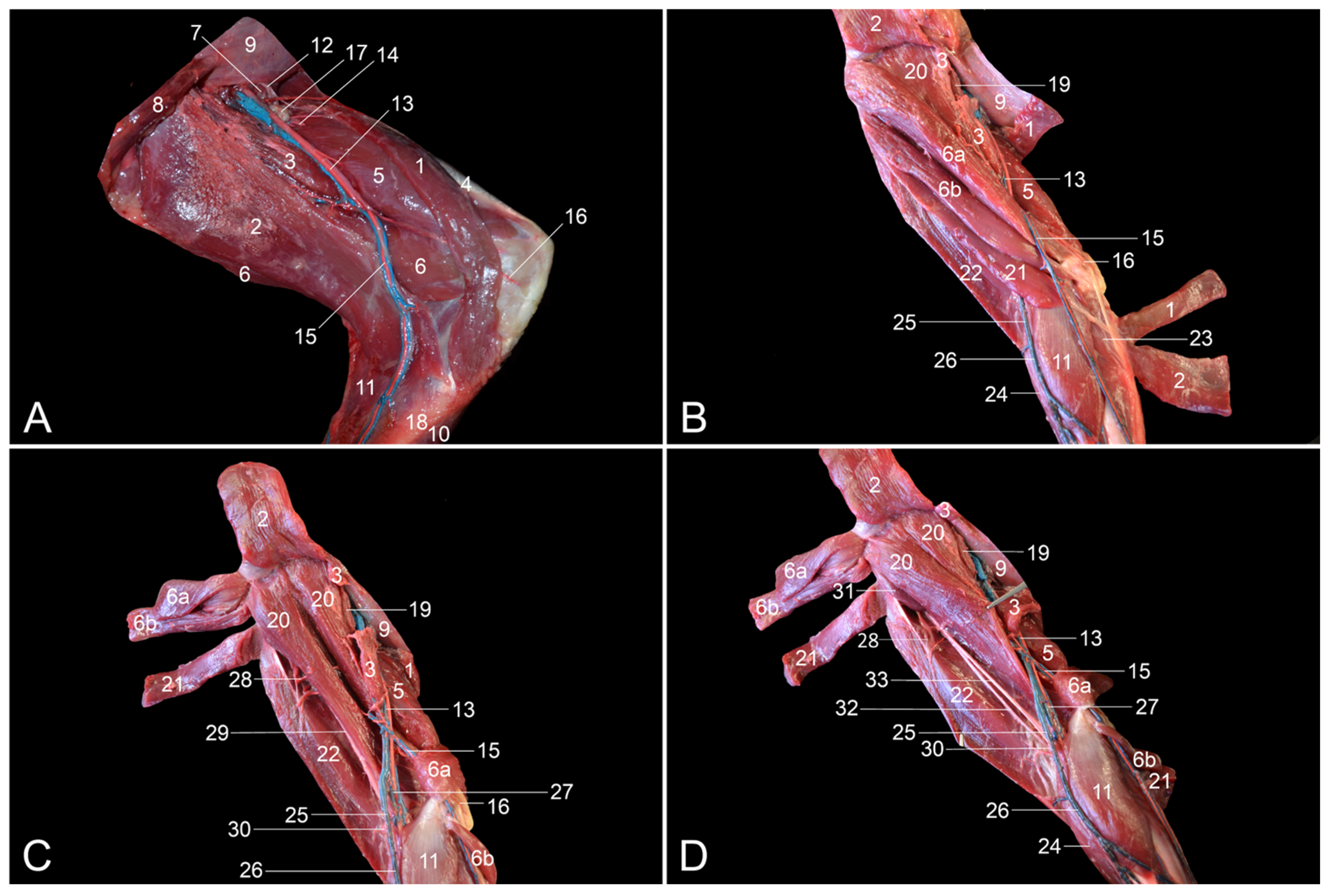
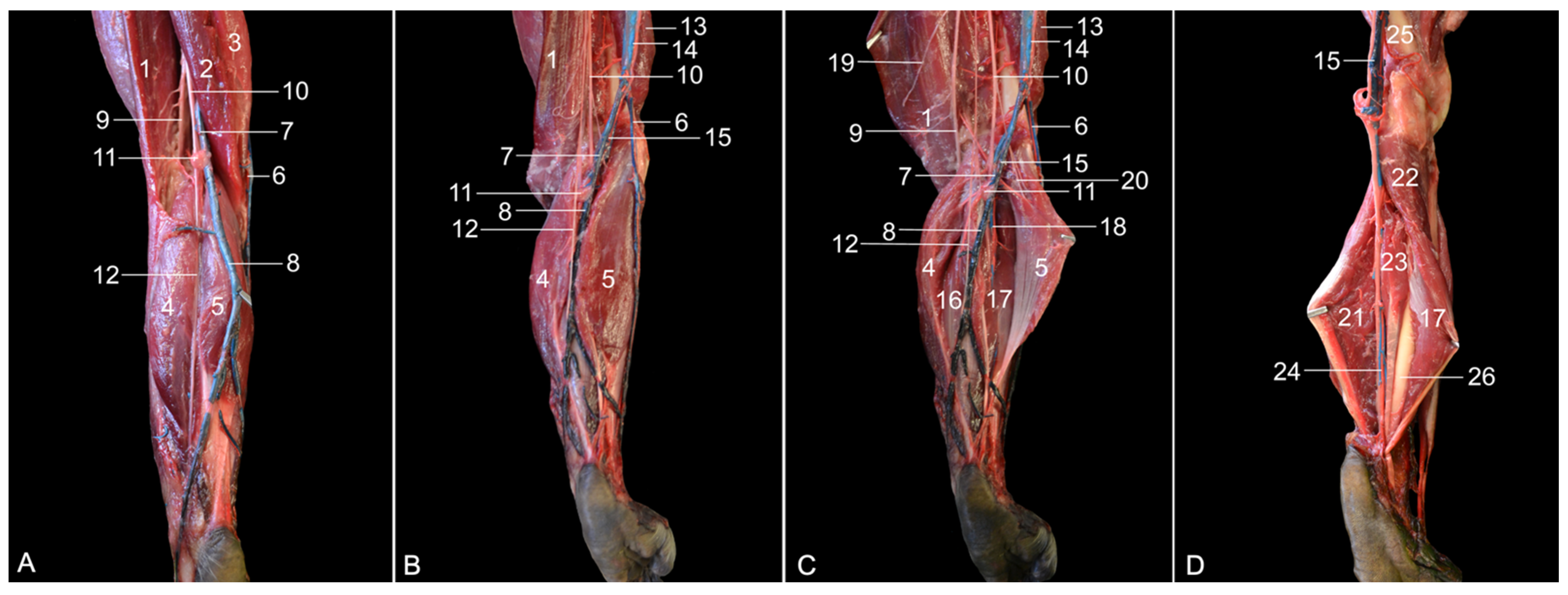
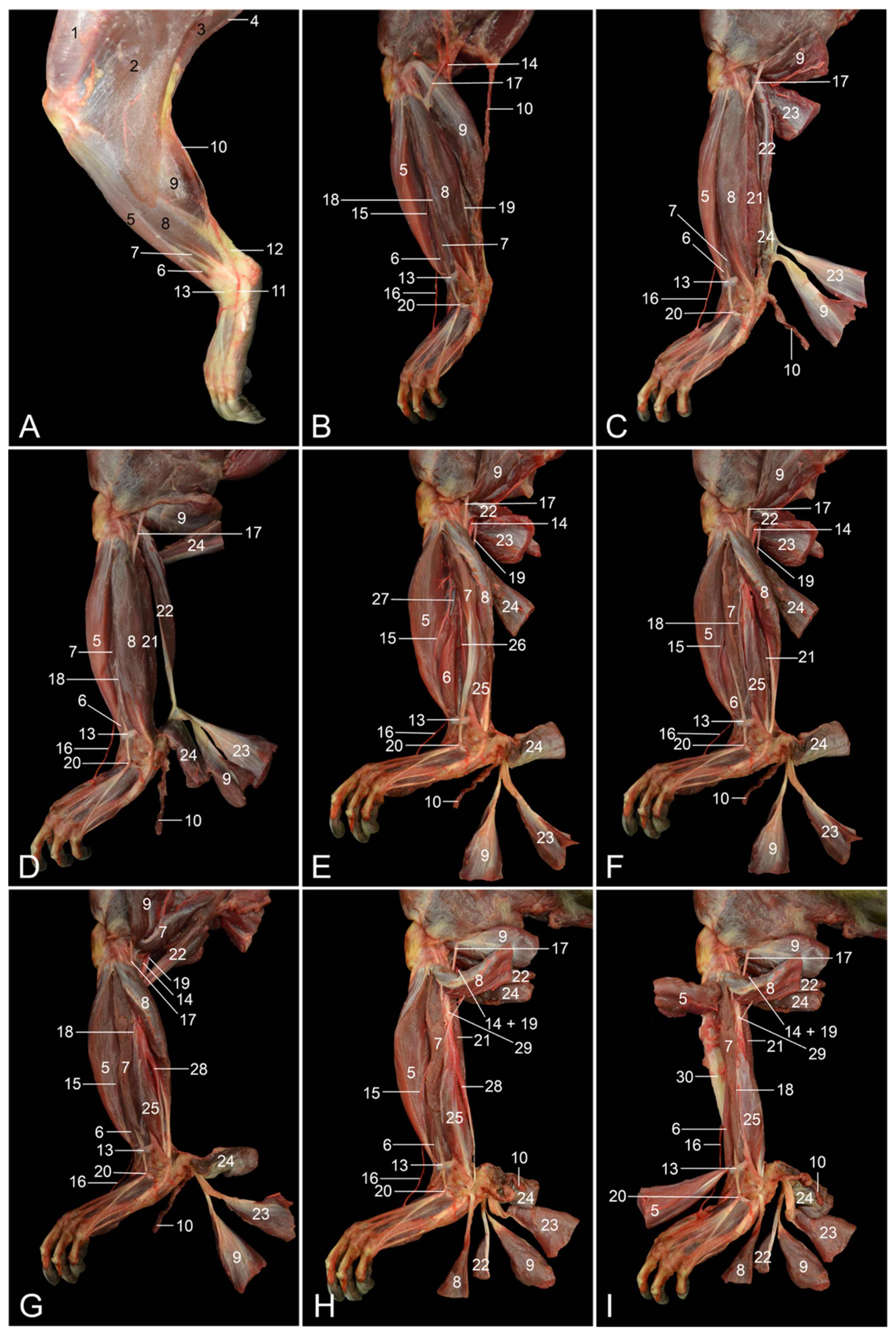
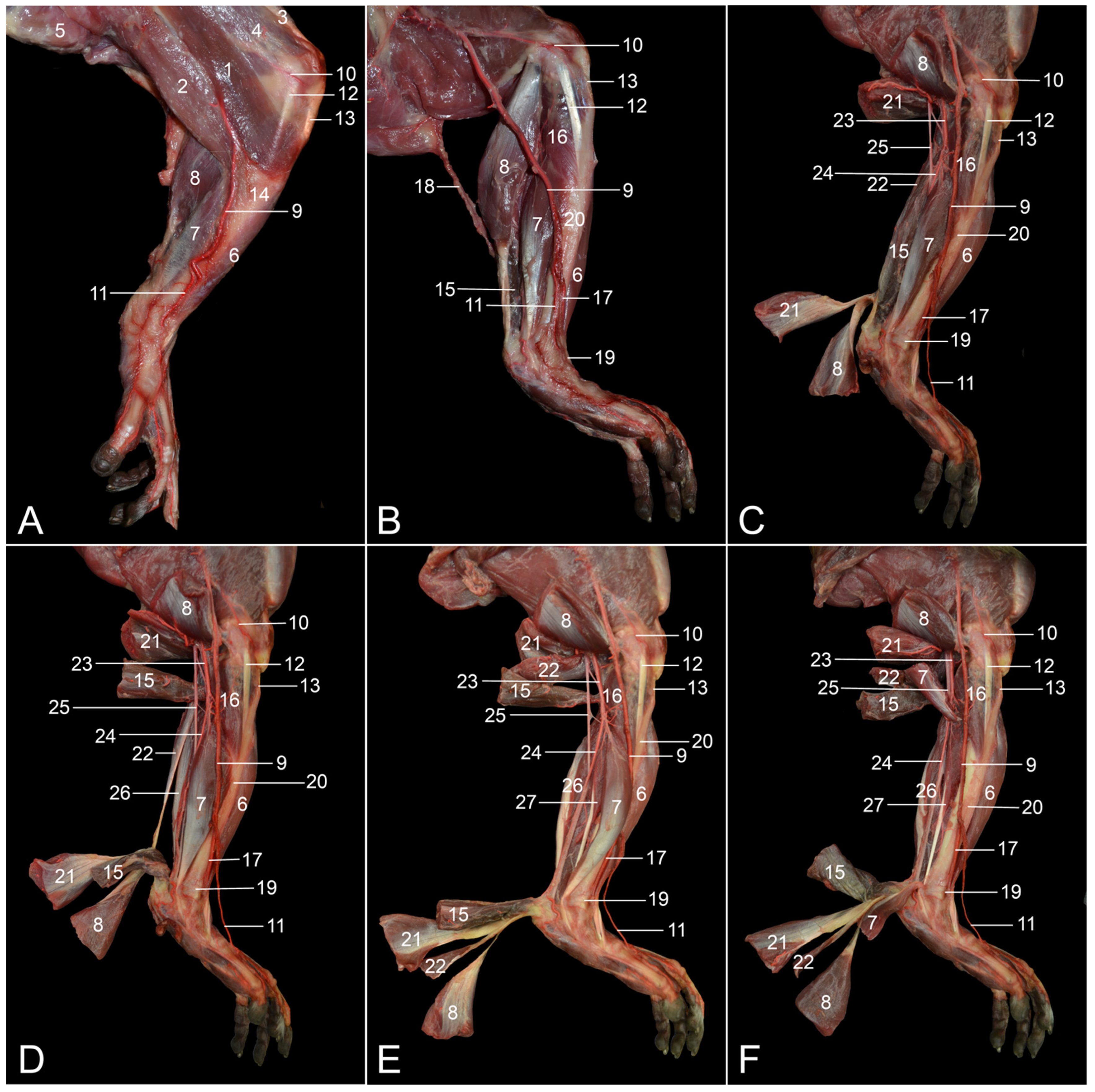
The objectives of this study were to describe the anatomy of the hindlimb of the rhesus monkey in a topographical manner and to identify the similarities and differences between the anatomy of the human leg and that of the rhesus monkey hindlimb. Therefore, the various anatomical systems are described, per region, in relation to each other. The textual descriptions are illustrated with numerous full-color images, thus making this article an extremely useful reference work for researchers and veterinarians.
4. Discussion
During this anatomical study, remarkable similarities between the rhesus monkey and a human were observed. Since these similarities stretch beyond anatomy and also apply to physiology, the rhesus monkey plays an important role as a model for humans [
2,
5]. However, the anatomy of the rhesus monkey does not appear to be identical to human anatomy. Several significant discrepancies were observed when comparing the dissected specimens with images in a human anatomy atlas [
16]. Access to a state-of-the-art anatomy atlas on the rhesus monkey could therefore be valuable to researchers who seek to extrapolate the results obtained in this nonhuman primate model to humans. Such a publication could also be consulted by veterinarians responsible for the medical care and welfare of rhesus monkeys kept in captivity. These professionals primarily rely on their knowledge of domestic mammal anatomy when treating, for example, wounds or performing surgery on a rhesus monkey. Although the anatomy of this species is remarkably similar to that of domestic mammals, the subtle dissimilarities could result in uncertainty about the optimal care or treatment. Once more, an easily accessible reference work on the rhesus monkey anatomy would be extremely useful.
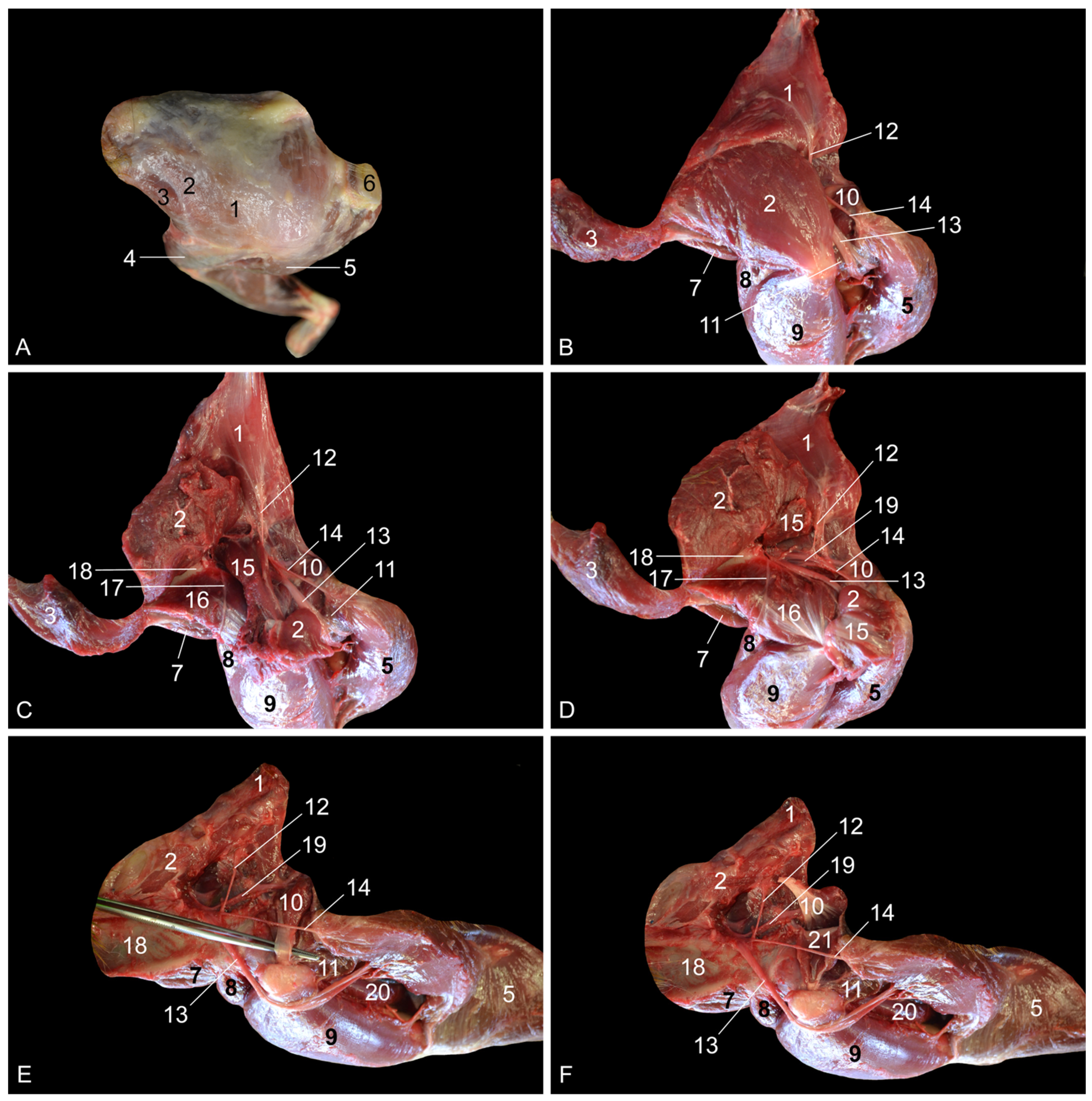
When the present findings were compared with the literature data on the rhesus monkey anatomy, a few contradictions were noticed. Our dissections revealed that the m. adductor brevis consists of two bellies in the rhesus monkey, whereas the reference works on the anatomy of the rhesus monkey mention the presence of only a single muscle belly [
7,
8]. These works also state that the v. saphena lateralis drains into the v. poplitea. In contrast, our results indicate that the v. saphena lateralis drains into the v. caudalis femoris, which in turn flows into the v. poplitea. In other words, the v. saphena lateralis does not directly join the v. poplitea. This is also the case in common domestic mammals [
13].
When comparing the anatomy of the rhesus monkey with that of common domestic mammals, a number of particularities caught our attention. First, superficial and deep digital flexors, m. flexor digitorum (digitalis in the horse) superficialis and m. flexor digitorum (digitalis in the horse) profundus, respectively, do not exist as such in the rhesus monkey. The m. flexor digitorum (longus) medialis, which has previously been denominated the m. flexor digitorum (longus) tibialis [
7]—hence the mention of “m. flexor digitorum (longus) medialis/tibialis” in
Section 3—can be regarded as the superficial digital flexor. In domestic mammals, the m. flexor digitorum/digitalis profundus is composed of the m. tibialis caudalis, the m. flexor digitorum/digitalis medialis and the m. flexor digitorum/digitalis lateralis. The latter muscle in the rhesus is the m. flexor digitorum (longus) lateralis, which has previously been termed the m. flexor digitorum (longus) fibularis, hence the mention of “m. flexor digitorum (longus) lateralis/fibularis” in
Section 3 [
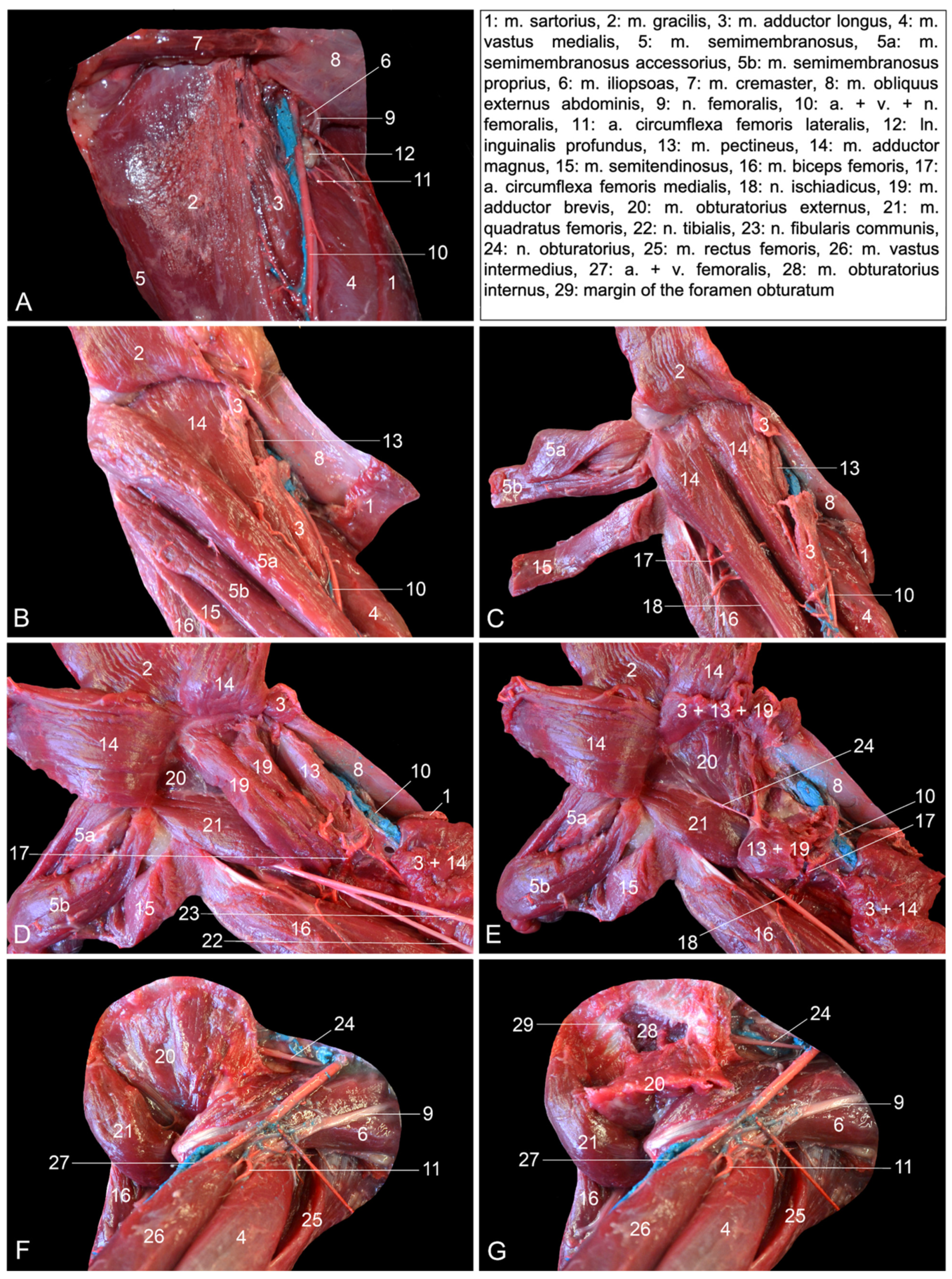
7]. It can be considered the deep digital flexor. The m. tibialis caudalis is a separate muscle in the rhesus monkey and is not one of the muscles that belong to the trigeminal deep digital flexor that is present in domestic mammals. Moreover, it is not a specific digital flexor, but rather a carpal flexor since its tendons insert into the plantar sides of the 2nd to 5th metatarsal bones and not the phalanges. Second, the rhesus monkey presents the m. adductor magnus, longus and brevis. This conformation of the adductor muscles is only seen in the cat. In other domestic mammals, the m. adductor longus has been fused with the m. pectineus [
14]. This might be the reason why the m. pectineus is that prominent is those species, while it is rather insignificant in the rhesus monkey and obscured by the m. adductor longus when the hip region is approached medially. Third, the m. semimembranosus consists of two muscle bellies in the rhesus monkey. The smaller and more lateral semimembranosus proprius muscle is inserted medially on the tibial tuberosity, while the larger and more medial semimembranosus accessorius muscle is broadly inserted more proximally, at the level of the medial femoral condyle. This configuration is also seen in the dog [
14].
The assessment of similarities and differences between the anatomy of the rhesus monkey and that of a human has been completed at the level of the musculature, the circulatory system, including the arteries and veins, and the nervous system.
Table 2 presents the major differences between rhesus monkeys and humans.
First, some peculiarities regarding the muscles are discussed. In the hip region of the rhesus monkey, the m. gluteus medius is more voluminous compared to the m. gluteus superficialis and the m. gluteus profundus. The former is by far the larger out of the three gluteal muscles in humans. Hence, the term m. gluteus maximus is applied [
16]. The deep gluteal muscle is denominated m. gluteus minimus in humans and can be absent in this species [
8,
14,
16]. The m. tensor fasciae latae is more voluminous in humans compared to the rhesus monkey [
8,
16]. The m. psoas minor seems constant in the rhesus monkey, whereas it can be absent in humans [
7,
8,
16]. Both in the rhesus monkey and domestic mammals, the mm. gemelli represent a single muscle, in contrast to what the anatomical term suggests [
14]. This term has been adopted from human anatomy, in which the m. gemellus superior and inferior exist [
16]. However, one or both mm. gemelli can be absent in humans [
7,
16].
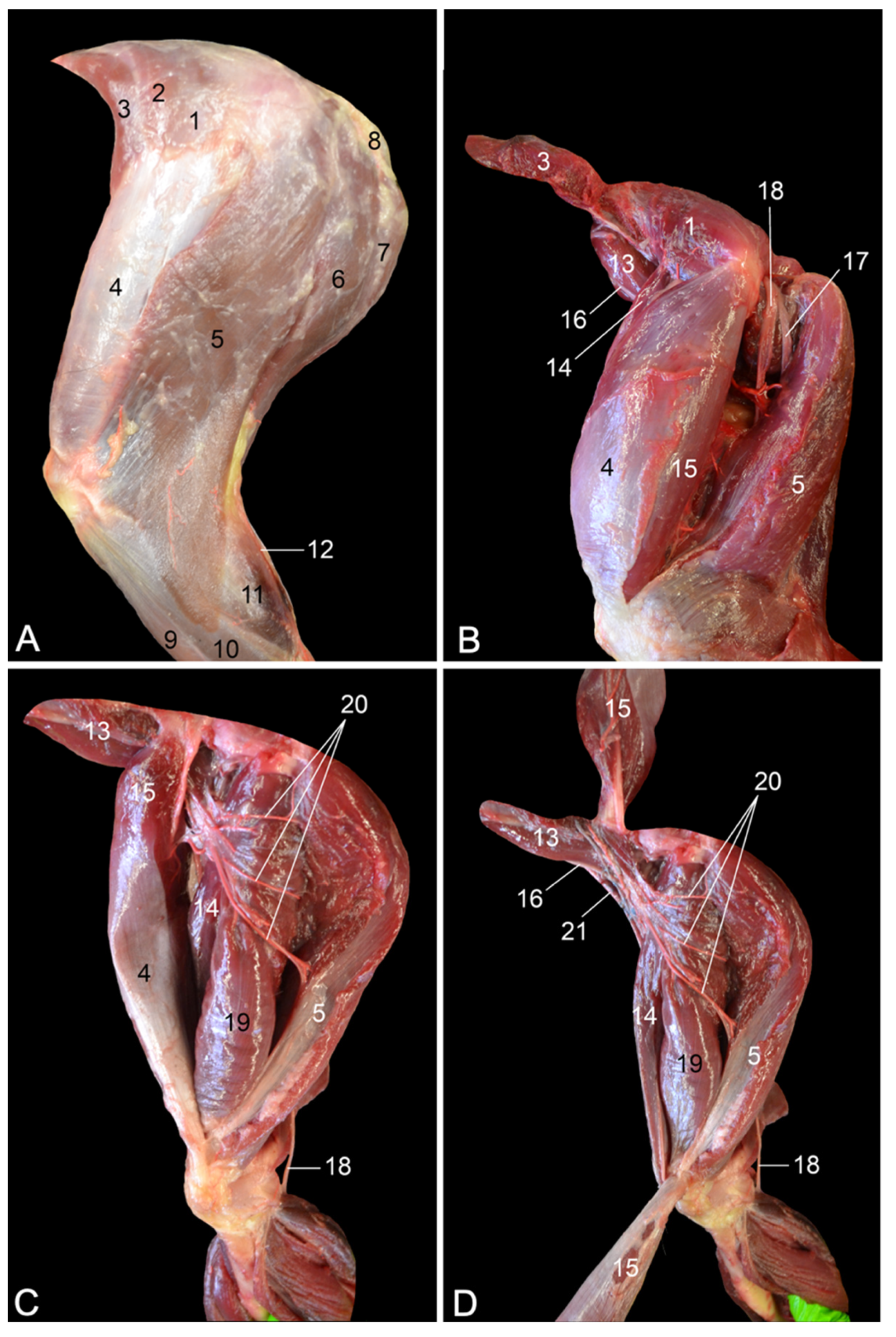
At the level of the upper limb, both the m. rectus femoris and the m. biceps femoris consist of two heads in humans and only a single head in the rhesus monkey [
7,
8,
16]. Comparable to humans, the equine m. rectus femoris presents a medial and a lateral head. Surprisingly, the equine m. biceps femoris does not contain a single head as in other domestic mammals, nor two heads as in humans—hence the anatomical term biceps—but three heads [
14]. The single m. semimembranosus in the horse is partly fused with the m. adductor magnus [
14]. In humans, it is the m. semimembranosus accessorius that is fused with m. adductor magnus [
7,
8,
16]. Finally, the human m. pectineus can be composed of two muscle bellies, while only one muscle belly can be discerned in the rhesus monkey [
7,
16].
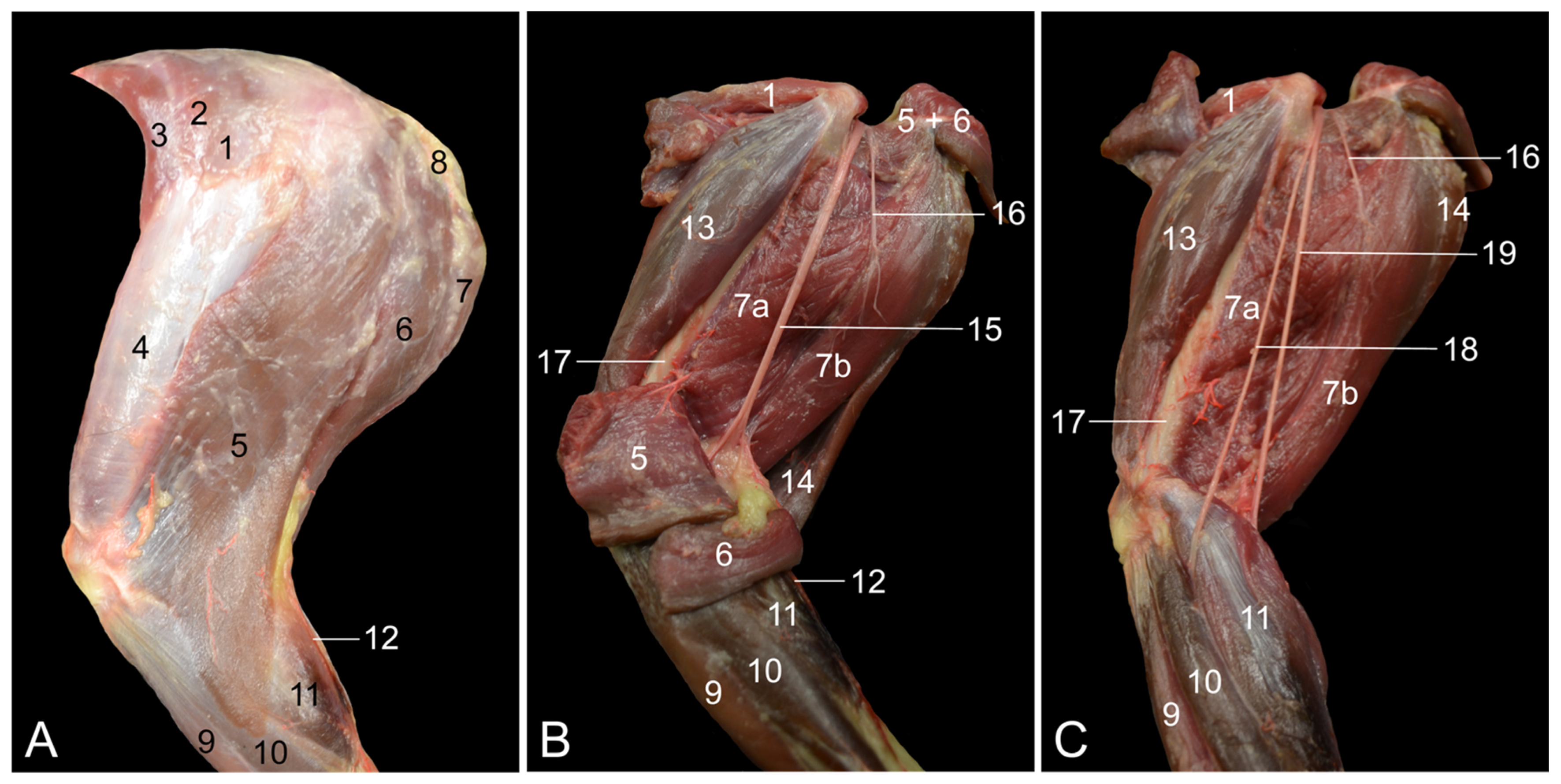
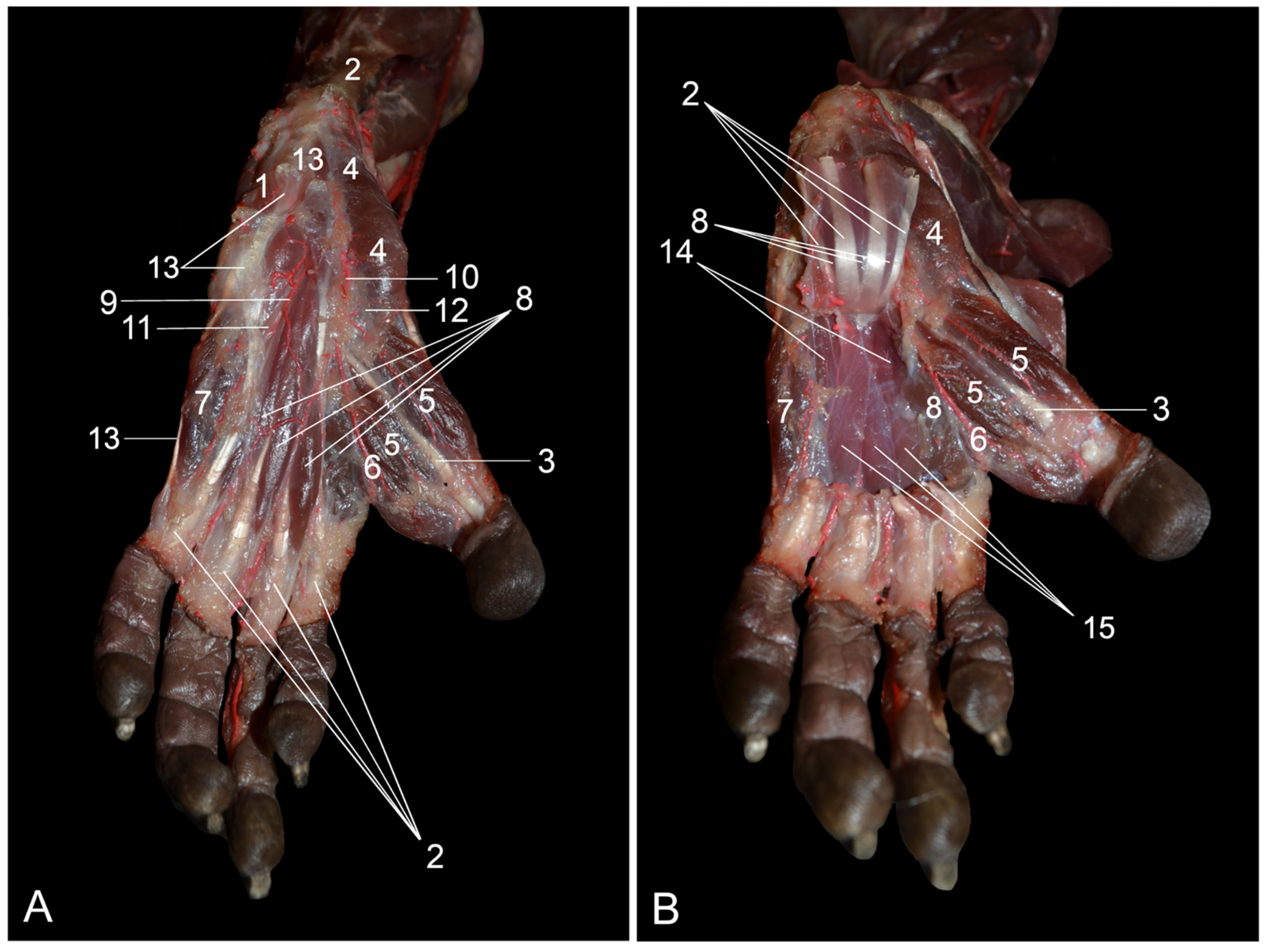
The m. tibialis cranialis is located on the cranial side of the lower leg. In contrast to humans, in which the m. tibialis anterior is most often single, the m. tibialis cranialis of the rhesus monkey has two muscle bellies [
7,
8]. The human m. fibularis longus sends a tendon to the first tarsal bone in addition to the tendon to the first metatarsal bone, which is the sole tendon in the rhesus monkey [
7,
8]. Both the rhesus monkey and humans possess the m. fibularis brevis, but a third fibularis muscle, i.e., the m. fibularis tertius—hence its name—is present in human beings. Its tendon inserts into the base of the fifth metatarsal bone [
16]. It is therefore analogous to the m. fibularis digiti quinti in the rhesus monkey [
7]. The presence of the m. fibularis tertius is unique to the horse, which, in contrast to other domestic mammals, lacks the other fibularis muscles [
14]. The m. soleus, which, among domestic mammals, can also be identified in the cat and rabbit, consists of two heads in humans and only one in the rhesus monkey, like in the cat and the rabbit [
7,
8,
14,
16]. The m. plantaris is more pronounced in the rhesus monkey in comparison with humans [
7,
8]. The m. flexor digitorum (longus) lateralis/fibularis is, in human anatomy, denoted as the m. flexor hallucis longus since its single tendon inserts into the plantar side of the distal phalanx of the first toe [
16], with no additional tendons to the third and fourth toes as seen in the rhesus monkey [
7]. Furthermore, the m. flexor digitorum (longus) medialis/tibialis of the rhesus monkey is known as the m. flexor digitorum longus in humans, since this muscle only sends tendons to the 2nd to 5th toes in that species and not to all digits as in the rhesus monkey [
7,
16].
At the level of the foot, the human m. quadratus plantae comprises two heads instead of one in the rhesus monkey [
7,
8,
16]. The m. flexor digitorum brevis only contains a superficial part in humans. The additional deep part that is seen in the rhesus monkey is absent [
7,
8,
16]. Finally, the mm. contrahentes digitorum pedis are absent in the human foot [
16].
The observed differences in the myology of the hindlimb between humans and rhesus monkeys are a reflection of the orthograde posture and associated bipedalism in humans vs. the pronograde posture and associated quadrupedalism in the rhesus monkey [
7]. This results in a different position of the center of mass between the two species and in a dissimilar arrangement of the hallux, which, in the rhesus monkey, sits more medially than the anterior human hallux [
16].
Besides differences in musculature, the branching pattern of the arterial system of the rhesus monkey slightly deviates from that in humans. In humans, rhesus monkeys and domestic mammals, the a. profunda femoris branches off the proximal segment of the a. femoralis. In humans, the a. profunda femoris gives off the a. circumflexa femoris medialis. This is a typical trait in domestic mammals [
13]. Surprisingly, the a. circumflexa femoris medialis of the rhesus monkey is not given off by the a. profunda femoris, but by the a. obturatoria [
7]. The a. circumflexa femoris lateralis branches off the a. profunda femoris in both the rhesus monkey and humans [
16]. In contrast, the a. circumflexa femoris lateralis is a branch of the a. femoralis in domestic mammals [
13].
At the level of the knee, the human femoral artery continues as the popliteal artery, which gives off the a. genus descendens [
16]. A similar configuration is seen in domestic mammals [
12]. This a. genus descendens is comparable to the a. genus proximalis that is described in the rhesus monkey [
7,
8]. However, this artery is a branch of the a. saphena, which is given off by the femoral artery. After the a. saphena has branched off, the femoral artery continues as the popliteal artery. Thus, the artery that supplies the craniolateral knee region of the rhesus monkey is not a direct branch of the femoral artery, but a branch of the a. saphena.
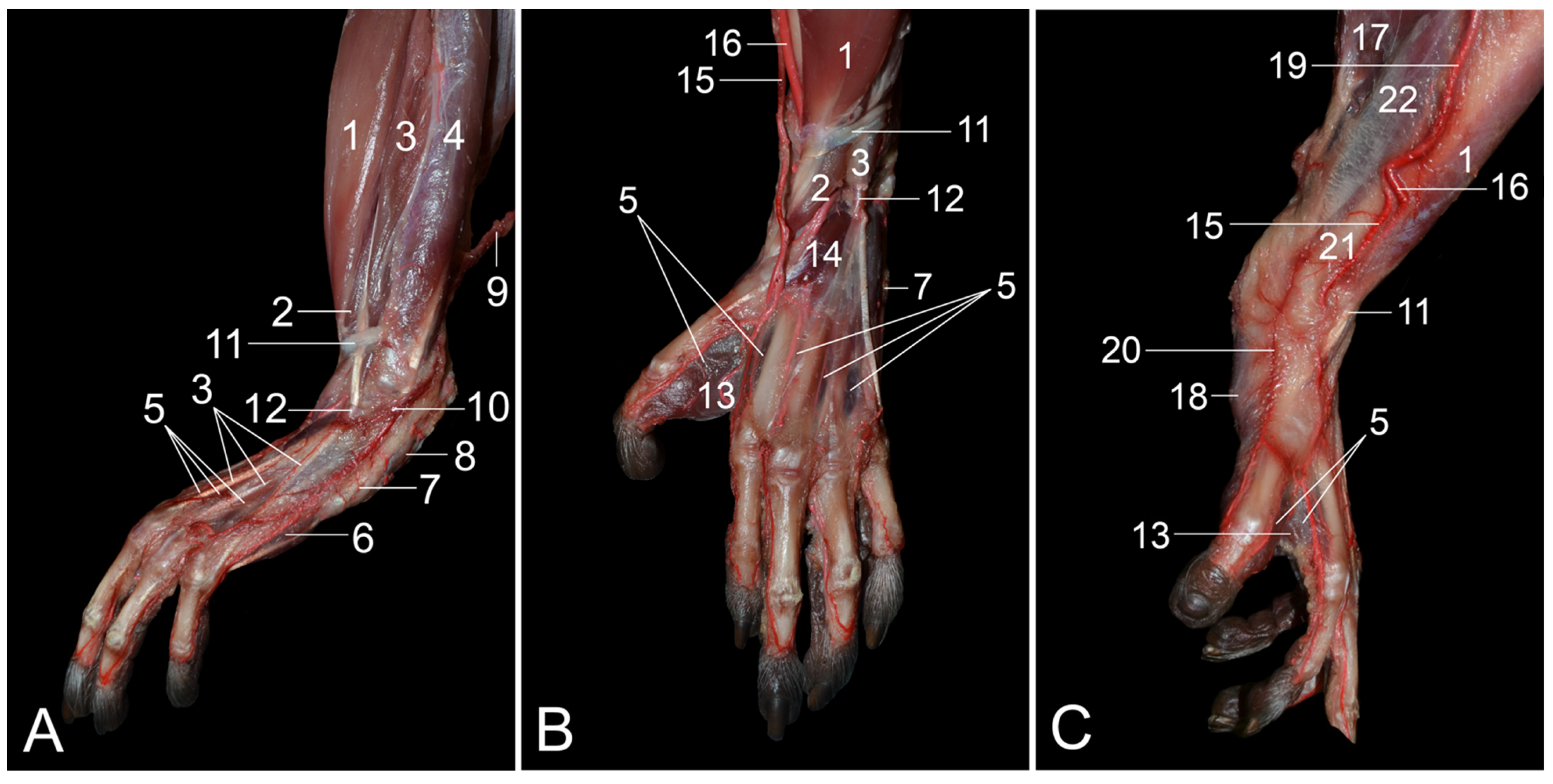
The a. saphena is very important in the rhesus monkey, as it plays a pivotal role in the blood supply of the foot, together with the a. tibialis caudalis, which is regarded as the continuation of the popliteal artery in this species [
7]. In contrast, the a. saphena is insignificant in humans, in which it is a tiny branch of the a. genus descendens that joins the n. saphenus. It can even be absent in some individuals [
7,
8,
16]. Consequently, the human foot is supplied with blood via the a. tibialis anterior and the a. tibialis posterior [
16]. In domestic mammals, the continuation of the popliteal artery is the a. tibialis cranialis. which provides blood to the dorsal side of the foot (a. dorsalis pedis). The plantar side of the foot, but also the dorsal side to some extent, is supplied by the vital a. saphena [
13]. Also in humans, the a. dorsalis pedis is in continuation with the a. tibialis anterior [
16]. This contrasts with the rhesus monkey, in which the a. dorsalis pedis (both superficialis and profundus) branches off the a. saphena [
7,
8].
The a. saphena has a venous counterpart called the v. saphena. In fact, the v. saphena is dual. The v. saphena parva in humans drains into the v. poplitea [
16]. In mammals, including the rhesus monkey, the v. saphena lateralis first flows into the v. caudalis femoris, which in turn drains into the v. poplitea [
13]. The v. caudalis femoris is absent in humans [
16]. The v. saphena parva is of clinical importance in the rhesus monkey since it allows for intravenous catheterization or blood collection [
9,
17].
The other saphenous vein is the v. saphena magna in humans or v. saphena medialis in mammals, including the rhesus monkey. Irrespective of the species, this vein drains into the femoral vein [
17,
18]. The v. saphena medialis in the rhesus monkey is a paired vein that flanks the a. saphena and, as such, is denominated the vv. comitantes (cum a. saphena). Vv. comitantes were also noticed along the a. tibialis caudalis (vv. comitantes cum a. tibialis caudalis) [
17,
18]. In humans, vv. comitantes are described alongside deep arteries, but not adjacent to superficial arteries such as the a. saphena [
16].
The human terms v. saphena magna and v. saphena parva could be mystifying in the rhesus monkey. Since “magna” means “big, large”, the term v. saphena magna is chosen for that saphenous vein, which is larger in humans. The smaller saphenous vein is termed the v. saphena parva, with “parva” being Latin for “small, tiny”. In the rhesus monkey, however, it is more appropriate to speak of the v. saphena medialis instead of the v. saphena magna and to use the term v. saphena lateralis and not v. saphena parva because, in this species, not the medial but the lateral saphenous vein is larger.
The observed differences between the rhesus monkey and humans regarding the conformation of the nerves are rather trivial. In humans, the n. ischiadicus divides into the n. fibularis communis and the n. tibialis just proximal to the knee [
16]. In contrast, the n. ischiadicus is very short in the rhesus monkey. The n. fibularis communis and the n. tibialis can already be recognized at the level of the hip joint after their common epineurium has been incised longitudinally [
7,
8]. The n. cutaneus surae medialis (known as n. cutaneus surae caudalis in veterinary anatomy) and the n. cutaneus surae lateralis communicate in humans [
13,
16]. More distally, both nerves join to form the n. suralis [
16]. In the rhesus monkey and domestic mammals, no communication is present between the two nerves, and the n. suralis is absent [
7,
15]. Finally, the m. pectineus is innervated uniquely by the n. femoralis in the rhesus monkey [
7], whereas this muscle is additionally innervated by the n. obturatorius in humans [
7,
16].















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}