




Development of a Real-Time TaqMan RT-PCR Assay for the Detection of NADC34-like Porcine Reproductive and Respiratory Syndrome Virus
 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Strains and Clinical Samples
2.2. DNA/RNA Extraction Method
2.3. Construction of the ORF5 Gene Plasmid
2.4. Primers and Probe for NADC34-like PRRSV ORF5 Gene
2.5. Cloning and Sequencing
2.6. Optimization of the Real-Time RT-PCR
2.7. Method Standardization
2.8. Specificity Analysis
2.9. Sensitivity Analysis
2.10. Repeatability and Reproducibility Analyses
2.11. Detection of NADC34-like PRRSV in Clinical Samples
3. Results
3.1. Optimization Results of Real-Time RT-qPCR System
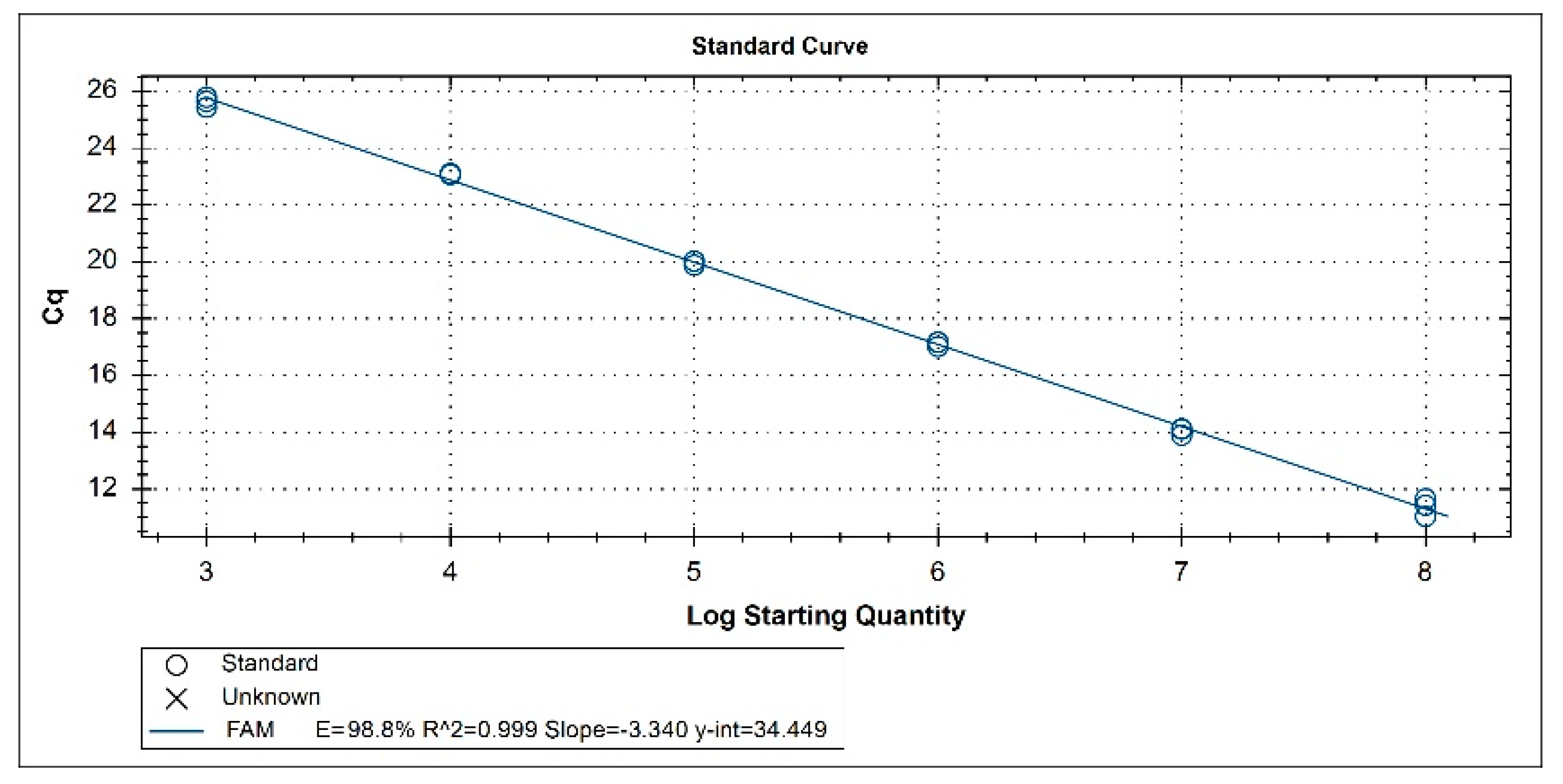
3.2. Standard Curve
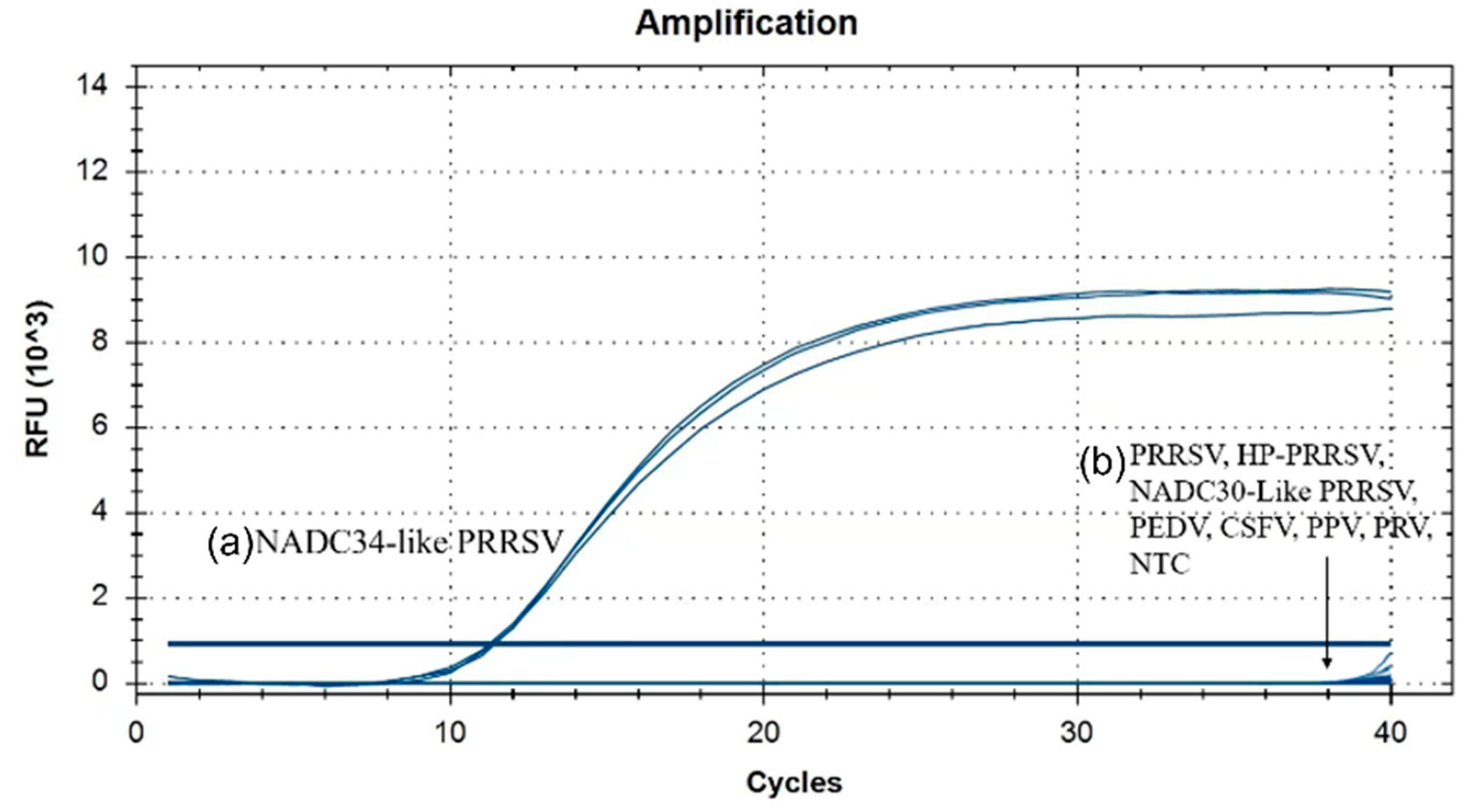
3.3. Specificity Analysis
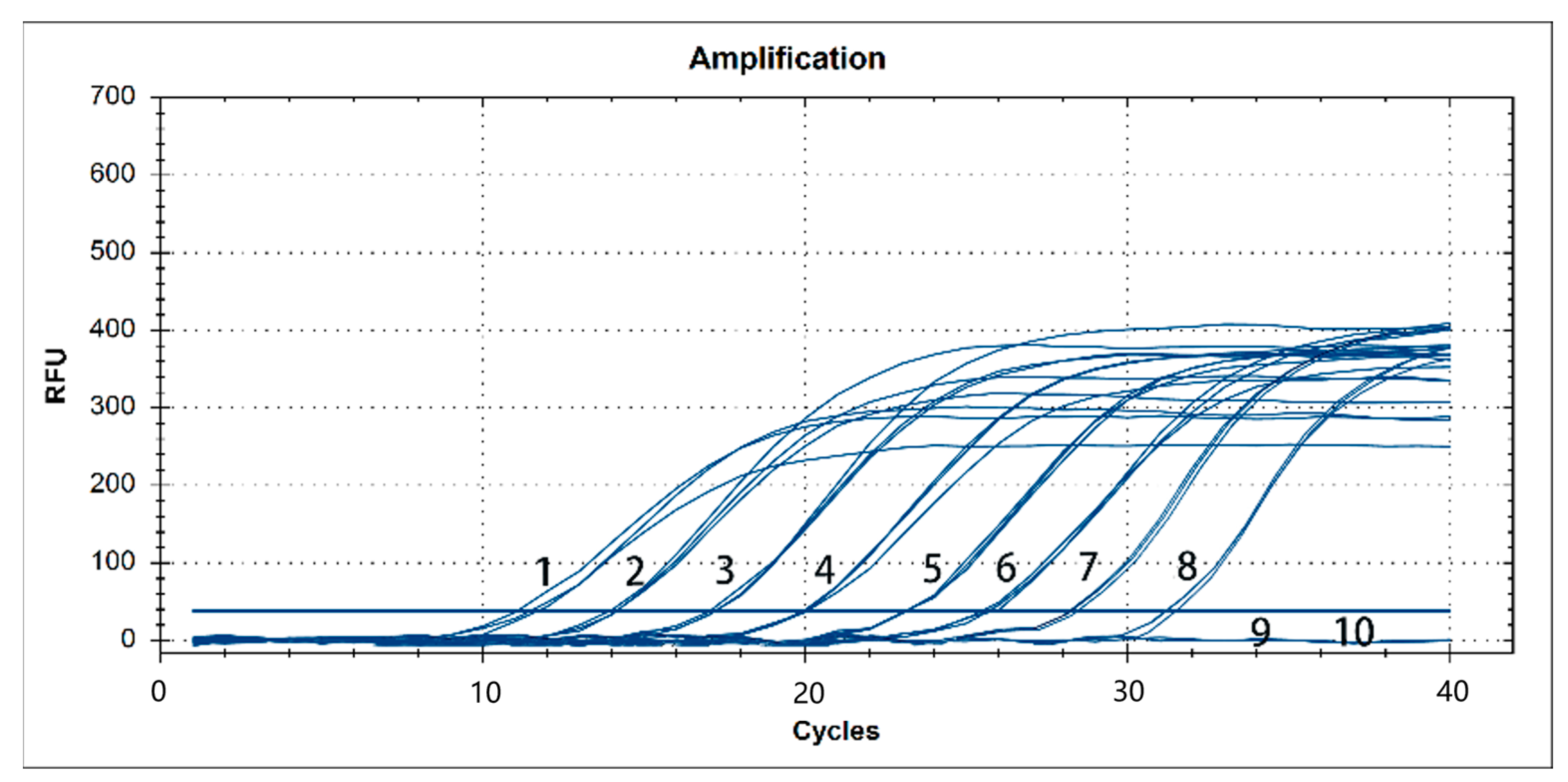
3.4. Sensitivity Analysis
3.5. Reproducibility Analysis
3.6. Detection of NADC34-like PRRSV in the Clinical Samples
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Woo, P.C.Y.; Lau, S.K.P.; Huang, Y.; Yuen, K. Coronavirus diversity, phylogeny and interspecies jumping. Exp. Biol. Med. 2009, 234, 1117–1127. [Google Scholar] [CrossRef] [Green Version]
- Mardassi, H.; Massie, B.; Dea, S. Intracellular synthesis, processing, and transport of proteins encoded by ORFs 5 to 7 of porcine reproductive and respiratory syndrome virus. Virology 1996, 221, 98–112. [Google Scholar] [CrossRef] [Green Version]
- Dokland, T. The structural biology of PRRSV. Virus Res. 2010, 154, 86–97. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.J.; Lefkowitz, E.J.; King, A.M.Q.; Harrach, B.; Harrison, R.L.; Knowles, N.J. Changes to taxonomy and the international code of virus classification and nomenclature ratified by the international committee on taxonomy of viruses (2017). Arch. Virol. 2017, 162, 2505–2538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, H.; Jung, K.; Vlasova, A.; Chepngeno, J.; Lu, Z.; Wang, Q.; Saif, L. Isolation and characterization of porcine deltacoronavirus from pigs with diarrhea in the United States. J. Clin. Microbiol. 2015, 53, 1537–1548. [Google Scholar] [CrossRef] [Green Version]
- Fenner, F. Classification and nomenclature of viruses. SummarizedF proceedings of International Committee for Virus Taxonomy. Ann. Microbiol. 1976, 127, 323. [Google Scholar]
- Yun, S.I.; Lee, Y.M. Overview: Replication of porcine reproductive and respiratory syndrome virus. J. Microbiol. 2013, 51, 711–717. [Google Scholar] [CrossRef]
- He, W.; Ji, X.; He, W.; Dellicour, S.; Wang, S.; Li, G.; Zhang, L.; Gilbert, M.; Zhu, H.; Xing, G.; et al. Genomic epidemiology, evolution, and transmission dynamics of porcine deltacoronavirus. Mol. Biol. Evol. 2020, 37, 2641–2654. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Treffers, E.E.; Li, Y. Efficient -2 frameshifting by mammalian ribosomes to synthesize an additional arterivirus protein. Proc. Natl. Acad. Sci. USA 2012, 109, E2920–E2928. [Google Scholar] [CrossRef] [Green Version]
- Ostrowski, M.; Galeota, J.A.; Jar, A.M.; Platt, K.B.; Osorio, F.A.; Lopez, O.J. Identification of neutralizing and nonneutralizing epitopes in the porcine reproductive and respiratory syndrome virus GP5 ectodomain. J. Virol. 2002, 76, 4241–4250. [Google Scholar] [CrossRef] [Green Version]
- Roques, E.; Girard, A.; Stlouis, M.C.; Massie, B.; Gagnon, C.; Lessard, M.; Archambault, D. Immunogenic and protective properties of GP5 and M structural proteins of porcine reproductive and respiratory syndrome virus expressed from replicating but nondisseminating adenovectors. Vet. Res. 2013, 44, 17. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Frey, M.L.; Yoon, K.J.; Zimmerman, J.J.; Platt, K.B. Categorization of North American porcine reproductive and respiratory syndrome viruses: Epitopic profiles of the N, M, GP5 and GP3 proteins and susceptibility to neutralization. Arch. Virol. 2000, 145, 1599–1619. [Google Scholar] [CrossRef]
- Hicks, J.A.; Yoo, D.; Liu, H.C. Interaction of porcine reproductive and respiratory syndrome virus major envelope proteins GP5 and M with the cellular protein Snapin. Virus Res. 2018, 249, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Jiang, P.; Li, Y.; Tang, J.; Wang, X.; Ma, S. Recombinant adenovirus expressing GP5 and M fusion proteins 185 of porcine reproductive and respiratory syndrome virus induce both humoral and cell-mediated immune responses in mice. Vet. Immunol. Immunopathol. 2006, 113, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Chen, S.; Zhang, J.; Zeng, J.; Guo, X.; Ge, X.; Zhang, D.; Yang, H. Molecular variation analysis of porcine reproductive and respiratory syndrome virus in China. Virus Res. 2009, 145, 97–105. [Google Scholar] [CrossRef]
- Shi, M.; Lam, T.T.Y.; Hon, C.C.; Murtaugh, M.P.; Davies, P.; Hui, R.K.H.; Wong, L.T.; Yip, C.; Jiang, J.; Leung, F.C. Phylogeny-based evolutionary, demographical, and geographical dissection of North American type 2 porcine reproductive and respiratory syndrome viruses. J. Virol. 2010, 84, 8700–8711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, B.; Wang, H.; Bai, J.; Zhang, L.; Jiang, P. A novel isolate with deletion in GP3 gene of porcine reproductive and respiratory syndrome virus from mid-eastern China. BioMed Res. Int. 2014, 2014, 3016130. [Google Scholar] [CrossRef] [Green Version]
- Pirzadeh, B.; Gagnon, C.A.; Dea, S. Genomic and antigenic variations of porcine reproductive and respiratory syndrome virus major envelope GP5 glycoprotein. Can. J. Vet. Res. 1998, 62, 170. [Google Scholar]
- Adams, M.J.; Lefkowitz, E.J.; King, A.M.Q.; Harrach, B.; Harrison, R.; Knowles, N.J.; Kropinski, A.M.; Krupovic, M.; Kuhn, J.H.; Mushegian, A.R.; et al. Ratification vote on taxonomic proposals to the International Committee on Taxonomy of Viruses. Arch. Virol. 2016, 161, 2921–2949. [Google Scholar] [CrossRef] [Green Version]
- Brar, M.S.; Shi, M.; Murtaugh, M.P.; Leung, F.C.C. Evolutionary diversification of type 2 porcine reproductive and respiratory syndrome virus. J. Gen. Virol. 2015, 96, 1570–1580. [Google Scholar] [CrossRef]
- Van, A.G.M.; Anderson, T.K.; Lager, K.M.; Das, P.; Otis, N.; Montiel, N.; Miller, L.C.; Kulshreshtha, V.; Buckley, A.; Brockmeier, S.; et al. Porcine reproductive and respiratory disease virus: Evolution and recombination yields distinct ORF5 RFLP 1-7-4 viruses with individual pathogenicity. Virology 2017, 51, 168–179. [Google Scholar] [CrossRef]
- Zhang, H.L.; Zhang, W.L.; Xiang, L.R.; Leng, C.; Tian, Z.; Tang, Y.; Cai, X. Emergence of novel porcine reproductive and respiratory syndrome viruses (ORF5 RFLP 1-7-4 viruses) in China. Vet. Microbiol. 2018, 222, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Song, S.J.; Zhao, J.; Leng, C.; Fu, J.; Li, C.; Tang, Y.; Xiang, L.; Peng, J.; Wang, Q.; et al. A potential endemic strain in China: NADC34-like porcine reproductive and respiratory syndrome virus. Transbound Emerg. Dis. 2020, 67, 1730–1738. [Google Scholar] [CrossRef] [PubMed]
- Long, Y.P.; Zhou, J.; Xiao, L.; Yu, J.; Zhang, B.; Zhou, L. Whole genome sequence analysis of a NADC34-like porcine reproductive and respiratory syndrome virus. Chin. J. Prev. Vet. Med. 2022, 44, 544–549+554. [Google Scholar] [CrossRef]
- Qiu, W.; Meng, K.; Liu, Y.; Zhang, Y.; Wang, Z.; Chen, Z.; Yang, J.; Sun, W.; Guo, L.; Ren, S.; et al. Simultaneous detection of classical PRRSV, highly pathogenic PRRSV and NADC30-like PRRSV by TaqMan probe real-time PCR. J. Virol. Methods 2020, 282, 113774. [Google Scholar] [CrossRef]
- Kappes, M.A.; Faaberg, K.S. PRRSV structure, replication and recombination: Origin of phenotype and genotype diversity. Virology 2015, 479, 475–486. [Google Scholar] [CrossRef] [Green Version]
- Guo, B.; Chen, Z. PRRSV isolation from aborted fetuses suspected of PRRS. Chin. J. Prev. Vet. Med. 1996, 87, 1–4. [Google Scholar]
- Yang, H.; Huang, F.; Guo, X. Sequencing of Genome of Porcine Reproductive and Respiratory Syndrome Virus isolate bj-4. Chin. J. Agric. Biotechol. 2001, 3, 212–218. [Google Scholar]
- Tian, K.; Yu, X.; Zhao, T.; Feng, Y.; Cao, Z.; Wang, C.; Hu, Y.; Chen, X.; Hu, D.; Tian, X.; et al. Emergence of fatal PRRSV variants: Unparalleled outbreaks of atypical PRRS in China and molecular dissection of the unique hall mark. PLoS ONE 2007, 2, e526. [Google Scholar] [CrossRef]
- Lu, H.; Tun, M.; Sun, L.; Mo, J.; Zhou, Q.; Deng, Y.; Xie, Q.; Bi, Y.; Leung, C.; Ma, J. Re-emerging of porcine respiratory and reproductive syndrome virus (lineage 3) and increased pathogenicity after genomic recombination with vaccine variant. Vet. Microbiol. 2015, 175, 332–340. [Google Scholar] [CrossRef]
- Liu, J.; Wei, C.; Lin, Z.; Xia, W.; Ma, Y.; Dai, A.; Yang, X. Full genome sequence analysis of a 1-7-4-like PRRSV strain in Fujian province. China Peer J. 2019, 7, e7859. [Google Scholar] [CrossRef]
- Song, S.; Xu, H.; Zhao, J.; Leng, C.; Xiang, L.; Li, C.; Fu, J.; Tang, Y.; Peng, J.; Wang, Q.; et al. Pathogenicity of NADC34-like PRRSV HLJDZD32-1901 isolated in China. Vet. Microbiol. 2020, 246, 108727. [Google Scholar] [CrossRef]
- Li, Y.L.; Ji, G.B.; Xu, X.D.; Wang, J.; Li, X. Development and application of an RT-PCR to differentiate the prevalent NAPRRSV strains in China. Open Virol. J. 2017, 11, 66–77. [Google Scholar] [CrossRef] [Green Version]
- Chen, N.H.; Ye, M.X.; Xiao, Y.Z.; Li, S.; Zhu, J. Development of universal and quadruplex real-time RT-PCR assays forsimultaneous detection and differentiation of porcine reproductive and respiratory syndrome viruses. Transbound Emerg. Dis. 2019, 66, 2271–2278. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Zhao, T.; Liang, W.; Song, W.; Gao, Z.; Tang, X.; Chen, H.; Wu, B. RT-PCR detection of porcine reproductive and respiratory syndrome virus based on the ORF5 gene in Mainland China, 2012–2015. Acta Virol. 2017, 61, 336–340. [Google Scholar] [CrossRef] [PubMed]
- Liang, W.; Zhao, T.; Peng, Z.; Sun, Y.; Stratton, C.W.; Zhou, D.; Tang, X.; Tian, Y.; Chen, H.; Wu, B. Epidemiological and genetic characteristics of porcine reproductive and respiratory syndrome virus circulating in central and South China in 2016. Acta Trop. 2019, 190, 83–91. [Google Scholar] [CrossRef]
- Chen, N.; Li, X.; Xiao, Y.; Li, S.; Zhu, J. Characterization of four types of MLV-derived porcine reproductive and respiratory syndrome viruses isolated in unvaccinated pigs from 2016 to 2020. Res. Vet. Sci. 2021, 134, 102–111. [Google Scholar] [CrossRef]
- Zhou, L.; Yang, Y.; Xia, Q.; Guan, Z.; Zhang, J.; Li, B.; Qiu, Y.; Liu, K.; Shao, D.; Ma, Z.; et al. Genetic characterization of porcine reproductive and respiratory syndrome virus from Eastern China during 2017–2022. Front. Microbiol. 2022, 13, 971817. [Google Scholar] [CrossRef]
- Fang, K.; Liu, S.; Li, X.; Chen, H.; Qian, P. Epidemiological and Genetic Characteristics of Porcine Reproductive and Respiratory Syndrome Virus in South China Between 2017 and 2021. Front. Vet. Sci. 2022, 9, 853044. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Kang, R.; Zhang, Y.; Ding, M.; Xie, B.; Tian, Y.; Wu, X.; Zuo, L.; Yang, X.; Wang, H. Whole Genome Analysis of Two Novel Type 2 Porcine Reproductive and Respiratory Syndrome Viruses with Complex Genome Recombination between Lineage 8, 3, and 1 Strains Identified in Southwestern China. Viruses 2018, 10, 328. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Qu, X.; Zhang, H.; Tang, X.; Bian, T.; Sun, Y.; Zhou, M.; Ren, F.; Wu, P. Evolutionary and recombination analysis of porcine reproductive and respiratory syndrome isolates in China. Virus Genes 2020, 56, 354–360. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Type of Virus | Primers and Probe(5′-3′) | Target Gene |
|---|---|---|
| NADC34-like PRRSV | F: CCTGTGTGACTCATATTGTCTCC R: CGGCGTAAATGCTACTCAAGAC P: FAM-CGCCCTCACCACCAGCCATTTCCT-BHQ1 | ORF5 |
| HP- PRRSV | F: GACGTGCCCCCAAGCTGAT | Nsp2 |
| R: GGATGCCCATGTTCTGCGA P: FAM-CGTAGAACTGTGACAACAACGCTGAC-BHQ1 | ||
| NADC30-like-PRRSV | F: CGTATTGGACACCTCTTTTGACTG R: AACTGGACCTAATCTTCCTGCG P: ROX-CCCAAAGGTCTTCGTCGGTATTCC-BHQ2 | Nsp2 |
| Positive Plasmid Concentration | Intra-Assay | Inter-Assay | ||
|---|---|---|---|---|
| (copies/µL) | Ct (Mean ± SD) | CV (%) | Ct (Mean ± SD) | CV (%) |
| 7.80 × 105 | 19.98 ± 0.11 | 0.55 | 20.04 ± 0.15 | 0.75 |
| 7.80 × 104 | 23.09 ± 0.13 | 0.56 | 23.11 ± 0.21 | 0.91 |
| 7.80 × 103 | 25.65 ± 0.21 | 0.82 | 25.80 ± 0.36 | 1.40 |
| 7.80 × 102 | 28.23 ± 0.25 | 0.89 | 28.45 ± 0.31 | 1.09 |
| Year | Sample Number | NADC34-like PRRSV | NADC30-like PRRSV | HP-PRRSV | NADC34-like PRRSV + NADC30-like PRRSV | NADC30-like PRRSV + HP-PRRSV | NADC34-like PRRSV + HP-PRRSV | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Number | Ratio | Number | Ratio | Number | Ratio | Number | Ratio | Number | Ratio | Number | Ratio | ||
| 2018 | 62 | 0 | 0 | 2 | 3.23% | 11 | 17.74% | 0 | 0 | 1 | 1.61% | 0 | 0 |
| 2019 | 53 | 0 | 0 | 5 | 9.43% | 14 | 26.42% | 0 | 0 | 3 | 5.66% | 0 | 0 |
| 2020 | 60 | 0 | 0 | 7 | 11.67% | 17 | 28.33% | 0 | 0 | 5 | 8.33% | 0 | 0 |
| 2021 | 57 | 0 | 0 | 5 | 8.77% | 14 | 24.56% | 0 | 0 | 5 | 8.77% | 0 | 0 |
| 2022 | 89 | 4 | 4.49% | 11 | 12.36% | 21 | 23.60% | 0 | 0 | 10 | 11.24% | 2 | 2.25% |
| Total | 321 | 4 | 1.24% | 30 | 9.35% | 77 | 23.99% | 0 | 0 | 24 | 7.48% | 2 | 0.62% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tu, T.; Pang, M.; Jiang, D.; Zhou, Y.; Wu, X.; Yao, X.; Luo, Y.; Yang, Z.; Ren, M.; Lu, A.; et al. Development of a Real-Time TaqMan RT-PCR Assay for the Detection of NADC34-like Porcine Reproductive and Respiratory Syndrome Virus. Vet. Sci. 2023, 10, 279. https://doi.org/10.3390/vetsci10040279
Tu T, Pang M, Jiang D, Zhou Y, Wu X, Yao X, Luo Y, Yang Z, Ren M, Lu A, et al. Development of a Real-Time TaqMan RT-PCR Assay for the Detection of NADC34-like Porcine Reproductive and Respiratory Syndrome Virus. Veterinary Sciences. 2023; 10(4):279. https://doi.org/10.3390/vetsci10040279
Chicago/Turabian StyleTu, Teng, Maonan Pang, Dike Jiang, You Zhou, Xulong Wu, Xueping Yao, Yan Luo, Zexiao Yang, Meishen Ren, Aiping Lu, and et al. 2023. "Development of a Real-Time TaqMan RT-PCR Assay for the Detection of NADC34-like Porcine Reproductive and Respiratory Syndrome Virus" Veterinary Sciences 10, no. 4: 279. https://doi.org/10.3390/vetsci10040279
APA StyleTu, T., Pang, M., Jiang, D., Zhou, Y., Wu, X., Yao, X., Luo, Y., Yang, Z., Ren, M., Lu, A., Zhang, G., Yu, Y., & Wang, Y. (2023). Development of a Real-Time TaqMan RT-PCR Assay for the Detection of NADC34-like Porcine Reproductive and Respiratory Syndrome Virus. Veterinary Sciences, 10(4), 279. https://doi.org/10.3390/vetsci10040279