
Locomotor Behavior Analysis in Spinal Cord Injured Macaca radiata after Predegenerated Peripheral Nerve Grafting—A Preliminary Evidence

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Selection
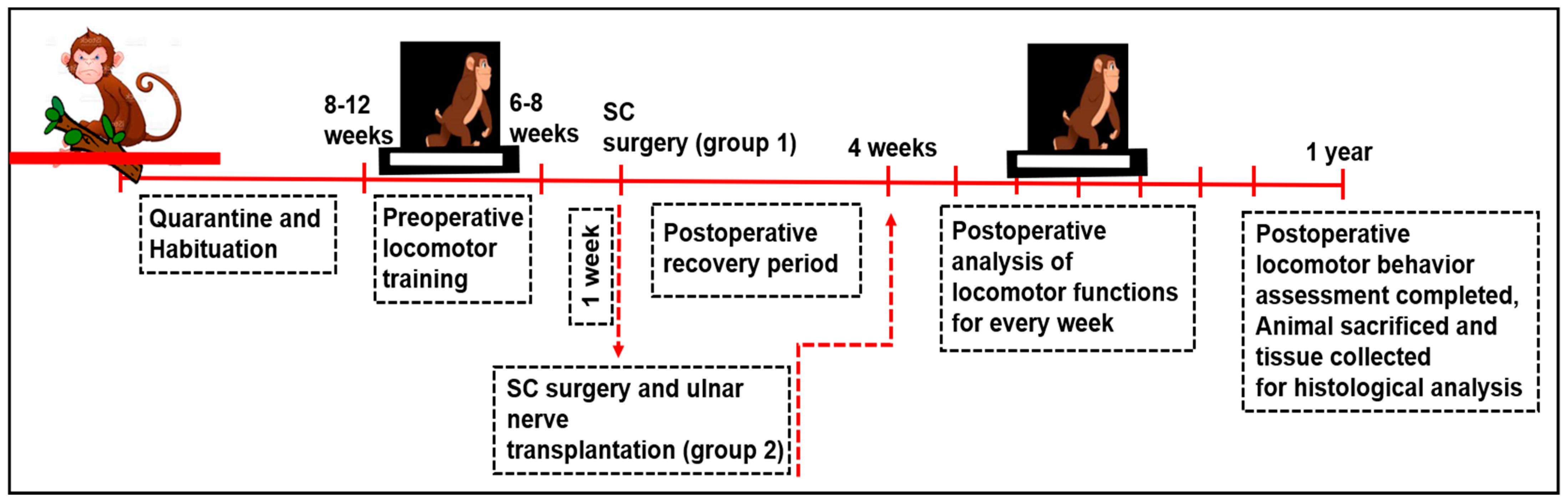
2.2. Experimental Design
2.3. The Locomotor Tests
2.4. Staircase Runway
2.5. Grid Runway
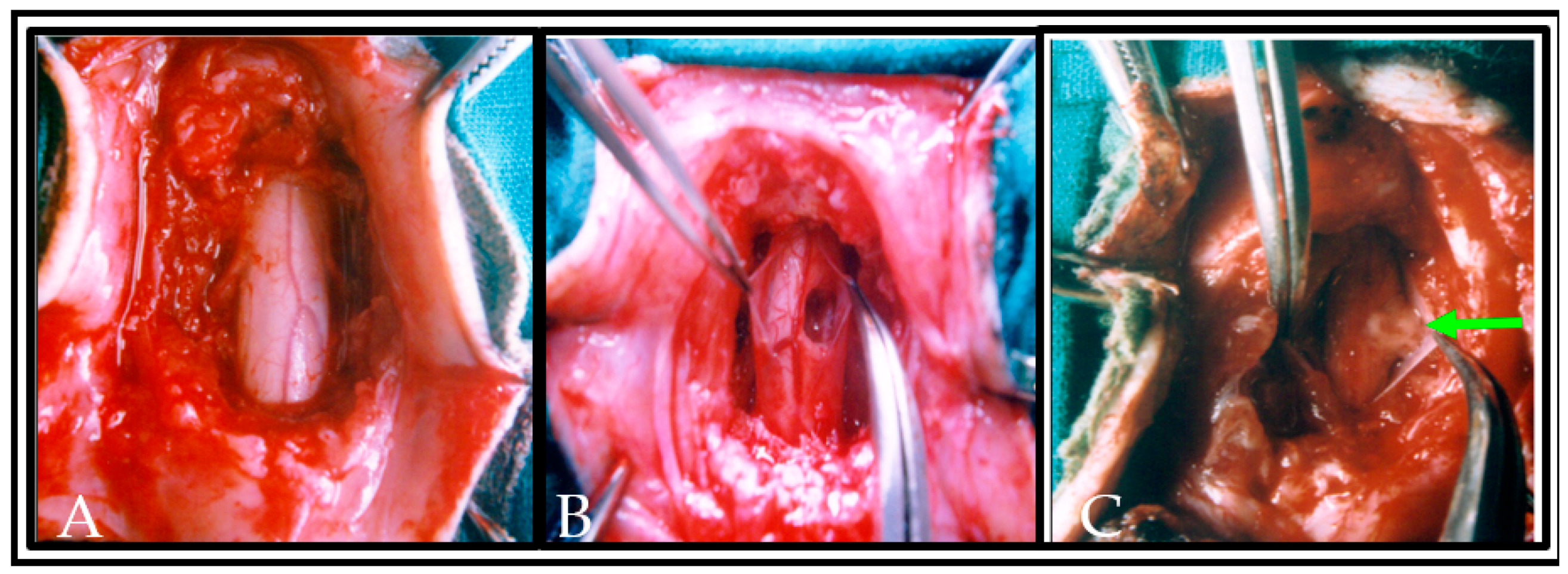
2.6. Spinal Cord Lesions
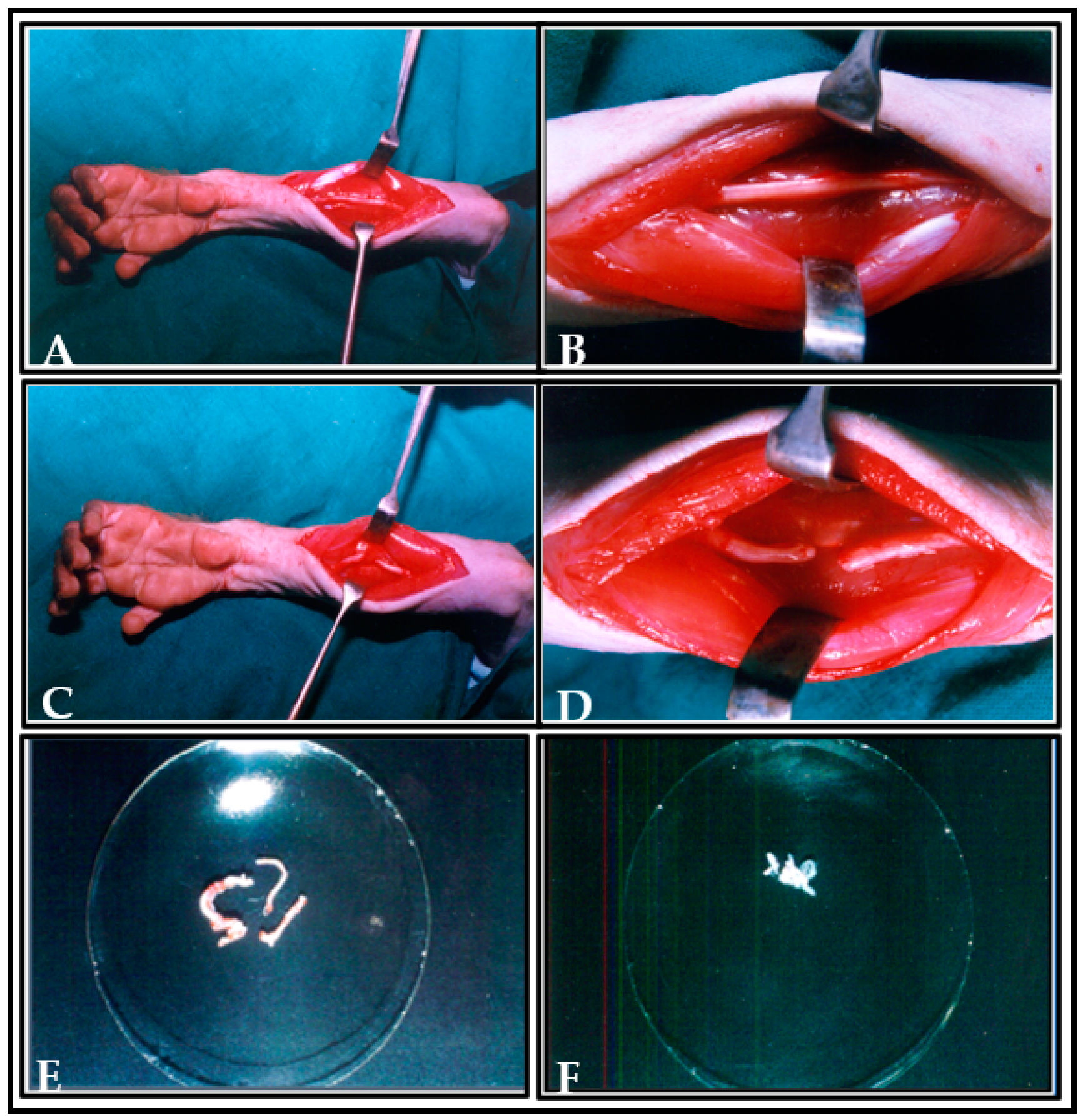
2.7. Preparation and Collection of Donor Tissue for Transplantation
2.8. Transplantation Animal Model
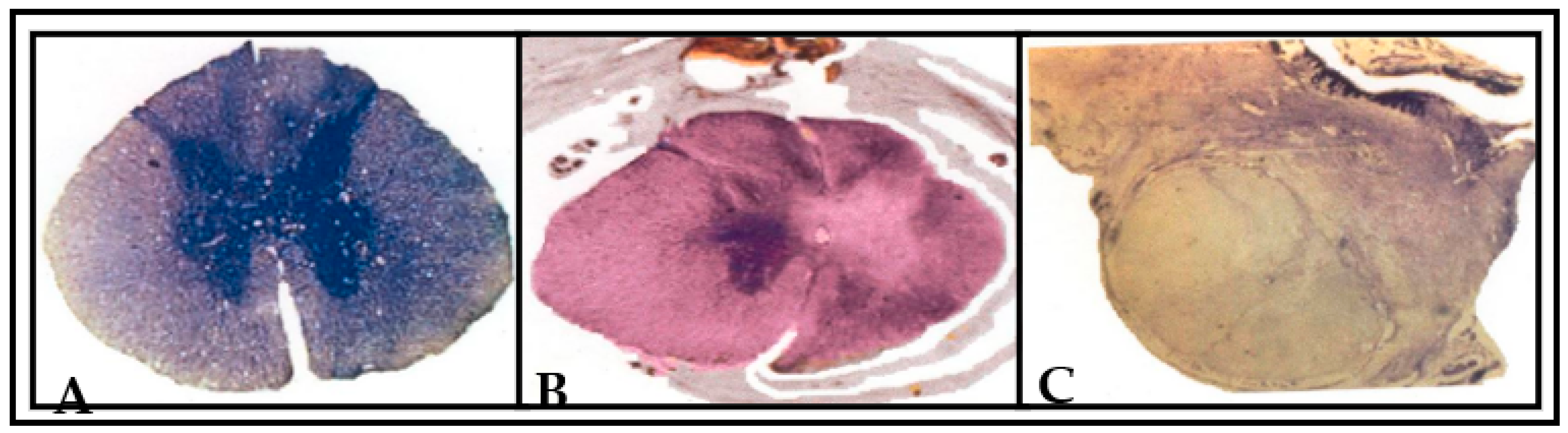
2.9. Histological Evaluation
2.10. Three (3-D) Dimensional Graphic Reconstruction
2.11. Statistical Analysis
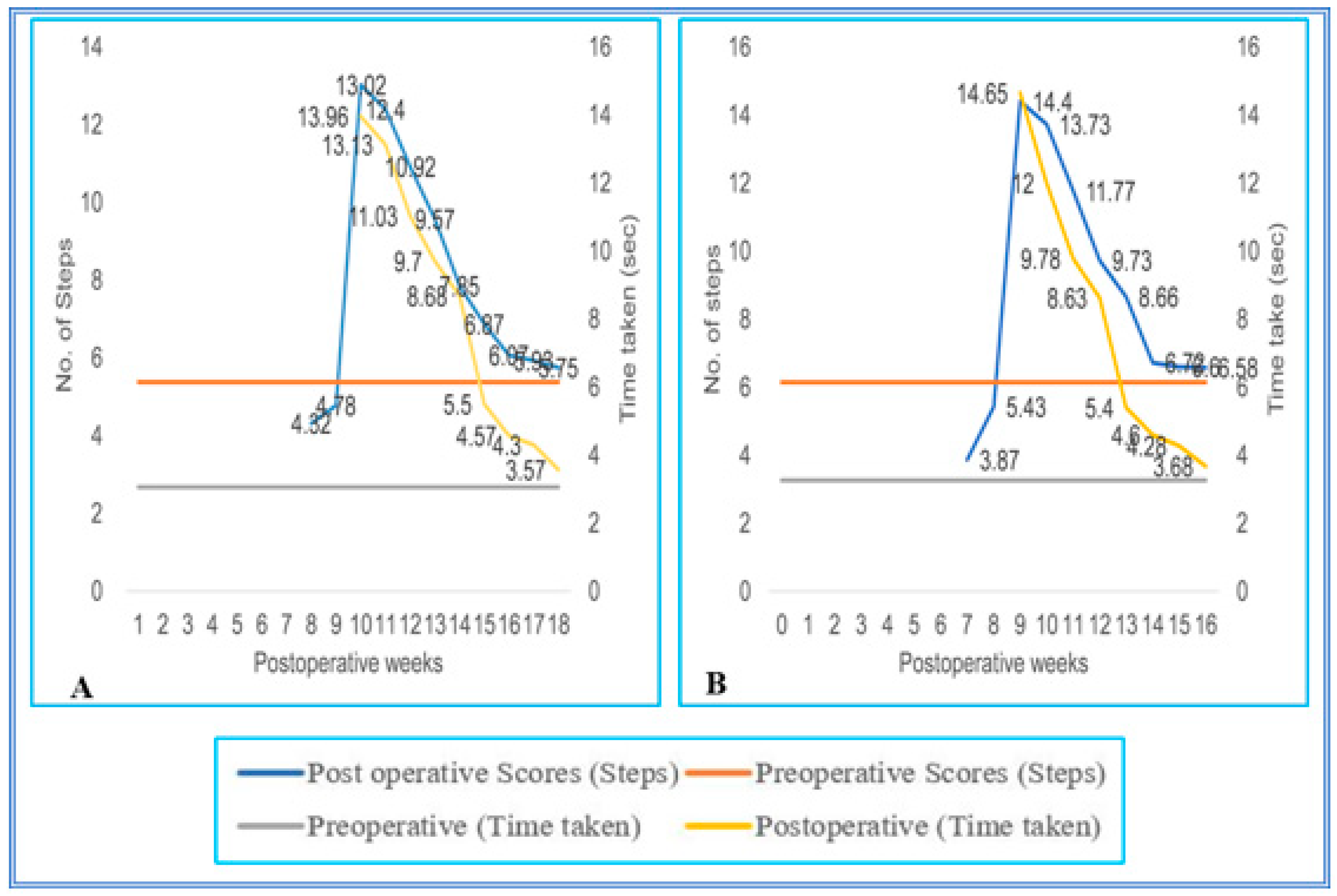
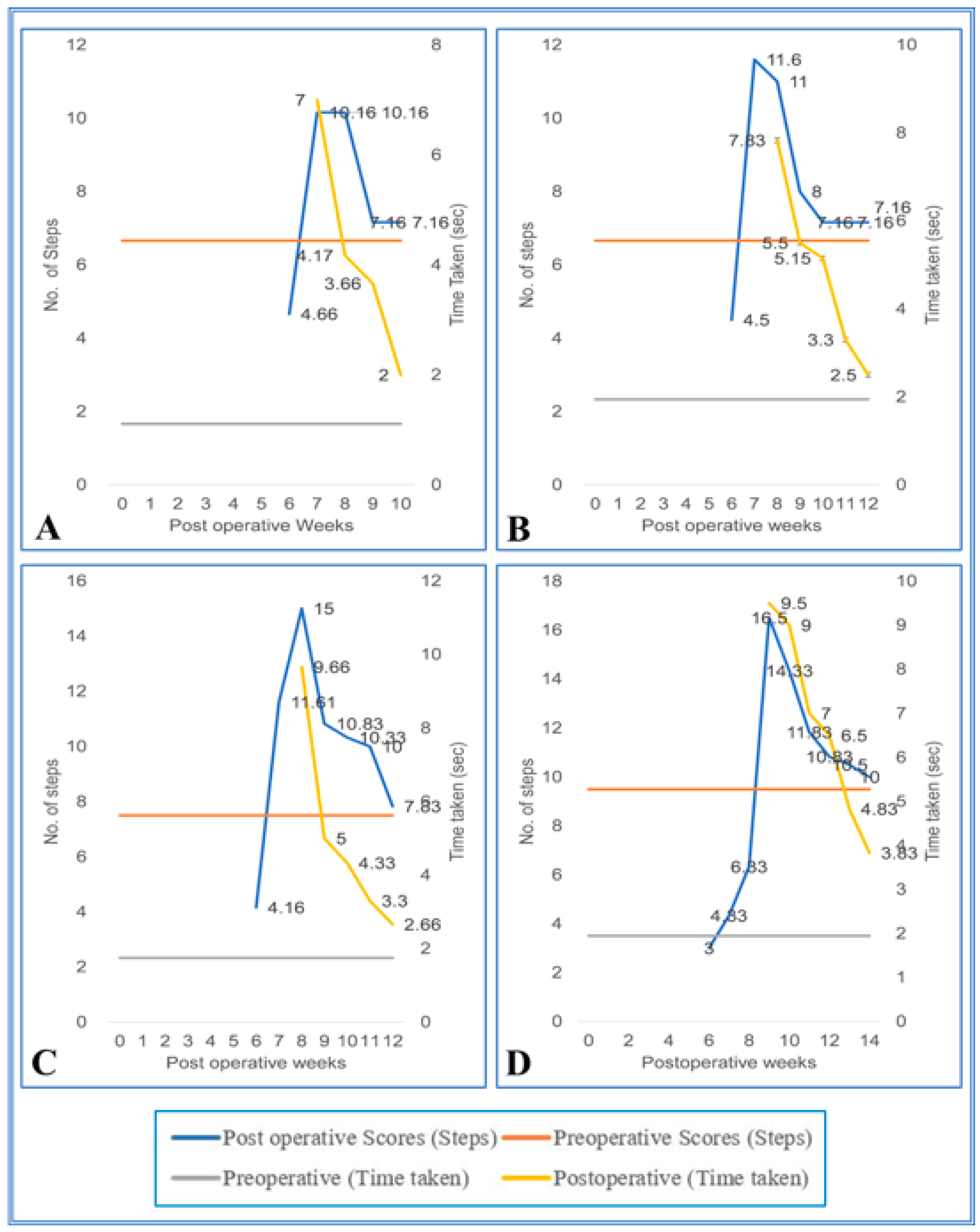
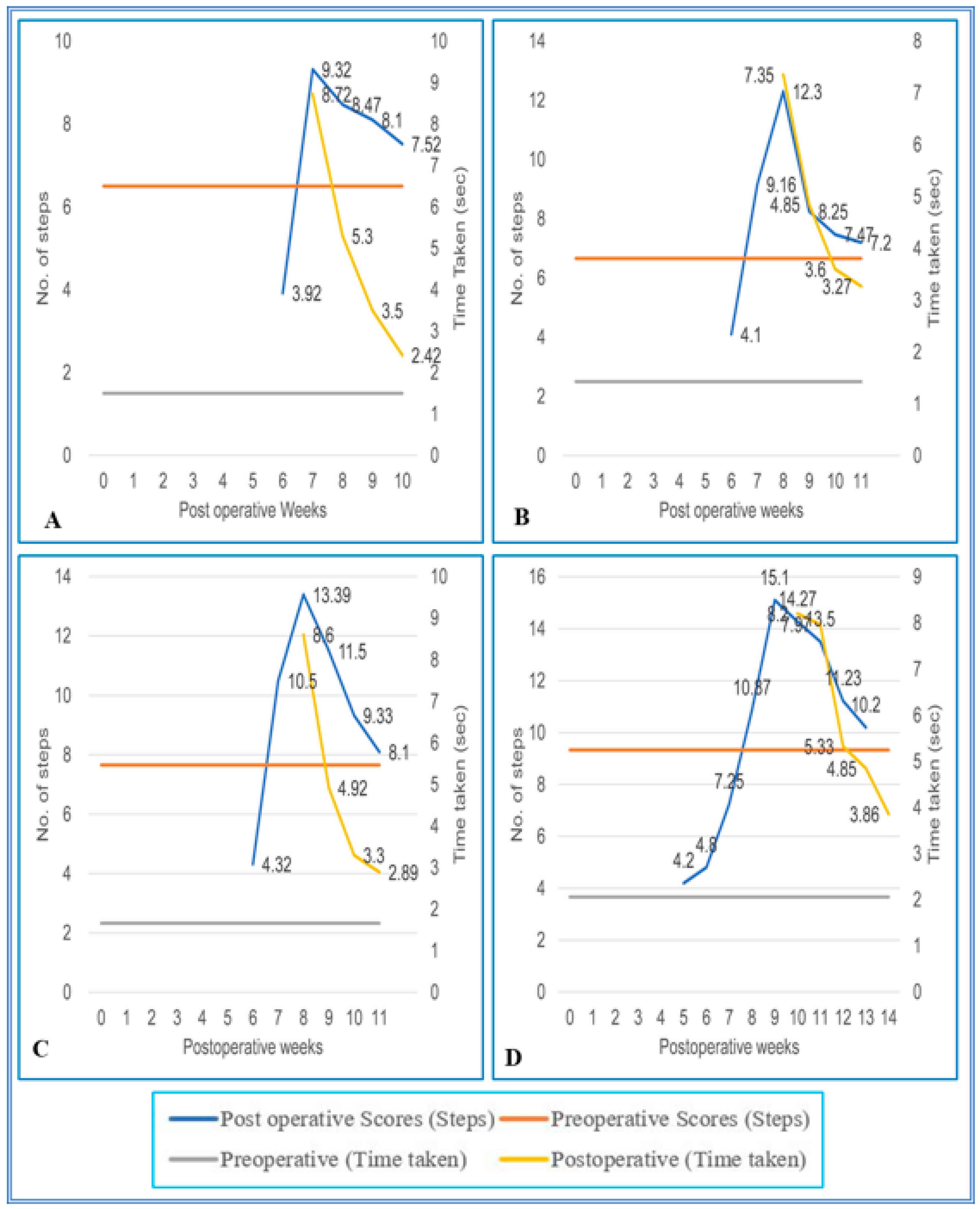
3. Results
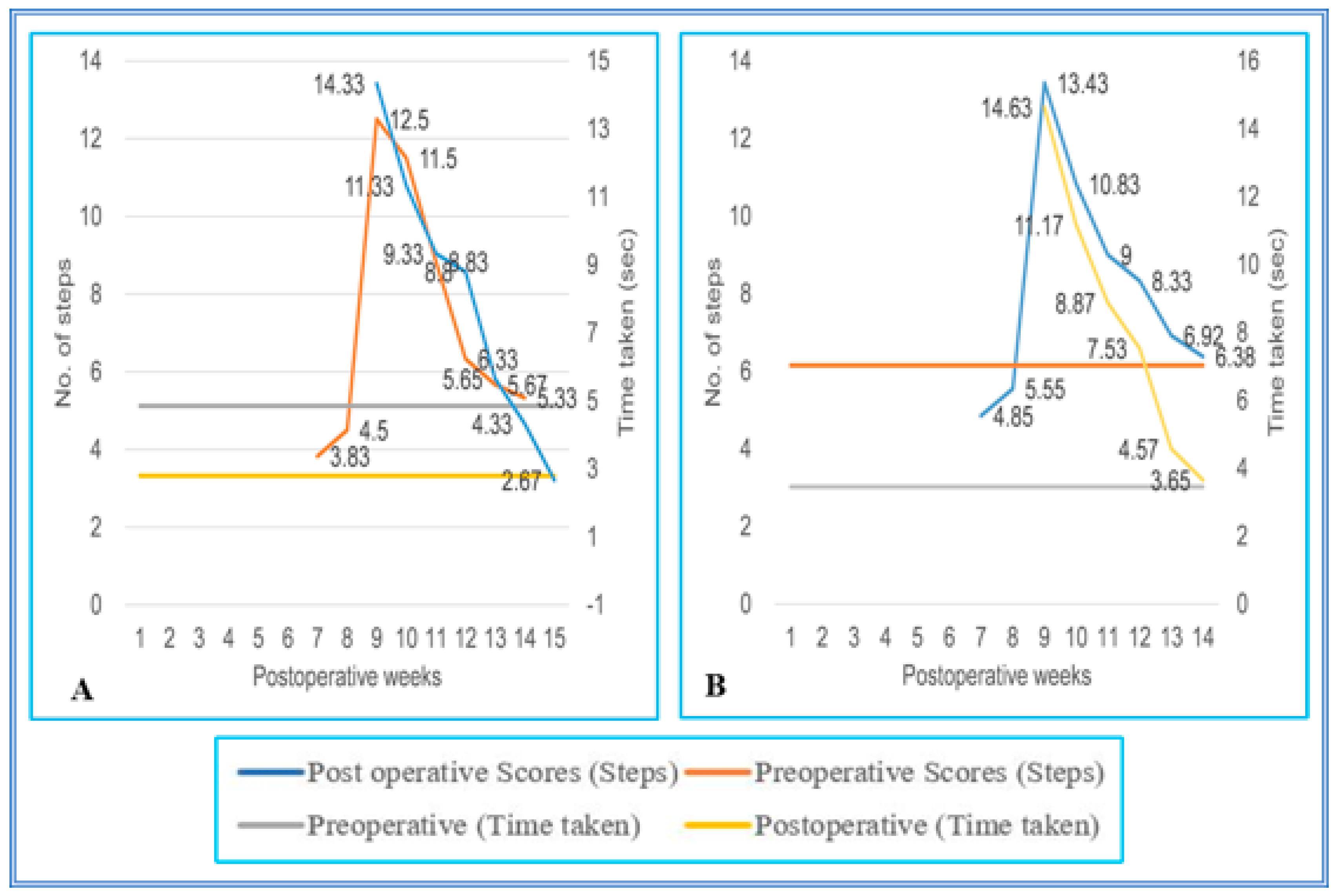
3.1. Staircase Runway
3.2. Grid Runways
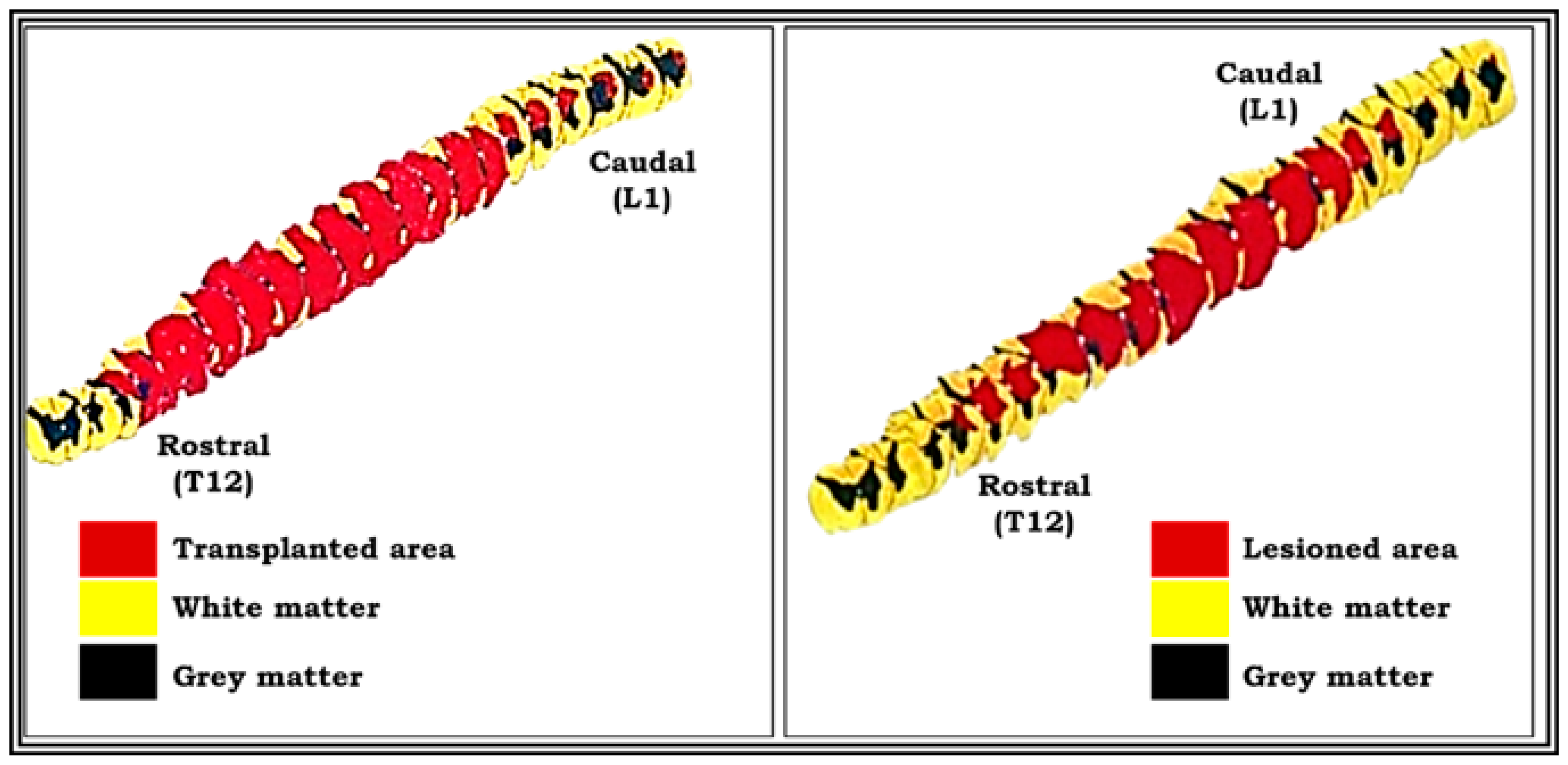
3.3. Histological and 3D Graphic Reconstruction Evaluation
4. Discussion
Strength and Limitations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reier, P.; Bregman, B.; Wujek, J. Intraspinal transplantation of embyronic spinal cord tissue in neonatal and adult rats. J. Comp. Neurol. 1986, 247, 275–296. [Google Scholar] [CrossRef]
- Lin, C.-C.; Lai, S.-R.; Shao, Y.-H.; Chen, C.-L.; Lee, K.-Z. The therapeutic effectiveness of delayed fetal spinal cord tissue transplantation on respiratory function following mid-cervical spinal cord injury. Neurotherapeutics 2017, 14, 792–809. [Google Scholar] [CrossRef] [Green Version]
- Côté, M.P.; Amin, A.A.; Tom, V.J.; Houle, J.D. Peripheral nerve grafts support regeneration after spinal cord injury. Neurotherapeutics 2011, 8, 294–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DePaul, M.A.; Lin, C.-Y.; Silver, J.; Lee, Y.-S. Peripheral nerve transplantation combined with acidic fibroblast growth factor and chondroitinase induces regeneration and improves urinary function in complete spinal cord transected adult mice. PLoS ONE 2015, 10, e0139335. [Google Scholar] [CrossRef]
- Kanno, H.; Pearse, D.D.; Ozawa, H.; Itoi, E.; Bunge, M.B. Schwann cell transplantation for spinal cord injury repair: Its significant therapeutic potential and prospectus. Rev. Neurosci. 2015, 26, 121–128. [Google Scholar] [CrossRef] [PubMed]
- De la Garza-Castro, O.; Martínez-Rodríguez, H.G.; Sánchez-González, S.G.; Vidal-Torres, O.; Arreola-Romero, A.; de la Garza-Pineda, O.; Ancer-Arellano, A.G.; Guzmán-López, S.; Elizondo-Omaña, R.E. Schwann cell precursor transplant in a rat spinal cord injury model. Rev. Investig. Clín. 2018, 70, 88–95. [Google Scholar] [CrossRef]
- Levi, A.D.; Dancausse, H.; Li, X.; Duncan, S.; Horkey, L.; Oliviera, M. Peripheral nerve grafts promoting central nervous system regeneration after spinal cord injury in the primate. J. Neurosurg. Spine 2002, 96, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Wang, Y.-L.; Kong, D.; Qu, B.; Su, X.-J.; Li, H.; Pi, H.-Y. Nerve autografts and tissue-engineered materials for the repair of peripheral nerve injuries: A 5-year bibliometric analysis. Neural Regen. Res. 2015, 10, 1003. [Google Scholar]
- Villegas-Perez, M.P.; Vidal-Sanz, M.; Bray, G.M.; Aguayo, A.J. Influences of peripheral nerve grafts on the survival and regrowth of axotomized retinal ganglion cells in adult rats. J. Neurosci. 1988, 8, 265–280. [Google Scholar] [CrossRef] [Green Version]
- Quan, M.-Z.; Kosaka, J.; Watanabe, M.; Wakabayashi, T.; Fukuda, Y. Survival of Axotomized Retinal Ganglion Cells in Peripheral Nerve–Grafted Ferrets. Investig. Ophthalmol. Vis. Sci. 1999, 40, 2360–2366. [Google Scholar]
- Richardson, P.; Issa, V.; Aguayo, A. Regeneration of long spinal axons in the rat. J. Neurocytol. 1984, 13, 165–182. [Google Scholar] [CrossRef]
- El Seblani, N.; Welleford, A.S.; Quintero, J.E.; van Horne, C.G.; Gerhardt, G.A. Invited review: Utilizing peripheral nerve regenerative elements to repair damage in the CNS. J. Neurosci. Methods 2020, 335, 108623. [Google Scholar] [CrossRef] [PubMed]
- Ramón-Cueto, A.; Plant, G.W.; Avila, J.; Bunge, M.B. Long-distance axonal regeneration in the transected adult rat spinal cord is promoted by olfactory ensheathing glia transplants. J. Neurosci. 1998, 18, 3803–3815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kao, C.C.; Chang, L.W.; Bloodworth, J., Jr. Axonal regeneration across transected mammalian spinal cords: An electron microscopic study of delayed microsurgical nerve grafting. Exp. Neurol. 1977, 54, 591–615. [Google Scholar] [CrossRef]
- Oudega, M.; Varon, S.; Hagg, T. Regeneration of adult rat sensory axons into intraspinal nerve grafts: Promoting effects of conditioning lesion and graft predegeneration. Exp. Neurol. 1994, 129, 194–206. [Google Scholar] [CrossRef] [PubMed]
- Dam-Hieu, P.; Liu, S.; Choudhri, T.; Said, G.; Tadié, M. Regeneration of primary sensory axons into the adult rat spinal cord via a peripheral nerve graft bridging the lumbar dorsal roots to the dorsal column. J. Neurosci. Res. 2002, 68, 293–304. [Google Scholar] [CrossRef] [PubMed]
- Decherchi, P.; Gauthier, P. In vitro pre-degenerated nerve autografts support CNS axonal regeneration. Brain Res. 1996, 726, 181–188. [Google Scholar] [CrossRef]
- Gauthier, P.; Rega, P.; Lammari-Barreault, N.; Polentes, J. Functional reconnections established by central respiratory neurons regenerating axons into a nerve graft bridging the respiratory centers to the cervical spinal cord. J. Neurosci. Res. 2002, 70, 65–81. [Google Scholar] [CrossRef]
- Senoo, E.; Tamaki, N.; Fujimoto, E.; Ide, C. Effects of prelesioned peripheral nerve graft on nerve regeneration in the rat spinal cord. Neurosurgery 1998, 42, 1347–1356. [Google Scholar] [CrossRef]
- Nout, Y.S.; Rosenzweig, E.S.; Brock, J.H.; Strand, S.C.; Moseanko, R.; Hawbecker, S.; Zdunowski, S.; Nielson, J.L.; Roy, R.R.; Courtine, G.; et al. Animal models of neurologic disorders: A nonhuman primate model of spinal cord injury. Neurotherapeutics 2012, 9, 380–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nardone, R.; Florea, C.; Höller, Y.; Brigo, F.; Versace, V.; Lochner, P.; Golaszewski, S.; Trinka, E. Rodent, large animal and non-human primate models of spinal cord injury. Zoology 2017, 123, 101–114. [Google Scholar] [CrossRef]
- Kerasidis, H.; Wrathall, J.R.; Gale, K. Behavioral assessment of functional deficit in rats with contusive spinal cord injury. J. Neurosci. Methods 1987, 20, 167–179. [Google Scholar] [CrossRef]
- Wrathall, J.; Rigamonti, D.; Braford, M.; Kao, C. Reconstruction of the contused cat spinal cord by the delayed nerve graft technique and cultured peripheral non-neuronal cells. Acta Neuropathol. 1982, 57, 59–69. [Google Scholar] [CrossRef]
- Fernandez, E.; Pallini, R.; Maira, G.; Rossi, G. Peripheral nerve autografts to the injured spinal cord of the rat: An experimental model for the study of spinal cord regeneration. Acta Neurochir. 1985, 78, 57–64. [Google Scholar] [CrossRef]
- Suresh Babu, R.; Anand, P.; Jeraud, M.; Periasamy, P.; Namasivayam, A. Bipedal locomotion of bonnet macaques after spinal cord injury. Mot. Control 2007, 11, 322–347. [Google Scholar] [CrossRef]
- Kao, C.C.; Chang, L.W.; Bloodworth, J.M. The mechanism of spinal cord cavitation following spinal cord transection: Part 2: Electron microscopic observations. J. Neurosurg. 1977, 46, 745–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richardson, P.; McGuinness, U.; Aguayo, A. Peripheral nerve autografts to the rat spinal cord: Studies with axonal tracing methods. Brain Res. 1982, 237, 147–162. [Google Scholar] [CrossRef]
- Lewin-Kowalik, J.; Sieron, A.L.; Krause, M.; Barski, J.-J.; Górka, D. Time-dependent regenerative influence of predegenerated nerve grafts on hippocampus. Brain Res. Bull. 1992, 29, 831–835. [Google Scholar] [CrossRef]
- Zhao, Q.; Kerns, J.M. Effects of predegeneration on nerve regeneration through silicone Y-chambers. Brain Res. 1994, 633, 97–104. [Google Scholar] [CrossRef]
- Gordon, T. The role of neurotrophic factors in nerve regeneration. Neurosurg. Focus 2009, 26, E3. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Notterpek, L. Promoting peripheral myelin repair. Exp. Neurol. 2016, 283, 573–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danielsen, N.; Kerns, J.M.; Holmquist, B.; Zhao, Q.; Lundborg, G.; Kanje, M. Pre-degenerated nerve grafts enhance regeneration by shortening the initial delay period. Brain Res. 1994, 666, 250–254. [Google Scholar] [CrossRef]
- Danielsen, N.; Kerns, J.M.; Holmquist, B.; Zhao, Q.; Lundborg, G.; Kanje, M. Predegeneration enhances regeneration into acellular nerve grafts. Brain Res. 1995, 681, 105–108. [Google Scholar] [CrossRef]
- Ellis, J.C.; McCaffrey, T.V. Nerve grafting: Functional results after primary vs delayed repair. Arch. Otolaryngol. 1985, 111, 781–785. [Google Scholar] [CrossRef]
- Lewin-Kowalik, J.; Górka, D.; Larysz-Brysz, M.; Goka, B.; Swiech-Sabuda, E.; Małecka-Tendera, E.; Krause, M. Short-time predegenerated peripheral nerve grafts promote regrowth of injured hippocampal neurites. Acta Physiol. Hung. 1997, 85, 259–268. [Google Scholar]
- Ochi, M.; Wakasa, M.; Ikuta, Y.; Kwong, W. Nerve regeneration in predegenerated basal lamina graft: The effect of duration of predegeneration on axonal extension. Exp. Neurol. 1994, 128, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Hall, S. Axonal regeneration through acellular muscle grafts. J. Anat. 1997, 190, 57–71. [Google Scholar] [CrossRef] [PubMed]
- Paino, C.; Fernandez-Valle, C.; Bates, M.; Bunge, M. Regrowth of axons in lesioned adult rat spinal cord: Promotion by implants of cultured Schwann cells. J. Neurocytol. 1994, 23, 433–452. [Google Scholar] [CrossRef]
- Oudega, M.; Hagg, T. Nerve growth factor promotes regeneration of sensory axons into adult rat spinal cord. Exp. Neurol. 1996, 140, 218–229. [Google Scholar] [CrossRef]
- Richardson, P.; Issa, V.; Shemie, S. Regeneration and retrograde degeneration of axons in the rat optic nerve. J. Neurocytol. 1982, 11, 949–966. [Google Scholar] [CrossRef]
- Kobayashi, N.R.; Fan, D.-P.; Giehl, K.M.; Bedard, A.M.; Wiegand, S.J.; Tetzlaff, W. BDNF and NT-4/5 prevent atrophy of rat rubrospinal neurons after cervical axotomy, stimulate GAP-43 and Tα1-tubulin mRNA expression, and promote axonal regeneration. J. Neurosci. 1997, 17, 9583–9595. [Google Scholar] [CrossRef] [Green Version]
- Kwon, B.K.; Liu, J.; Oschipok, L.; Teh, J.; Liu, Z.W.; Tetzlaff, W. Rubrospinal neurons fail to respond to brain-derived neurotrophic factor applied to the spinal cord injury site 2 months after cervical axotomy. Exp. Neurol. 2004, 189, 45–57. [Google Scholar] [CrossRef]
- Tetzlaff, W.; Alexander, S.W.; Miller, F.D.; Bisby, M.A. Response of facial and rubrospinal neurons to axotomy: Changes in mRNA expression for cytoskeletal proteins and GAP-43. J. Neurosci. 1991, 11, 2528–2544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, K.J.; Fan, D.P.; Tsui, B.; Cassar, S.; Tetzlaff, W. Influence of the axotomy to cell body distance in rat rubrospinal and spinal motoneurons: Differential regulation of GAP-43, tubulins, and neurofilament-M. J. Comp. Neurol. 1999, 414, 495–510. [Google Scholar] [CrossRef]
- Jenkins, R.; Tetzlaff, W.; Hunt, S. Differential expression of immediate early genes in rubrospinal neurons following axotomy in rat. Eur. J. Neurosci. 1993, 5, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Broude, E.; McAtee, M.; Kelley, M.; Bregman, B. c-Jun expression in adult rat dorsal root ganglion neurons: Differential response after central or peripheral axotomy. Exp. Neurol. 1997, 148, 367–377. [Google Scholar] [CrossRef] [Green Version]
- Weidner, N.; Blesch, A.; Grill, R.; Tuszynski, M. Nerve growth factor–hypersecreting Schwann cell grafts augment and guide spinal cord axonal growth and remyelinate central nervous system axons in a phenotypically appropriate manner that correlates with expression of L1. J. Comp. Neurol. 1999, 413, 495–506. [Google Scholar] [CrossRef]
- Liuzzi, F.J.; Tedeschi, B. Axo-glial interactions at the dorsal root transitional zone regulate neurofilament protein synthesis in axotomized sensory neurons. J. Neurosci. 1992, 12, 4783–4792. [Google Scholar] [CrossRef] [Green Version]
- Fraher, J.P. The transitional zone and CNS regeneration. J. Anat. 2000, 196, 137–158. [Google Scholar] [CrossRef] [PubMed]
- Pindzola, R.; Doller, C.; Silver, J. Putative inhibitory extracellular matrix molecules at the dorsal root entry zone of the spinal cord during development and after root and sciatic nerve lesions. Dev. Biol. 1993, 156, 34–48. [Google Scholar] [CrossRef] [PubMed]
- Roitbak, T.; Syková, E. Diffusion barriers evoked in the rat cortex by reactive astrogliosis. Glia 1999, 28, 40–48. [Google Scholar] [CrossRef]
- Stichel, C.C.; Hermanns, S.; Luhmann, H.J.; Lausberg, F.; Niermann, H.; D’Urso, D.; Servos, G.; Hartwig, H.G.; Müller, H.W. Inhibition of collagen IV deposition promotes regeneration of injured CNS axons. Eur. J. Neurosci. 1999, 11, 632–646. [Google Scholar] [CrossRef] [PubMed]
- Keirstead, H.S.; Dyer, J.K.; Sholomenko, G.N.; McGraw, J.; Delaney, K.; Steeves, J. Axonal regeneration and physiological activity following transection and immunological disruption of myelin within the hatchling chick spinal cord. J. Neurosci. 1995, 15, 6963–6974. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.S.; Huber, A.B.; van der Haar, M.E.; Frank, M.; Schnell, L.; Spillmann, A.A.; Christ, F.; Schwab, M.E. Nogo-A is a myelin-associated neurite outgrowth inhibitor and an antigen for monoclonal antibody IN-1. Nature 2000, 403, 434–439. [Google Scholar] [CrossRef] [PubMed]
- Oertle, T.; Van Der Haar, M.E.; Bandtlow, C.E.; Robeva, A.; Burfeind, P.; Buss, A.; Huber, A.B.; Simonen, M.; Schnell, L.; Brösamle, C.; et al. Nogo-A inhibits neurite outgrowth and cell spreading with three discrete regions. J. Neurosci. 2003, 23, 5393–5406. [Google Scholar] [CrossRef] [PubMed]
- Fitch, M.T.; Silver, J. Glial cell extracellular matrix: Boundaries for axon growth in development and regeneration. Cell Tissue Res. 1997, 290, 379–384. [Google Scholar] [CrossRef] [PubMed]
- Fawcett, J.W.; Asher, R.A. The glial scar and central nervous system repair. Brain Res. Bull. 1999, 49, 377–391. [Google Scholar] [CrossRef]
- Tang, X.; Davies, J.E.; Davies, S.J. Changes in distribution, cell associations, and protein expression levels of NG2, neurocan, phosphacan, brevican, versican V2, and tenascin-C during acute to chronic maturation of spinal cord scar tissue. J. Neurosci. Res. 2003, 71, 427–444. [Google Scholar] [CrossRef] [PubMed]
- Buss, A.; Pech, K.; Kakulas, B.A.; Martin, D.; Schoenen, J.; Noth, J.; Brook, G.A. NG2 and phosphacan are present in the astroglial scar after human traumatic spinal cord injury. BMC Neurol. 2009, 9, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasterkamp, R.J.; Giger, R.J.; Ruitenberg, M.J.; Holtmaat, A.; De Wit, J.; De Winter, F.; Verhaagen, J. Expression of the gene encoding the chemorepellent semaphorin III is induced in the fibroblast component of neural scar tissue formed following injuries of adult but not neonatal CNS. Mol. Cell. Neurosci. 1999, 13, 143–166. [Google Scholar] [CrossRef]
- De Winter, F.; Oudega, M.; Lankhorst, A.; Hamers, F.; Blits, B.; Ruitenberg, M.J.; Pasterkamp, R.J.; Gispen, W.H.; Verhaagen, J. Injury-induced class 3 semaphorin expression in the rat spinal cord. Exp. Neurol. 2002, 175, 61–75. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, M.; Ino, H.; Koda, M.; Murakami, M.; Yoshinaga, K.; Yamazaki, M.; Moriya, H. Regulation of semaphorin 3A expression in neurons of the rat spinal cord and cerebral cortex after transection injury. Acta Neuropathol. 2004, 107, 250–256. [Google Scholar] [CrossRef]
- Haas, C.A.; Rauch, U.; Thon, N.; Merten, T.; Deller, T. Entorhinal cortex lesion in adult rats induces the expression of the neuronal chondroitin sulfate proteoglycan neurocan in reactive astrocytes. J. Neurosci. 1999, 19, 9953–9963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemons, M.L.; Howland, D.R.; Anderson, D.K. Chondroitin sulfate proteoglycan immunoreactivity increases following spinal cord injury and transplantation. Exp. Neurol. 1999, 160, 51–65. [Google Scholar] [CrossRef] [PubMed]
- Ikegami, T.; Nakamura, M.; Yamane, J.; Katoh, H.; Okada, S.; Iwanami, A.; Watanabe, K.; Ishii, K.; Kato, F.; Fujita, H. Chondroitinase ABC combined with neural stem/progenitor cell transplantation enhances graft cell migration and outgrowth of growth-associated protein-43-positive fibers after rat spinal cord injury. Eur. J. Neurosci. 2005, 22, 3036–3046. [Google Scholar] [CrossRef] [PubMed]
- Iseda, T.; Okuda, T.; Kane-Goldsmith, N.; Mathew, M.; Ahmed, S.; Chang, Y.-W.; Young, W.; Grumet, M. Single, high-dose intraspinal injection of chondroitinase reduces glycosaminoglycans in injured spinal cord and promotes corticospinal axonal regrowth after hemisection but not contusion. J. Neurotrauma 2008, 25, 334–349. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Winsor, K.; Nienhaus, C.; Hess, E.; Blackmore, M.G. Combined chondroitinase and KLF7 expression reduce net retraction of sensory and CST axons from sites of spinal injury. Neurobiol. Dis. 2017, 99, 24–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schreyer, D.; Jones, E. Growth of corticospinal axons on prosthetic substrates introduced into the spinal cord of neonatal rats. Dev. Brain Res. 1987, 35, 291–299. [Google Scholar] [CrossRef]
- Schnell, L.; Schwab, M. Sprouting and regeneration of lesioned corticospinal tract fibres in the adult rat spinal cord. Eur. J. Neurosci. 1993, 5, 1156–1171. [Google Scholar] [CrossRef] [PubMed]
- Khan, T.; Dauzvardis, M.; Sayers, S. Carbon filament implants promote axonal growth across the transected rat spinal cord. Brain Res. 1991, 541, 139–145. [Google Scholar] [CrossRef]
- Xu, X.M.; Chen, A.; Guenard, V.; Kleitman, N.; Bunge, M.B. Bridging Schwann cell transplants promote axonal regeneration from both the rostral and caudal stumps of transected adult rat spinal cord. J. Neurocytol. 1997, 26, 1–16. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Score | Behavior in Staircase Runway | Behavior in Grid Runway |
|---|---|---|
| 0 grade/10 points | No attempt to stand, walk, no weight bearing | No attempt to stand, walk, no weight bearing |
| 1 grade/8 points | Weak and/or delayed attempt to stand, no attempt to walk on the runway (initiated one or two steps) | Weak and/or delayed attempt to stand, no attempt to walk on the runway (initiated one or two steps) |
| 2 grade/6 points | Good attempt to support body weight, weak attempt to walk on the runway with frequent slipping/errors seen | Good attempt to support body weight, weak attempt to walk on the runway with frequent errors seen |
| 3 grade/4 points | Good attempt to stand and walk on the runway with few errors, significant change in time to cross the runway | Good attempt to stand and walk on the runway with few errors, significant change in time to cross the runway |
| 4 grade/2 points | Good attempt to stand and walk on the runway with only mild deficits, no slipping, no significant change in time taken to cross the runway | Good attempt to stand and walk on the runway with only mild deficits, no misplacement, no significant change in time taken to cross the runway |
| 5 grade/0 points | Good attempt to walk on the runway, no significant change in time compared to control animals | Good attempt to walk on the runway, no significant change in time compared to control animals |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paramasivam, A.; Mickymaray, S.; Jayakumar, S.; Jeraud, M.; Perumal, P.; Alassaf, A.; Aljabr, A.A.; Dasarathy, S.; Rangasamy, S.B. Locomotor Behavior Analysis in Spinal Cord Injured Macaca radiata after Predegenerated Peripheral Nerve Grafting—A Preliminary Evidence. Vet. Sci. 2021, 8, 288. https://doi.org/10.3390/vetsci8120288
Paramasivam A, Mickymaray S, Jayakumar S, Jeraud M, Perumal P, Alassaf A, Aljabr AA, Dasarathy S, Rangasamy SB. Locomotor Behavior Analysis in Spinal Cord Injured Macaca radiata after Predegenerated Peripheral Nerve Grafting—A Preliminary Evidence. Veterinary Sciences. 2021; 8(12):288. https://doi.org/10.3390/vetsci8120288
Chicago/Turabian StyleParamasivam, Anand, Suresh Mickymaray, Saikarthik Jayakumar, Mathew Jeraud, Periasamy Perumal, Abdullah Alassaf, Abdullah Abdulrahman Aljabr, Sridevi Dasarathy, and Suresh Babu Rangasamy. 2021. "Locomotor Behavior Analysis in Spinal Cord Injured Macaca radiata after Predegenerated Peripheral Nerve Grafting—A Preliminary Evidence" Veterinary Sciences 8, no. 12: 288. https://doi.org/10.3390/vetsci8120288
APA StyleParamasivam, A., Mickymaray, S., Jayakumar, S., Jeraud, M., Perumal, P., Alassaf, A., Aljabr, A. A., Dasarathy, S., & Rangasamy, S. B. (2021). Locomotor Behavior Analysis in Spinal Cord Injured Macaca radiata after Predegenerated Peripheral Nerve Grafting—A Preliminary Evidence. Veterinary Sciences, 8(12), 288. https://doi.org/10.3390/vetsci8120288