
The Infectious Bronchitis Coronavirus Pneumonia Model Presenting a Novel Insight for the SARS-CoV-2 Dissemination Route

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Formulations
2.2. Bacteriophage φ6 In Vitro Model
2.3. Poultry
2.4. Inoculum Administration
2.5. Dosage Administration
2.6. Histopathological Evaluation
2.7. qRT-PCR
3. Results
3.1. Verification of the Virucidal Action with Bacteriophage φ6
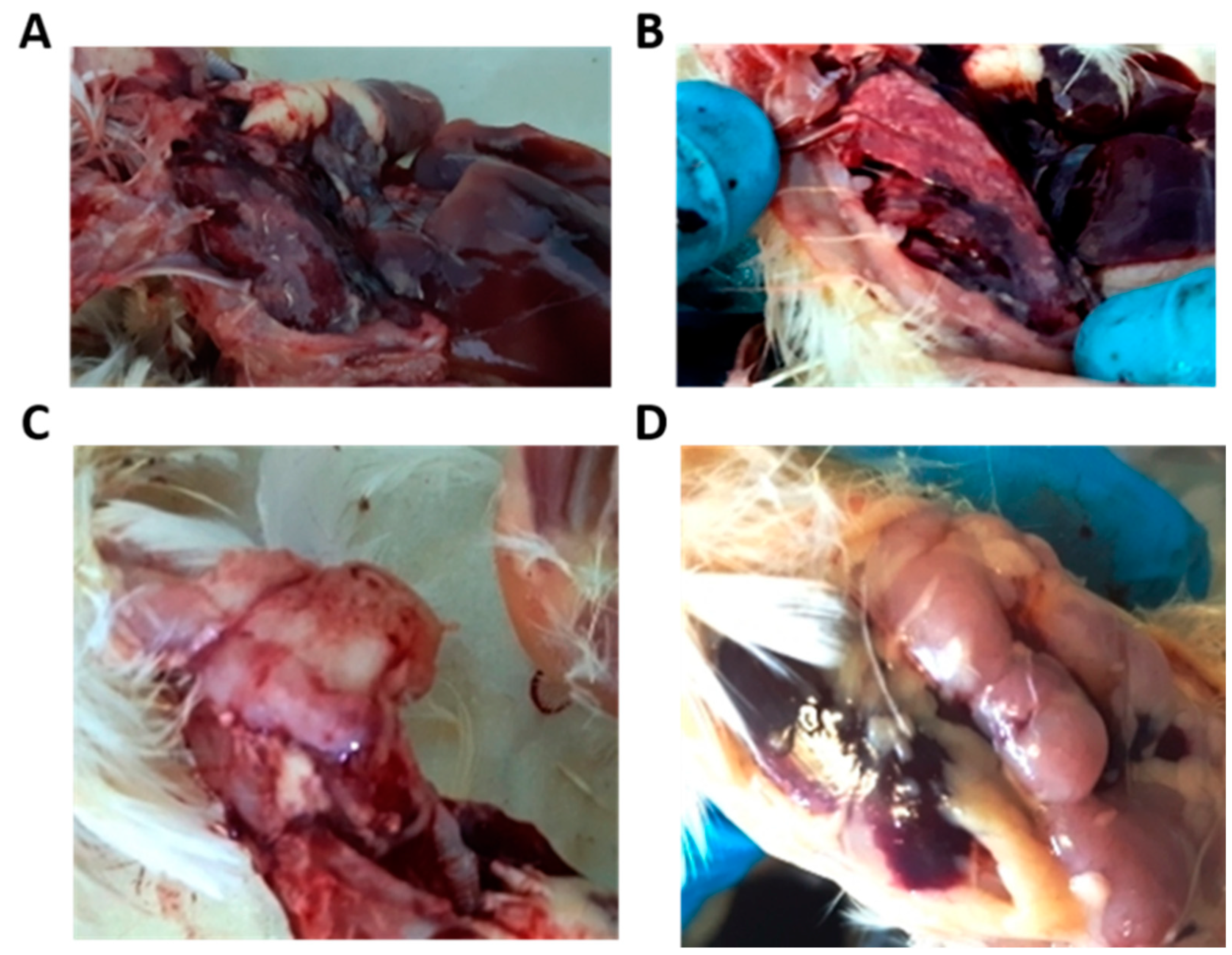
3.2. Postmortem Examination
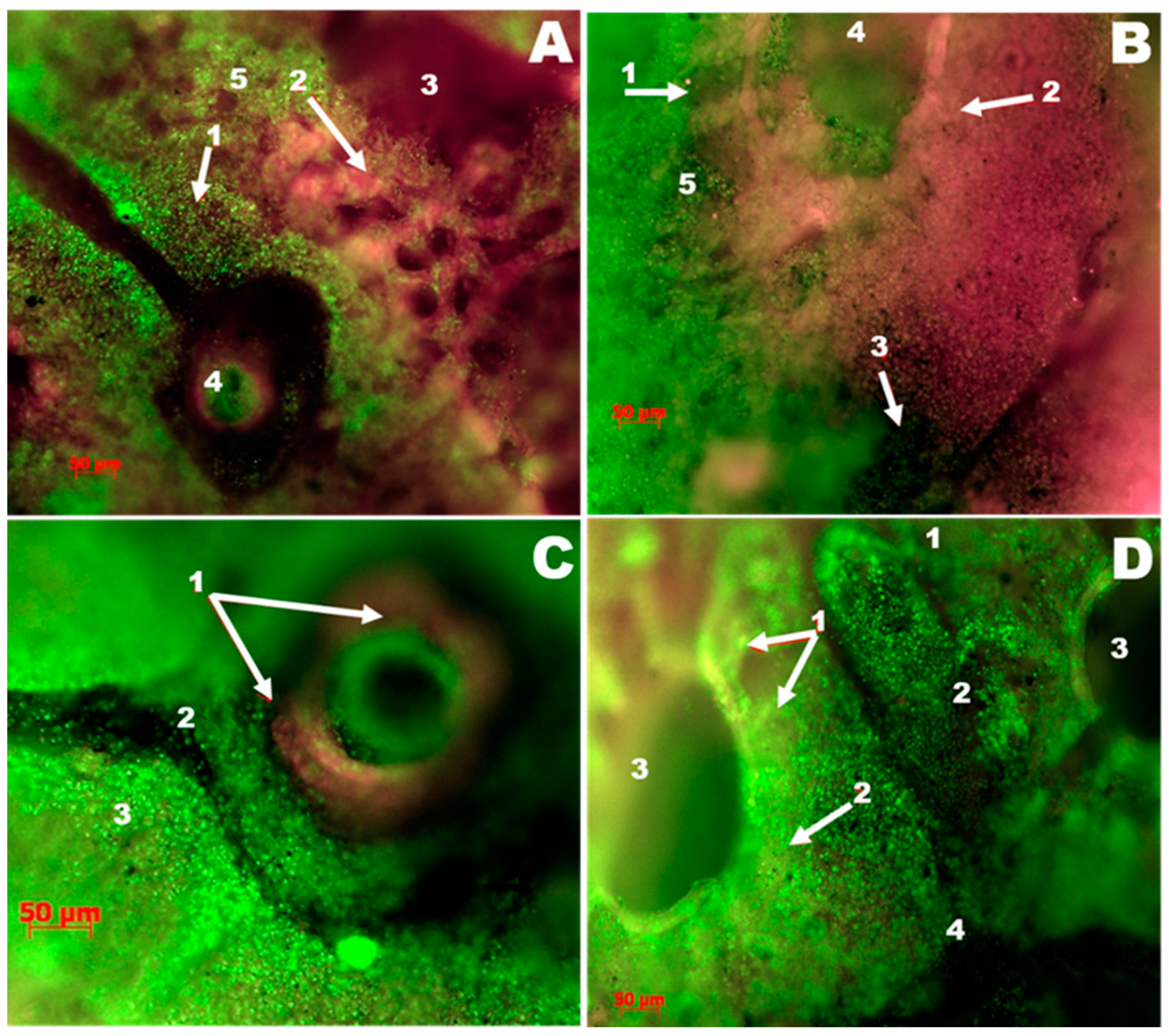
3.3. Lung Luminescence Microscopy Histopathological Study
3.3.1. The Control Group
3.3.2. The Group Treated with Argovit
3.3.3. IBV in Chickens with RT-PCR
Intestine
Lungs
Thymus
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Afonyushkin, V.N.; Mironova, V.S.C. Possible pathogenesis of coronavirus infections on the example of IBV as a model of infection associated with COVID-19 in humans. BIO. J. Spec. Poult. Livest. Farms 2020, 4, 4–6. [Google Scholar]
- Afonyushkin, V.N.; Litvinov, A.V. An alternative way to prevent and treat PRRS. Pig-Breeding 2017, 56–58. Available online: https://www.svinoprom.ru/ (accessed on 11 October 2021).
- Cavanagh, D. Coronaviruses in poultry and other birds. Avian Pathol. 2005, 34, 439–448. [Google Scholar] [CrossRef] [PubMed]
- Cowen, B.S.; Wideman, R.F.; Braune, M.O.; Owen, R.L. An infectious bronchitis virus isolated from chickens experiencing a urolithiasis outbreak. I. In vitro characterization studies. Avian Dis. 1987, 31, 878–883. [Google Scholar] [CrossRef]
- Su, S.; Wong, G.; Shi, W.; Liu, J.; Lai, A.C.; Zhou, J.; Liu, W.; Bi, Y.; Gao, G.F. Epidemiology, genetic recombination, and pathogenesis of coronaviruses. Trends Microbiol. 2016, 24, 490–502. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Balagna, C.; Perero, S.; Percivalle, E.; Nepita, E.V.; Ferraris, M. Virucidal effect against coronavirus SARS-CoV-2 of a silver nanocluster/silica composite sputtered coating. Open Ceram. 2020, 1, 100006. [Google Scholar] [CrossRef]
- Elechiguerra, J.L.; Burt, J.L.; Morones, J.R.; Camacho-Bragado, A.; Gao, X.; Lara, H.H.; Yacaman, M.J. Interaction of silver nanoparticles with HIV-1. J. Nanobiotechnol. 2005, 3, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawase, M.; Shirato, K.; van der Hoek, L.; Taguchi, F.; Matsuyama, S. Simultaneous Treatment of Human Bronchial Epithelial Cells with Serine and Cysteine Protease Inhibitors Prevents Severe Acute Respiratory Syndrome Coronavirus Entry. J. Virol. 2012, 86, 6537–6545. [Google Scholar] [CrossRef] [Green Version]
- Fedorenko, A.; Grinberg, M.; Orevi, T.; Kashtan, N. Survival of the enveloped bacteriophage Phi6 (a surrogate for SARS-CoV-2) in evaporated saliva microdroplets deposited on glass surfaces. Sci. Rep. 2020, 10, 22419. [Google Scholar] [CrossRef] [PubMed]
- Kakutani, K.; Matsuda, Y.; Nonomura, T.; Takikawa, Y.; Takami, T.; Toyoda, H. A Simple Electrostatic Precipitator for Trapping Virus Particles Spread via Droplet Transmission. Int. J. Environ. Res. Public Health 2021, 18, 4934. [Google Scholar] [CrossRef] [PubMed]
- Afonyushkin, V.N.; Cherepushkina, V.S.; Tatarchuk, O.P.; Frolova, O.A. Study of anti-phage activity of disinfectants as a factor of suppressing horizontal gene transfer. Bull. KSAU 2020, 4, 88–96. [Google Scholar]
- Afonyushkin, V.N.; Tabanyukhov, K.A.; Cherepushkina, V.S.; Khomenko, Y.S.; Tatarchuk, O.P. Effect of disinfectants based on potassium persulfate, hydrogen peroxide, glutaraldehyde and quaternary ammonium compounds on the genetic material of the pathogen bacteria specific to meat processing industry. Theory Pract. Meat Process. 2016, 1, 54–61. [Google Scholar] [CrossRef]
- Afonyushkin, V.N.; Shirshova, A.N.; Shamovskaya, D.V.; Plomodyalov, D.N. A study of the antiviral effect of drug triviron on IBV. Veterinary 2018, 24–28. Available online: https://agris.fao.org/agris-search/search.do?recordID=RU2019000107 (accessed on 11 October 2021).
- Burakova, E.; Kovalev, N.; Zenkova, M.; Vlassov, V.; Silnikov, V. Structure–activity relationships in new polycationic molecules based on two 1,4-diazabicyclo[2.2.2]octanes as artificial ribonucleases. Bioorg. Chem. 2014, 57, 127–131. [Google Scholar] [CrossRef]
- Afonyushkin, V.N. Influence of flaviviral infection on broiler productivity. Veterinary 2014, 8, 15–19. [Google Scholar]
- Baram-Pinto, D.; Shukla, S.; Perkas, N.; Gedanken, A.; Sarid, R. Inhibition of Herpes Simplex Virus Type 1 Infection by Silver Nanoparticles Capped with Mercaptoethane Sulfonate. Bioconjug. Chem. 2009, 20, 1497–1502. [Google Scholar] [CrossRef]
- Lara, H.H.; Ayala-Nuñez, N.V.; Ixtepan-Turrent, L.; Rodriguez-Padilla, C. Mode of antiviral action of silver nanoparticles against HIV-1. J. Nanobiotechnol. 2010, 8, 1. [Google Scholar] [CrossRef]
- Lara, H.H.; Ixtepan-Turrent, L.; Garza-Treviño, E.N.; Rodriguez-Padilla, C. PVP-coated silver nanoparticles block the transmission of cell-free and cell-associated HIV-1 in human cervical culture. J. Nanobiotechnol. 2010, 8, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Singh, A.K.; Vig, K.; Pillai, S.R.; Singh, S.R. Silver nanoparticles inhibit replication of respiratory syncytial virus. J. Biomed. Nanotechnol. 2008, 4, 149–158. [Google Scholar]
- Romo-Quiñonez, C.R.; Álvarez-Sánchez, A.R.; Álvarez-Ruiz, P.; Chávez-Sánchez, M.C.; Bogdanchikova, N.; Pestryakov, A.; Mejia-Ruiz, C.H. Evaluation of a new Argovit as an antiviral agent included in feed to protect the shrimp Litopenaeus vannamei against White Spot Syndrome Virus infection. PeerJ 2020, 8, e8446. [Google Scholar] [CrossRef] [Green Version]
- Borrego, B.; Lorenzo, G.; Mota-Morales, J.D.; Almanza-Reyes, H.; Mateos, F.; López-Gil, E.; de la Losa, N.; Burmistrov, V.A.; Pestryakov, A.N.; Brun, A.; et al. Potential application of silver nanoparticles to control the infectivity of Rift valley fever virus in vitro and in vivo. Nanomed. Nanotechnol. Biol. Med. 2016, 12, 1185–1192. [Google Scholar] [CrossRef] [PubMed]
- Bogdanchikova, N.; Vázquez-Muñoz, R.; Huerta-Saquero, A.; Pena-Jasso, A.; Aguilar-Uzcanga, G.; Picos-Díaz, P.L.; Pestryakov, A.; Burmistrov, V.; Martynyuk, O.; Luna-Vazquez-Gomez, R.; et al. Silver nanoparticles composition for treatment of distemper in dogs. Int. J. Nanotechnol. 2016, 13, 225–235. Available online: https://www.researchgate.net/publication/315664816_Silver_nanoparticles_composition_for_treatment_of_distemper_in_dogs (accessed on 8 September 2021). [CrossRef]
- Glotov, A.G.; Glotova, T.I.; Sergeev, A.A.; Sergeev, A.N. Study of Antiviral Activity of Different Drugs against Bovine Herpes Virus and Pestivirus. Antibiot. Chemother. 2004, 49, 6–9. Available online: https://www.researchgate.net/publication/8101788_Study_of_antiviral_activity_of_different_drugs_against_bovine_herpes_virus_and_pestivirus (accessed on 9 August 2021).
- Isaacs, C.E.; Kims, K.; Thormar, H. Inactivation of Enveloped Viruses in Human Bodily Fluids by Purified Lipids. Ann. N. Y. Acad. Sci. 1994, 724, 457–464. [Google Scholar] [CrossRef]
- Thormar, H.; Isaacs, C.E.; Brown, H.R.; Barshatzky, M.R.; Pessolano, T. Inactivation of enveloped viruses and killing of cells by fatty acids and monoglycerides. Antimicrob. Agents Chemother. 1987, 31, 27–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arora, R.; Chawla, R.; Marwah, R.; Arora, P.; Sharma, R.K.; Kaushik, V.; Goel, R.; Kaur, A.; Silambarasan, M.; Tripathi, R.P.; et al. Potential of Complementary and Alternative Medicine in Preventive Management of Novel H1N1 Flu (Swine Flu) Pandemic: Thwarting Potential Disasters in the Bud. Evid.-Based Complement. Altern. Med. 2011, 2011, 586506. [Google Scholar] [CrossRef]
- Sands, J.; Auperin, D.; Snipes, W. Extreme Sensitivity of Enveloped Viruses, Including Herpes Simplex, to Long-Chain Unsaturated Monoglycerides and Alcohols. Antimicrob. Agents Chemother. 1979, 15, 67–73. [Google Scholar] [CrossRef] [Green Version]
- Stephano-Hornedo, J.L.; Torres-Gutiérrez, O.; Toledano-Magaña, Y.; Gradilla-Martínez, I.; Pestryakov, A.; Sánchez-González, A.; García-Ramos, J.C.; Bogdanchikova, N. Argovit™ silver nanoparticles to fight Huanglongbing disease in Mexican limes (Citrus aurantifolia Swingle). RSC Adv. 2020, 10, 6146–6155. [Google Scholar] [CrossRef] [Green Version]
- Mironova, T.E.; Afonyushkin, V.N.; Cherepushkina, V.S.; Kozlova, Y.N.; Bobikova, A.S.; Koptev, V.Y.; Sigareva, N.A.; Kolpakov, F.A. Study of the protective effects of virucidal drugs on the model of coronavirus pneumonia. Vet. Korml. 2020, 35–38. [Google Scholar] [CrossRef]
- Joseph, T. International Pulmonologist’s Consensus on COVID-19, 2nd ed. 2020. Available online: https://teampata.org/portfolio/international-pulmonologists-consensus-on-covid-19/ (accessed on 11 October 2021).
- Qi, F.; Qian, S.; Zhang, S.; Zhang, Z. Single cell RNA sequencing of 13 human tissues identify cell types and receptors of human coronaviruses. Biochem. Biophys. Res. Commun. 2020, 526, 135–140. [Google Scholar] [CrossRef]
- The Human Protein Atlas Project Funded by the Knut & Alice Wallenberg Foundation. Available online: https://www.proteinatlas.org/ENSG00000130234-ACE2 (accessed on 1 October 2021).
- Lamers, M.M.; Beumer, J.; van der Vaart, J.; Knoops, K.; Puschhof, J.; Breugem, T.I.; Ravelli, R.B.G.; Paul van Schayck, J.; Mykytyn, A.Z.; Duimel, H.Q.; et al. SARS-CoV-2 productively infects human gut enterocytes. Science 2020, 369, 50–54. [Google Scholar] [CrossRef]
- Al-Beltagi, M.; Saeed, N.K.; Bediwy, A.S.; El-Sawaf, Y. Paediatric gastrointestinal disorders in SARS-CoV-2 infection: Epidemiological and clinical implications. World J. Gastroenterol. 2021, 27, 1716–1727. [Google Scholar] [CrossRef]
- Sfera, A.; Osorio, C.; Zapata Martín del Campo, C.M.; Pereida, S.; Maurer, S.; Maldonado, J.C.; Kozlakidis, Z. Endothelial Senescence and Chronic Fatigue Syndrome, a COVID-19 Based Hypothesis. Front. Cell. Neurosci. 2021, 15, 673217. [Google Scholar] [CrossRef] [PubMed]
- García-Pérez, B.E.; González-Rojas, J.A.; Salazar, M.I.; Torres-Torres, C.; Castrejón-Jiménez, N.S. Taming the Autophagy as a Strategy for Treating COVID-19. Cells 2020, 9, 2679. [Google Scholar] [CrossRef]
- Marino, G.; Niso-Santano, M.; Baehrecke, E.H.; Kroemer, G. Self-consumption: The interplay of autophagy and apoptosis. Nat. Rev. Mol. Cell Biol. 2014, 15, 81–94. [Google Scholar] [CrossRef] [Green Version]
- Young, M.M.; Takahashi, Y.; Khan, O.; Park, S.; Hori, T.; Yun, J.; Sharma, A.K.; Amin, S.; Hu, C.D.; Zhang, J.; et al. Autophagosomal Membrane Serves as Platform for Intracellular Death-inducing Signaling Complex (iDISC)-mediated Caspase-8 Activation and Apoptosis. J. Biol. Chem. 2012, 287, 12455–12468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yerlikaya, A.; Okur, E.; Ulukaya, E. The p53-independent induction of apoptosis in breast cancer cells in response to proteasome inhibitor bortezomib. Tumor Biol. 2012, 33, 1385–1392. [Google Scholar] [CrossRef]
- Bersani, F.; Taulli, R.; Accornero, P.; Morotti, A.; Miretti, S.; Crepaldi, T.; Ponzetto, C. Bortezomib-mediated proteasome inhibition as a potential strategy for the treatment of rhabdomyosarcoma. Eur. J. Cancer 2008, 44, 876–884. [Google Scholar] [CrossRef] [PubMed]
- Zhimin, L.; Shuichan, X. ERK1/2 MAP Kinases in Cell Survival and Apoptosis. IUBMB Life 2006, 58, 621–631. [Google Scholar]
- Valenzuela-Salas, L.M.; Girón-Vázquez, N.G.; García-Ramos, J.C.; Torres-Bugarín, O.; Gómez, C.; Pestryakov, A.; Villarreal-Gómez, L.J.; Toledano-Magaña, Y.; Bogdanchikova, N. Antiproliferative and Antitumour Effect of Nongenotoxic Silver Nanoparticles on Melanoma Models. Oxid. Med. Cell. Longev. 2019, 2019, 4528241. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Ramírez, O.U.; Valenzuela-Salas, L.M.; Blanco-Salazar, A.; Rodríguez-Arenas, J.A.; Mier-Maldonado, P.A.; García-Ramos, J.C.; Bogdanchikova, N.; Pestryakov, A.; Toledano-Magaña, Y. Antitumor Activity Against Human Colorectal Adenocarcinoma of Silver Nanoparticles: Influene of [Ag]/[PVP] Ratio. Pharmaceutics 2021, 13, 1000. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Ruiz, B.; Arellano-García, M.E.; Radilla-Chávez, P.; Salas-Vargas, D.S.; Toledano-Magaña, Y.; Casillas-Figueroa, F.; Luna Vazquez-Gomez, R.; Pestryakov, A.; García-Ramos, J.C.; Bogdanchikova, N. Cytokinesis-Block Micronucleus Assay Using Human Lymphocytes as a Sensitive Tool for Cytotoxicity/Genotoxicity Evaluation of AgNPs. ACS Omega 2020, 5, 12005–12015. [Google Scholar] [CrossRef] [PubMed]
- Almanza-Reyes, H.; Moreno, S.; Plascencia-López, I.; Alvarado-Vera, M.; Patrón-Romero, L.; Borrego, B.; Reyes-Escamilla, A.; Valencia-Manzo, D.; Brun, A.; Pestryakov, A.; et al. Evaluation of silver nanoparticles for the prevention of SARS-CoV-2 infection in health workers: In vitro and in vivo. PLoS ONE 2021, 16, e0256401. [Google Scholar] [CrossRef]
- Barberis, E.; Amede, E.; Tavecchia, M.; Marengo, E.; Cittone, M.G.; Rizzi, E.; Pedrinelli, A.R.; Tonello, S.; Minisini, R.; Pirisi, M.; et al. Understanding protection from SARS-CoV-2 using metabolomics. Sci. Rep. 2021, 11, 13796. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Formulation | Consumption of Active Component per Head per Day (µg) |
|---|---|---|
| 1 | Argovit C, 0.06% (metallic silver) | 150 |
| 2 | Triviron, 0.03% | 85.5 |
| 3 | Lauric acid monoglyceride (C12), 0.05% | 100 |
| 4 | Ecocid C, 0.05% | 28,500 |
| Formulation Name and Concentration | Bacteriophage φ6 Concentration, PFU */mL | |||||
|---|---|---|---|---|---|---|
| Initial | After Exposure Time (min). | |||||
| 1 | 5 | 15 | 30 | 60 | ||
| Ecocid 0.05% (500 µg/mL) | 9.6 ± 0.17 × 105 | 0 | 0 | 0 | 0 | 0 |
| Triviron 0.003%, (30 µg/mL) | 1 ± 0.00 × 107 | 0 | 0 | 0 | 0 | 0 |
| Lauric acid monoglyceride (C12), 0.05% | 1 ± 0.57 × 107 | 7.4 + 1.3 × 107 | 1.1 + 0.11 × 107 | 2.0 + 3.3 × 107 | 1.3 + 0.33 × 107 | 8.9 + 6.3 × 106 |
| Argovit, 0.00001% (10 µg/mL) of metallic silver | 4.2 ± 0.15 × 106 | 3.4 + 0.63 × 104 | 6 + 0.115 × 102 | 0 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nefedova, E.; Koptev, V.; Bobikova, A.S.; Cherepushkina, V.; Mironova, T.; Afonyushkin, V.; Shkil, N.; Donchenko, N.; Kozlova, Y.; Sigareva, N.; et al. The Infectious Bronchitis Coronavirus Pneumonia Model Presenting a Novel Insight for the SARS-CoV-2 Dissemination Route. Vet. Sci. 2021, 8, 239. https://doi.org/10.3390/vetsci8100239
Nefedova E, Koptev V, Bobikova AS, Cherepushkina V, Mironova T, Afonyushkin V, Shkil N, Donchenko N, Kozlova Y, Sigareva N, et al. The Infectious Bronchitis Coronavirus Pneumonia Model Presenting a Novel Insight for the SARS-CoV-2 Dissemination Route. Veterinary Sciences. 2021; 8(10):239. https://doi.org/10.3390/vetsci8100239
Chicago/Turabian StyleNefedova, Ekaterina, Vyacheslav Koptev, Anna S. Bobikova, Viktoria Cherepushkina, Tatyana Mironova, Vasily Afonyushkin, Nikolai Shkil, Nikolai Donchenko, Yulia Kozlova, Natalia Sigareva, and et al. 2021. "The Infectious Bronchitis Coronavirus Pneumonia Model Presenting a Novel Insight for the SARS-CoV-2 Dissemination Route" Veterinary Sciences 8, no. 10: 239. https://doi.org/10.3390/vetsci8100239
APA StyleNefedova, E., Koptev, V., Bobikova, A. S., Cherepushkina, V., Mironova, T., Afonyushkin, V., Shkil, N., Donchenko, N., Kozlova, Y., Sigareva, N., Davidova, N., Bogdanchikova, N., Pestryakov, A., & Toledano-Magaña, Y. (2021). The Infectious Bronchitis Coronavirus Pneumonia Model Presenting a Novel Insight for the SARS-CoV-2 Dissemination Route. Veterinary Sciences, 8(10), 239. https://doi.org/10.3390/vetsci8100239