
Roles of WNT6 in Sheep Endometrial Epithelial Cell Cycle Progression and Uterine Glands Organogenesis


Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Animals and Tissues
2.2. Immunofluorescence
2.3. RNA-Seq
2.4. Differential Expression and Functional Enrichment Analysis
2.5. RT-qPCR
2.6. Western Blot Analysis
2.7. Transient Transfection in Endometrial Epithelial Cells (EECs)
2.8. Analysis of Cell Cycle
2.9. Organoid Culture of Ovine UGs
2.10. Statistical Analysis
3. Results
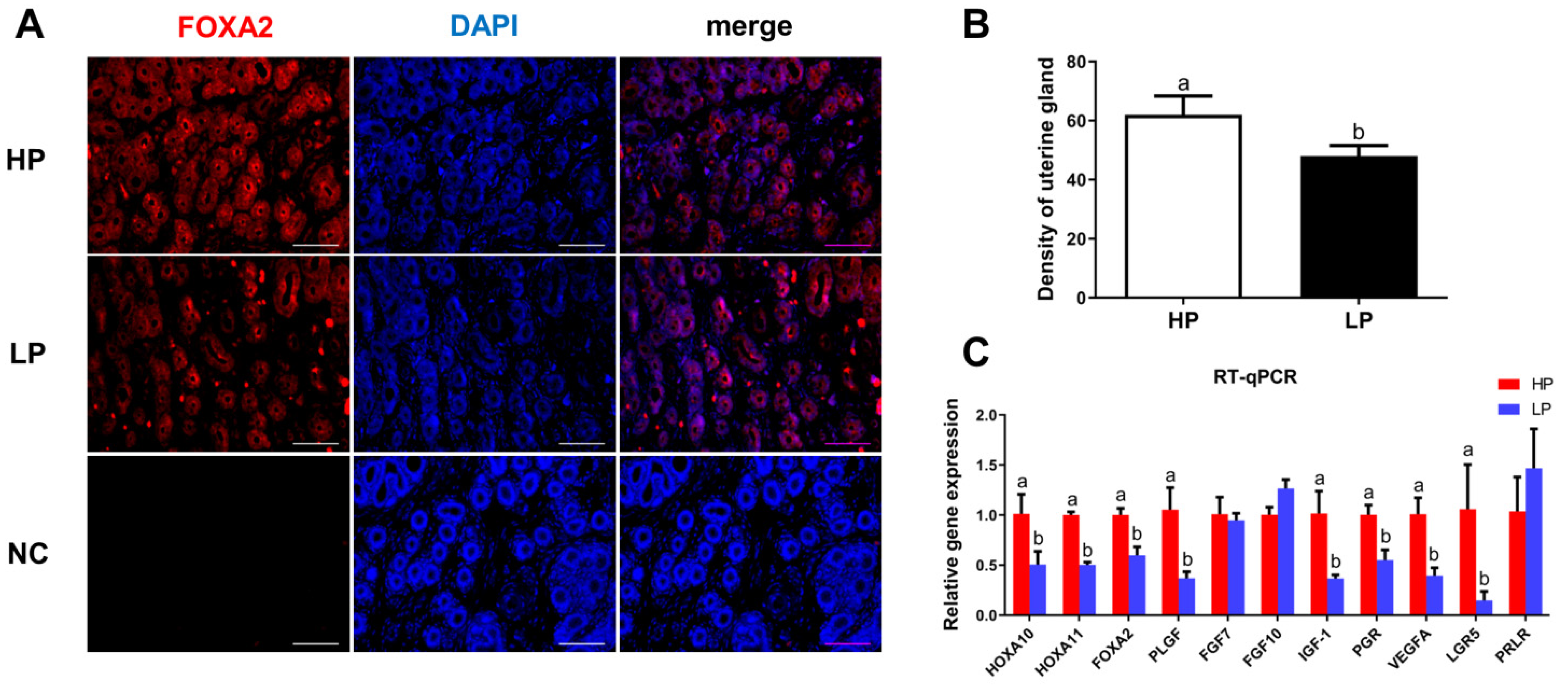
3.1. Analysis of UGs in Hu Sheep with Different Fecundity
3.2. Analysis of DE mRNAs in Endometrium by RNA-Seq
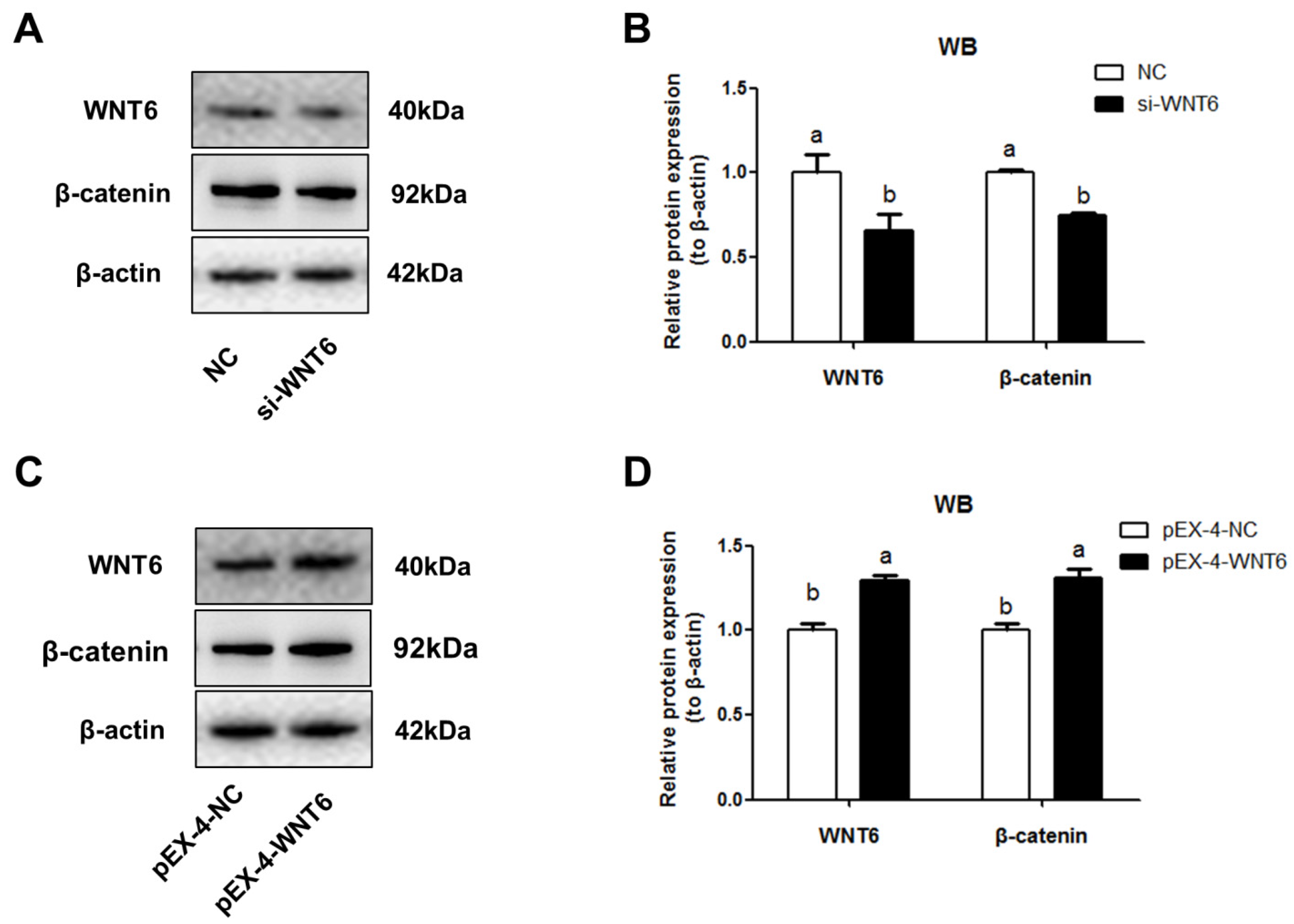
3.3. Effects of WNT6 on β-Catenin Expression in EECs
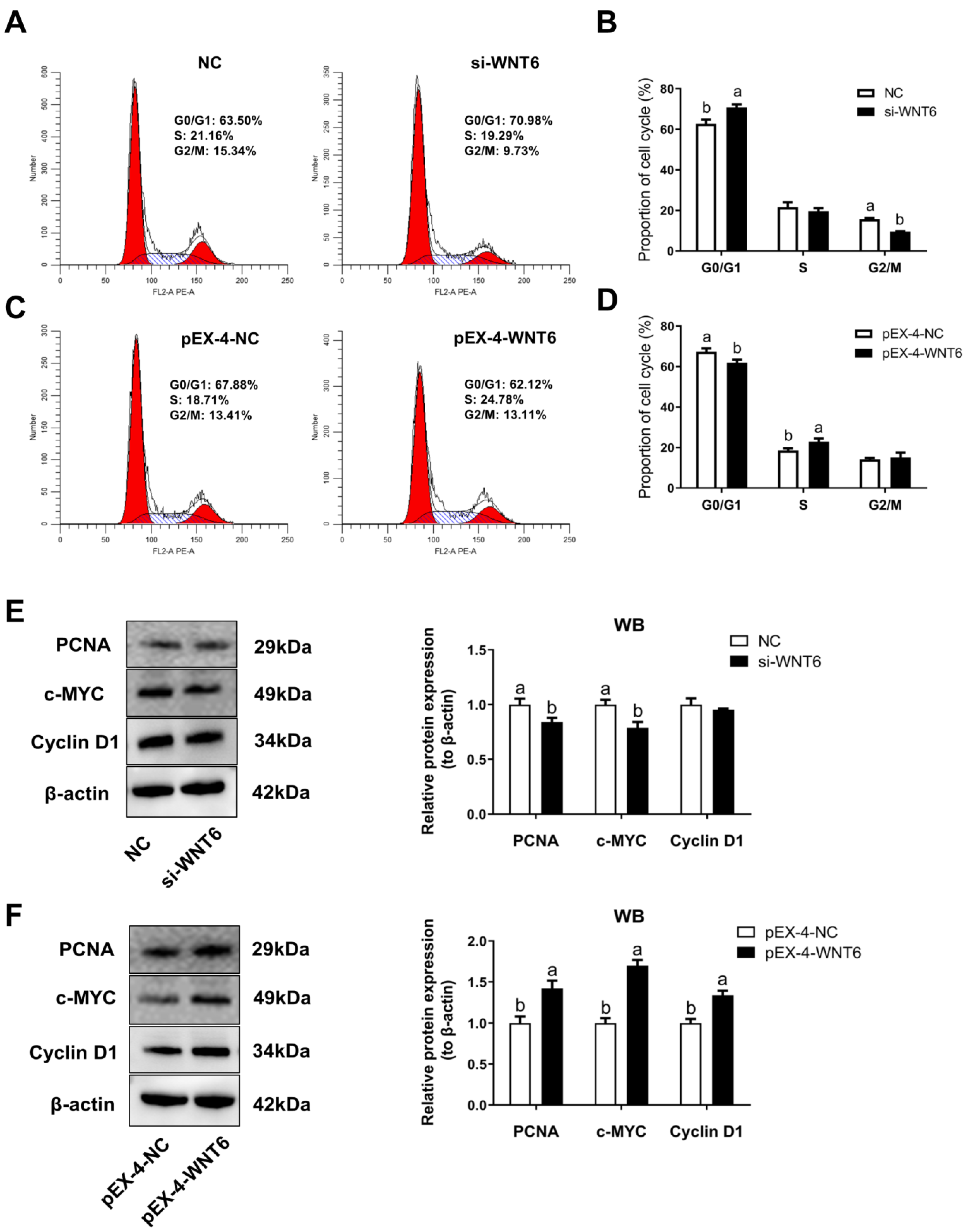
3.4. The Effect of WNT6 Transfection on Cell Cycle of EECs
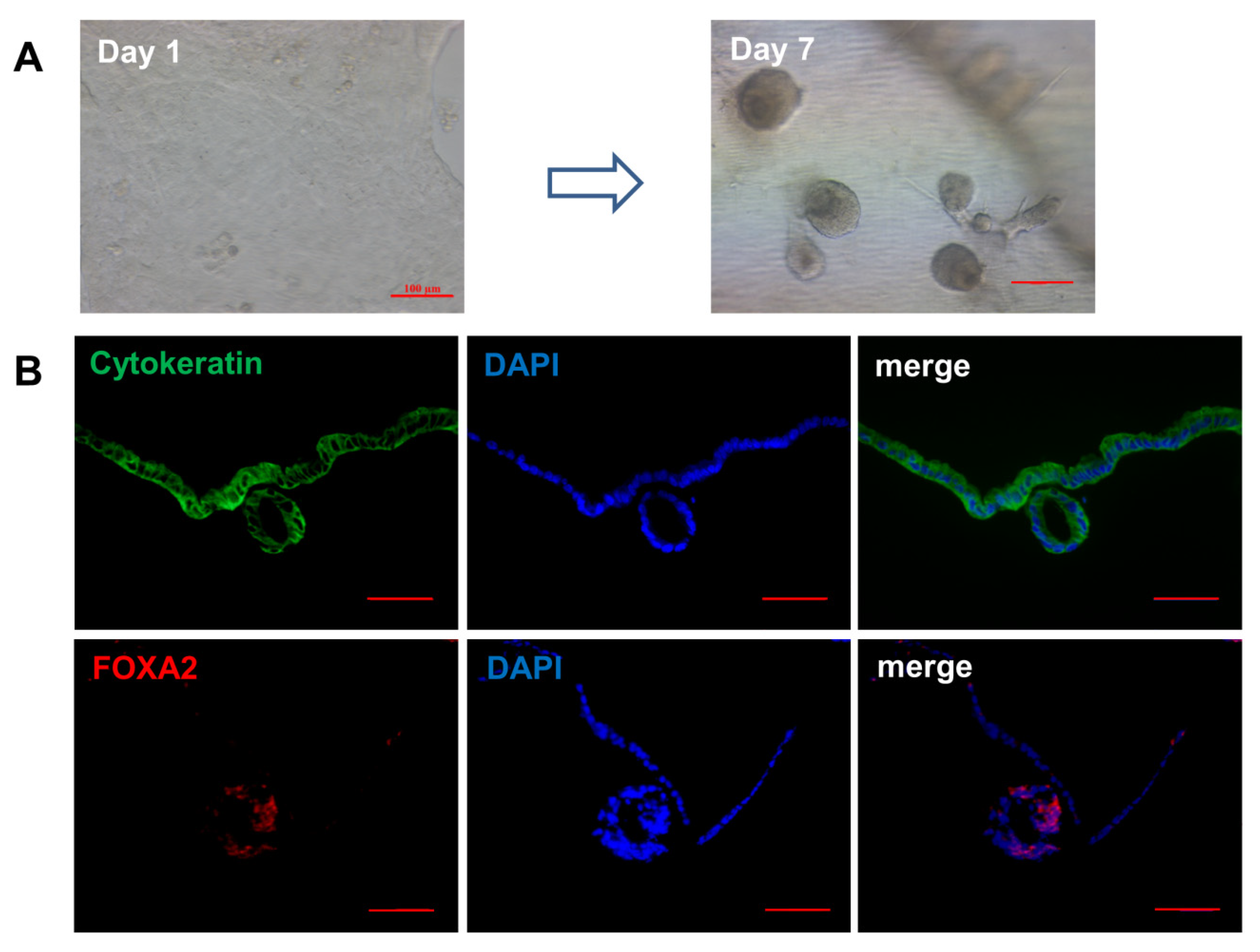
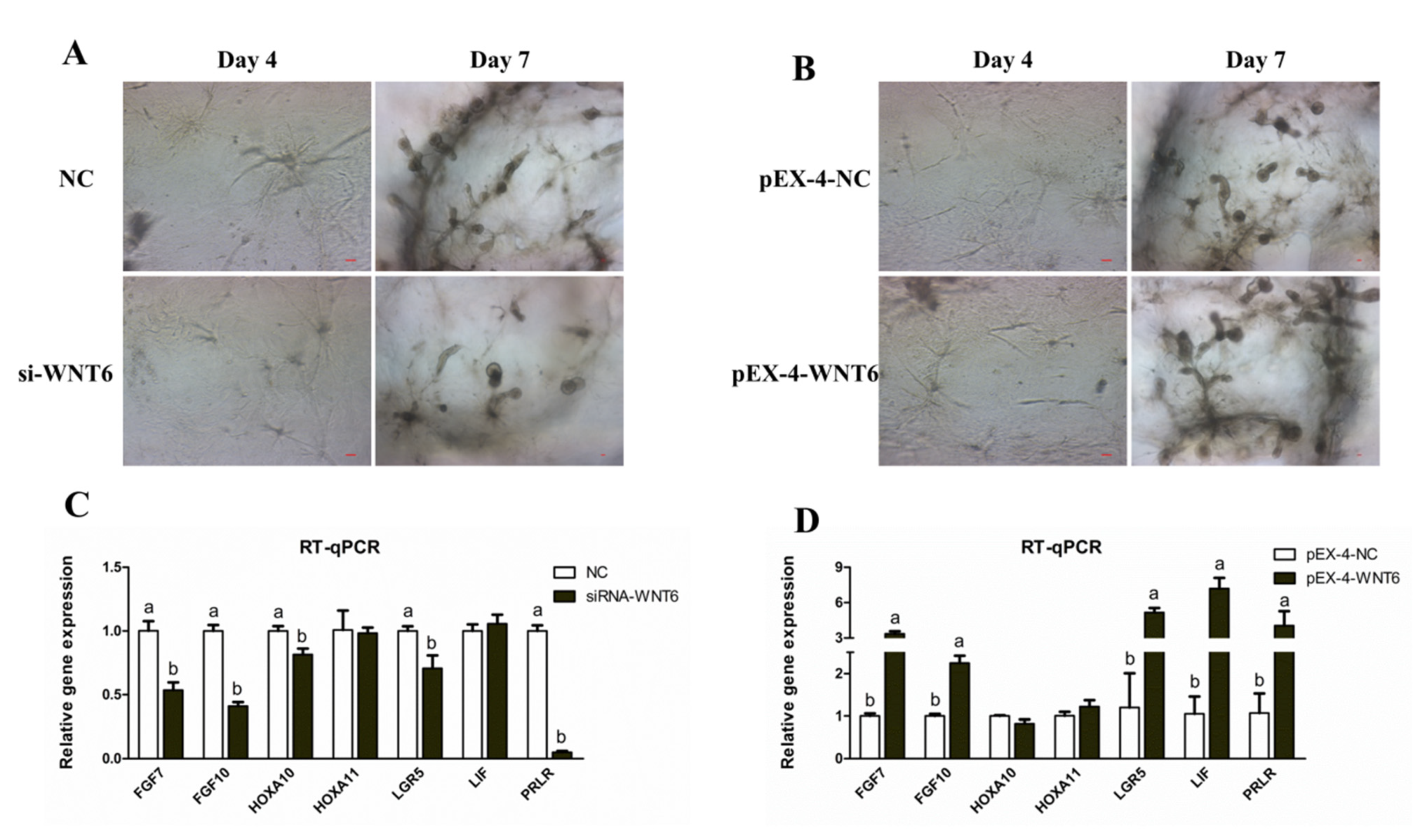
3.5. Effects of WNT6 on Organoid Development of Ovine UGs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yao, X.; Gao, X.; Bao, Y.; El-Samahy, M.A.; Yang, J.; Wang, Z.; Li, X.; Zhang, G.; Zhang, Y.; Liu, W.; et al. Lncrna Fdncr Promotes Apoptosis of Granulosa Cells by Targeting the Mir-543-3p/Dcn/Tgf-Beta Signaling Pathway in Hu Sheep. Mol. Ther. Nucleic Acids 2021, 24, 223–240. [Google Scholar] [CrossRef] [PubMed]
- Spencer, T.E.; Bazer, F.W. Uterine and Placental Factors Regulating Conceptus Growth in Domestic Animals. J. Anim. Sci. 2004, 82, E4–E13. [Google Scholar] [PubMed]
- Gray, C.A.; Bazer, F.W.; Spencer, T.E. Effects of Neonatal Progestin Exposure on Female Reproductive Tract Structure and Function in the Adult Ewe. Biol. Reprod. 2001, 64, 797–804. [Google Scholar] [CrossRef]
- Gray, C.A.; Burghardt, R.C.; Johnson, G.A.; Bazer, F.W.; Spencer, T.E. Evidence That Absence of Endometrial Gland Secretions in Uterine Gland Knockout Ewes Compromises Conceptus Survival and Elongation. Reproduction 2002, 124, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Gray, C.A.; Taylor, K.M.; Ramsey, W.S.; Hill, J.R.; Bazer, F.W.; Bartol, F.F.; Spencer, T.E. Endometrial Glands Are Required for Preimplantation Conceptus Elongation and Survival. Biol. Reprod. 2001, 64, 1608–1613. [Google Scholar] [CrossRef] [Green Version]
- Goad, J.; Ko, Y.-A.; Kumar, M.; Syed, S.; Tanwar, P.S. Differential Wnt Signaling Activity Limits Epithelial Gland Development to the Anti-Mesometrial Side of the Mouse Uterus. Dev. Biol. 2017, 423, 138–151. [Google Scholar] [CrossRef] [PubMed]
- Robinson, G.W.; Hennighausen, L.; Johnson, P.F. Side-Branching in the Mammary Gland: The Progesterone-Wnt Connection. Genes Dev. 2000, 14, 889–894. [Google Scholar] [CrossRef]
- Davies, J.A.; Fisher, C.E. Genes and Proteins in Renal Development. Exp. Nephrol. 2002, 10, 102–113. [Google Scholar] [CrossRef]
- Hayashi, K.; Spencer, T.E. Wnt Pathways in the Neonatal Ovine Uterus: Potential Specification of Endometrial Gland Morphogenesis by Sfrp2. Biol. Reprod. 2006, 74, 721–733. [Google Scholar] [CrossRef] [Green Version]
- Tulac, S.; Nayak, N.R.; Kao, L.C.; Van Waes, M.; Huang, J.; Lobo, S.; Germeyer, A.; Lessey, B.A.; Taylor, R.N.; Suchanek, E.; et al. Identification, Characterization, and Regulation of the Canonical Wnt Signaling Pathway in Human Endometrium. J. Clin. Endocrinol. Metab. 2003, 88, 3860–3866. [Google Scholar] [CrossRef] [Green Version]
- Pollheimer, J.; Loregger, T.; E Sonderegger, S.; Saleh, L.; Bauer, S.; Bilban, M.; Czerwenka, K.; Husslein, P.; Knöfler, M. Activation of the Canonical Wingless/T-Cell Factor Signaling Pathway Promotes Invasive Differentiation of Human Trophoblast. Am. J. Pathol. 2006, 168, 1134–1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kessler, M.; Hoffmann, K.; Brinkmann, V.; Thieck, O.; Jackisch, S.; Toelle, B.; Berger, H.; Mollenkopf, H.-J.; Mangler, M.; Sehouli, J.; et al. The Notch and Wnt Pathways Regulate Stemness and Differentiation in Human Fallopian Tube Organoids. Nat. Commun. 2015, 6, 8989. [Google Scholar] [CrossRef]
- Kim, S.; Choi, Y.; Bazer, F.W.; Spencer, T.E. Identification of Genes in the Ovine Endometrium Regulated by Interferon Tau Independent of Signal Transducer and Activator of Transcription 1. Endocrinology 2003, 144, 5203–5214. [Google Scholar] [CrossRef]
- Zhang, L.N.; Zhao, L.; Yan, X.L.; Huang, Y.H. Loss of G3bp1 Suppresses Proliferation, Migration, and Invasion of Esophageal Cancer Cells Via Wnt/Beta-Catenin and Pi3k/Akt Signaling Pathways. J. Cell. Physiol. 2019, 234, 20469–20484. [Google Scholar] [CrossRef] [PubMed]
- Boretto, M.; Cox, B.; Noben, M.; Hendriks, N.; Fassbender, A.; Roose, H.; Amant, F.; Timmerman, D.; Tomassetti, C.; Vanhie, A.; et al. Development of Organoids from Mouse and Human Endometrium Showing Endometrial Epithelium Physiology and Long-Term Expandability. Development 2017, 144, 1775–1786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, J.; Wang, Z.; Yang, H.; Yao, X.; Yang, P.; Ren, C.; Wang, F.; Zhang, Y. Pituitary Transcriptomic Study Reveals the Differential Regulation of Lncrnas and Mrnas Related to Prolificacy in Different Fecb Genotyping Sheep. Genes 2019, 10, 157. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Li, X.; Zhang, Q.; Zhu, M.; Guo, Y.; Deng, K.; Zhang, G.; Wang, F. Effects of L-Arginine on Endometrial Microvessel Density in Nutrient-Restricted Hu Sheep. Theriogenology 2018, 119, 252–258. [Google Scholar] [CrossRef]
- Gao, X.; Yao, X.; Yang, H.; Deng, K.; Guo, Y.; Zhang, T.; Zhang, G.; Wang, F. Role of Fgf9 in Sheep Testis Steroidogenesis During Sexual Maturation. Anim. Reprod. Sci. 2018, 197, 177–184. [Google Scholar] [CrossRef]
- Kelleher, A.M.; Behura, S.K.; Burns, G.W.; Young, S.L.; Demayo, F.J.; Spencer, T.E. Spencer. Integrative Analysis of the Forkhead Box A2 (Foxa2) Cistrome for the Human Endometrium. FASEB J. 2019, 33, 8543–8554. [Google Scholar] [CrossRef]
- Fitzgerald, H.C.; Dhakal, P.; Behura, S.K.; Schust, D.J.; Spencer, T.E. Self-Renewing Endometrial Epithelial Organoids of the Human Uterus. Proc. Natl. Acad. Sci. USA 2019, 116, 23132–23142. [Google Scholar] [CrossRef]
- Chen, H.; Xia, R.; Li, Z.; Zhang, L.; Xia, C.; Ai, H.; Yang, Z.; Guo, Y. Mesenchymal Stem Cells Combined with Hepatocyte Growth Factor Therapy for Attenuating Ischaemic Myocardial Fibrosis: Assessment Using Multimodal Molecular Imaging. Sci. Rep. 2016, 6, 33700. [Google Scholar] [CrossRef] [Green Version]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript Assembly and Quantification by Rna-Seq Reveals Unannotated Transcripts and Isoform Switching During Cell Differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [Green Version]
- Flegel, C.; Manteniotis, S.; Osthold, S.; Hatt, H.; Gisselmann, G. Expression Profile of Ectopic Olfactory Receptors Determined by Deep Sequencing. PLoS ONE 2013, 8, e55368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anders, S.; Huber, W. Differential Expression Analysis for Sequence Count Data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [Green Version]
- Mao, X.Z.; Cai, T.; Olyarchuk, J.G.; Wei, L.P. Automated Genome Annotation and Pathway Identification Using the Kegg Orthology (Ko) as a Controlled Vocabulary. Bioinformatics 2005, 21, 3787–3793. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T.; et al. Kegg for Linking Genomes to Life and the Environment. Nucleic Acids Res. 2008, 36, 480–484. [Google Scholar] [CrossRef]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene Ontology Analysis for Rna-Seq: Accounting for Selection Bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, X.; Zhang, Q.; Zhu, M.; Li, X.; Guo, Y.; Pang, J.; Liu, Z.; You, P.; Zhang, G.; Wang, F. Effects of L-Arginine on Endometrial Estrogen Receptor Alpha/Beta and Progesterone Receptor Expression in Nutrient-Restricted Sheep. Theriogenology 2019, 138, 137–144. [Google Scholar] [CrossRef]
- Jiang, X.; Cao, X.; Huang, Y.; Chen, J.; Yao, X.; Zhao, M.; Liu, Y.; Meng, J.; Li, P.; Li, Z.; et al. Effects of Treatment with Astragalus Membranaceus on Function of Rat Leydig Cells. BMC Complement. Altern. Med. 2015, 15, 261. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Huang, Y.; Zhou, J.; Men, Y.; Ghazal, S.; Flannery, C.; Müller, M.; Mueller, M.; Mc Kinnon, B.; Taylor, H.S. H19 Lncrna Alters Stromal Cell Growth Via Igf Signaling in the Endometrium of Women with Endometriosis. EMBO Mol. Med. 2015, 7, 996–1003. [Google Scholar]
- Zhang, H.; Peng, A.; Yu, Y.; Guo, S.; Wang, M.; Wang, H. L-Arginine Protects Ovine Intestinal Epithelial Cells from Lipopolysaccharide-Induced Apoptosis through Alleviating Oxidative Stress. J. Agric. Food Chem. 2019, 67, 1683–1690. [Google Scholar] [CrossRef] [PubMed]
- Spencer, T.E.; Hayashi, K.; Hu, J.; Carpenter, K.D. Comparative Developmental Biology of the Mammalian Uterus. Curr. Top. Dev. Biol. 2005, 68, 85–122. [Google Scholar] [CrossRef] [PubMed]
- Kelleher, A.; DeMayo, F.J.; E Spencer, T.E. Uterine Glands: Developmental Biology and Functional Roles in Pregnancy. Endocr. Rev. 2019, 40, 1424–1445. [Google Scholar] [CrossRef] [PubMed]
- Gray, C.A.; Bartol, F.F.; Tarleton, B.J.; Wiley, A.A.; Johnson, G.A.; Bazer, F.W.; Spencer, T.E. Developmental Biology of Uterine Glands. Biol. Reprod. 2001, 65, 1311–1323. [Google Scholar] [CrossRef]
- Knight, J.W.; Bazer, F.W.; Thatcher, W.W.; Franke, D.E.; Wallace, H.D. Conceptus Development in Intact and Unilaterally Hysterectomized-Ovariectomized Gilts: Interrelations among Hormonal Status, Placental Development, Fetal Fluids and Fetal Growth. J. Anim. Sci. 1977, 44, 620–637. [Google Scholar] [CrossRef]
- Binder, N.K.; Evans, J.; Salamonsen, L.; Gardner, D.K.; Kaitu’U-Lino, T.J.; Hannan, N.J. Placental Growth Factor Is Secreted by the Human Endometrium and Has Potential Important Functions During Embryo Development and Implantation. PLoS ONE 2016, 11, e0163096. [Google Scholar] [CrossRef] [Green Version]
- Seishima, R.; Leung, C.; Yada, S.; Murad, K.B.A.; Tan, L.T.; Hajamohideen, A.; Tan, S.H.; Itoh, H.; Murakami, K.; Ishida, Y.; et al. Neonatal Wnt-Dependent Lgr5 Positive Stem Cells Are Essential for Uterine Gland Development. Nat. Commun. 2019, 10, 5378. [Google Scholar] [CrossRef]
- Benson, G.; Lim, H.; Paria, B.; Satokata, I.; Dey, S.; Maas, R. Mechanisms of Reduced Fertility in Hoxa-10 Mutant Mice: Uterine Homeosis and Loss of Maternal Hoxa-10 Expression. Development 1996, 122, 2687–2696. [Google Scholar] [CrossRef]
- Hayashi, K.; Yoshioka, S.; Reardon, S.N.; Rucker, E.B., 3rd; Spencer, T.E.; DeMayo, F.J.; Lydon, J.P.; MacLean, J.A., 2nd. Wnts in the Neonatal Mouse Uterus: Potential Regulation of Endometrial Gland Development. Biol. Reprod. 2011, 84, 308–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Itäranta, P.; Lin, Y.; Peräsaari, J.; Roël, G.; Destrée, O.; Vainio, S. Wnt-6 Is Expressed in the Ureter Bud and Induces Kidney Tubule Development in Vitro. Genesis 2002, 32, 259–268. [Google Scholar] [CrossRef]
- Gao, X.; Yao, X.; Wang, Z.; Li, X.; Li, X.; An, S.; Wei, Z.; Zhang, G.; Wang, F. Long Non-Coding Rna366.2 Controls Endometrial Epithelial Cell Proliferation and Migration by Upregulating Wnt6 as a Cerna of Mir-1576 in Sheep Uterus. Biochim. Biophys. Acta. Gene Regul. Mech. 2020, 1863, 194606. [Google Scholar] [CrossRef] [PubMed]
- Azzolin, L.; Panciera, T.; Soligo, S.; Enzo, E.; Bicciato, S.; Dupont, S.; Bresolin, S.; Frasson, C.; Basso, G.; Guzzardo, V.; et al. Yap/Taz Incorporation in the Beta-Catenin Destruction Complex Orchestrates the Wnt Response. Cell 2014, 158, 157–170. [Google Scholar] [CrossRef] [Green Version]
- van der Horst, P.H.; Wang, Y.; van der Zee, M.; Burger, C.W.; Blok, L.J. Interaction between Sex Hormones and Wnt/Beta-Catenin Signal Transduction in Endometrial Physiology and Disease. Mol. Cell. Endocrinol. 2012, 358, 176–184. [Google Scholar] [CrossRef]
- Cui, T.; He, B.; Kong, S.; Zhou, C.; Zhang, H.; Ni, Z.; Bao, H.; Qiu, J.; Xin, Q.; Reinberg, D.; et al. Pr-Set7 Deficiency Limits Uterine Epithelial Population Growth Hampering Postnatal Gland Formation in Mice. Cell Death Differ. 2017, 24, 2013–2021. [Google Scholar] [CrossRef] [Green Version]
- Davidson, G. The Cell Cycle and Wnt. Cell Cycle 2010, 9, 1667–1668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Pilla, F.; Anderson, S.; Martínez-Escribano, S.; Herrer, I.; Moreno-Moya, J.M.; Musti, S.; Bocca, S.; Oehninger, S.; Horcajadas, J.A. A Novel Model of Human Implantation: 3d Endometrium-Like Culture System to Study Attachment of Human Trophoblast (Jar) Cell Spheroids. Mol. Hum. Reprod. 2021, 18, 33–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibodies | Cat No. | Source | IF | WB | Definition/Function |
|---|---|---|---|---|---|
| PCNA | ab15497 | Abcam, Cambridge, UK | _ | 1:600 | Proliferating cell nuclear antigen, commonly known as PCNA, is a protein that acts as a processivity factor for DNA polymerase δ in eukaryotic cells. |
| WNT6 | ab154144 | Abcam, Cambridge, USA | _ | 1:1000 | The WNT gene family consists of structurally related genes, which encode secreted signaling proteins. |
| β-catenin | 51067-2-AP | Proteintech, Wuhan, China | _ | 1:5000 | β-Catenin, also known as CTNNB1, is a key downstream component of the canonical Wnt pathway that plays diverse and critical roles in embryonic development and adult tissue homeostasis. |
| Cyclin D1 | 26939-1-AP | Proteintech, Wuhan, China | _ | 1:1000 | Cyclin D1, also known as PRAD1 or BCL1, belongs to the highly conserved cyclin family, whose members are characterized by a dramatic periodicity in protein abundance throughout the cell cycle. |
| c-MYC | 10828-1-AP | Proteintech, Wuhan, China | _ | 1:1000 | MYC is a multifunctional, nuclear phosphoprotein that plays a role in cell cycle progression, apoptosis, and cellular transformation. |
| FOXA2 | Ab108422 | Abcam, Cambridge, USA | 1:300 | _ | FOXA2 as a transcription factor involves in embryonic development, establishment of tissue-specific gene expression and regulation of gene expression in differentiated tissues, and it is a mark of epithelial cell |
| Cytokeratin | AF1618 | Beyotime, Shanghai, China | 1:100 | _ | Cytokeratin are a large family of proteins that form the intermediate filament cytoskeleton of epithelial cells. |
| β-actin | bs-0061R | Bioss, Beijing, China | _ | 1:2000 | Actins are highly conserved globular proteins that are involved in various types of cell motility and are ubiquitously expressed in all eukaryotic cells. β-actin is the protein for internal reference. |
| Gene | Forward Primer, 5′-3′ | Reverse Primer, 5′-3′ | Product Size (bp) |
|---|---|---|---|
| HOXA10 | CTTTTCGCGTCCAGAGACCC | ATCCCCGTCACCACTTGACA | 269 |
| HOXA11 | CCAATGACATACTCCTACTCTTCC | GGCTCAATGGCGTATTCTCTG | 83 |
| FOXA2 | GTAGCCGCTCTGGGTCTTAAC | TCGTGCCCTTCCATCTTCAC | 99 |
| PLGF | GCCTTGTCTCCTGGGAACATT | CTCCACAAAGAAGGGCTGGT | 276 |
| FGF7 | TCCTGCCAAGTTTGCTCTAC | CTCACTCTTATATCTCCTCCTTCC | 175 |
| FGF10 | ACCAAGAAGGAGAACTGCCC | TTCGAGCCATAGAGTTTCCCC | 131 |
| IGF-1 | ACCTACACAGGTGAAGATGCC | CTTGAGAGGCGCACAGTACA | 286 |
| PGR | CAGCCAGAGCCCACAGTACA | TGCAATCGTTTCTTCCAGCA | 176 |
| VEGFA | GCCTTGCCTTGCTGCTCTAC | GGTTTCTGCCCTCCTTCTGC | 76 |
| LGR5 | CGAAACGCAAACTCCCTTCC | ATCACGAGGAAGCAAAGCGA | 204 |
| PRLR | CAGGTACGTACAGGGAAGCATTC | GAGTGCTTTTCATTCTGCTACTTTTTC | 66 |
| LIF | ACAGCCTCTTTATCCTCTATTAC | GCGATGATGCGATACAGC | 151 |
| DKK1 | AGGTACCGTCTGTCTTCGCT | CAAGACAGGCCTTCTCCACA | 180 |
| TCF7 | AGGAGACGACAGAGTCCCAA | ATTGAGGGCCCCAGGTTTAG | 124 |
| FRAT2 | TACGGCGTCCTACTTACC | CCACTAACTTCTCGGCTTG | 115 |
| WNT2 | GAACCGCCAAGGACAACAAG | TTCATCAGGGCTCTGGCATC | 133 |
| WNT4 | GCCTTCACGGTGACTCTTCA | GCTTTGCCCTTTGTCCTGTG | 248 |
| WNT5B | CTGAGGACCTGGTGTACGTG | AACGCTCTTGAACTGGTCGT | 164 |
| WNT6 | GTCGAGGCTCTTCATGGACG | GCCCGCCTCGTTGTTATGAA | 85 |
| WNT11 | AAGGACTCGGAGCTCGTCTA | GCGGTCTGTGTAAGGGTTGT | 168 |
| ACTB | TCAGCAAGCAGGAGTACGAC | ACGAGGCCAATCTCATCTCG | 137 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, X.; Yao, X.; Li, X.; Liang, Y.; Liu, Z.; Wang, Z.; Li, K.; Li, Y.; Zhang, G.; Wang, F. Roles of WNT6 in Sheep Endometrial Epithelial Cell Cycle Progression and Uterine Glands Organogenesis. Vet. Sci. 2021, 8, 316. https://doi.org/10.3390/vetsci8120316
Gao X, Yao X, Li X, Liang Y, Liu Z, Wang Z, Li K, Li Y, Zhang G, Wang F. Roles of WNT6 in Sheep Endometrial Epithelial Cell Cycle Progression and Uterine Glands Organogenesis. Veterinary Sciences. 2021; 8(12):316. https://doi.org/10.3390/vetsci8120316
Chicago/Turabian StyleGao, Xiaoxiao, Xiaolei Yao, Xiaodan Li, Yaxu Liang, Zifei Liu, Zhibo Wang, Kang Li, Yingqi Li, Guomin Zhang, and Feng Wang. 2021. "Roles of WNT6 in Sheep Endometrial Epithelial Cell Cycle Progression and Uterine Glands Organogenesis" Veterinary Sciences 8, no. 12: 316. https://doi.org/10.3390/vetsci8120316
APA StyleGao, X., Yao, X., Li, X., Liang, Y., Liu, Z., Wang, Z., Li, K., Li, Y., Zhang, G., & Wang, F. (2021). Roles of WNT6 in Sheep Endometrial Epithelial Cell Cycle Progression and Uterine Glands Organogenesis. Veterinary Sciences, 8(12), 316. https://doi.org/10.3390/vetsci8120316