
Modulation of Hepatic Insulin and Glucagon Signaling by Nutritional Factors in Broiler Chicken

 , , , , ,
, , , , ,  ,
,
Abstract
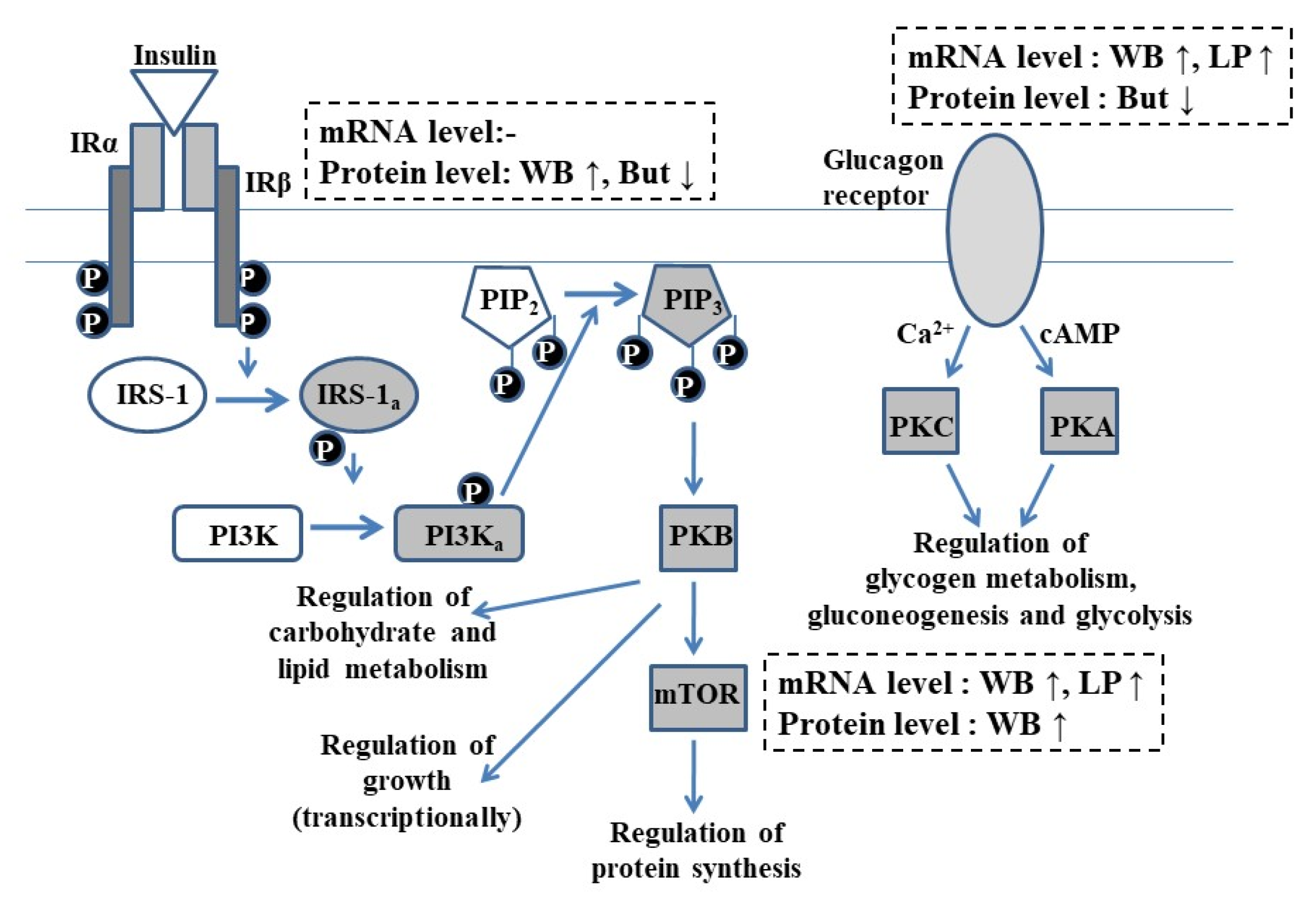
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Sampling
2.3. qRT-PCR Measurements
2.4. Western Blot Measurements
2.5. Statistical Analyses
3. Results
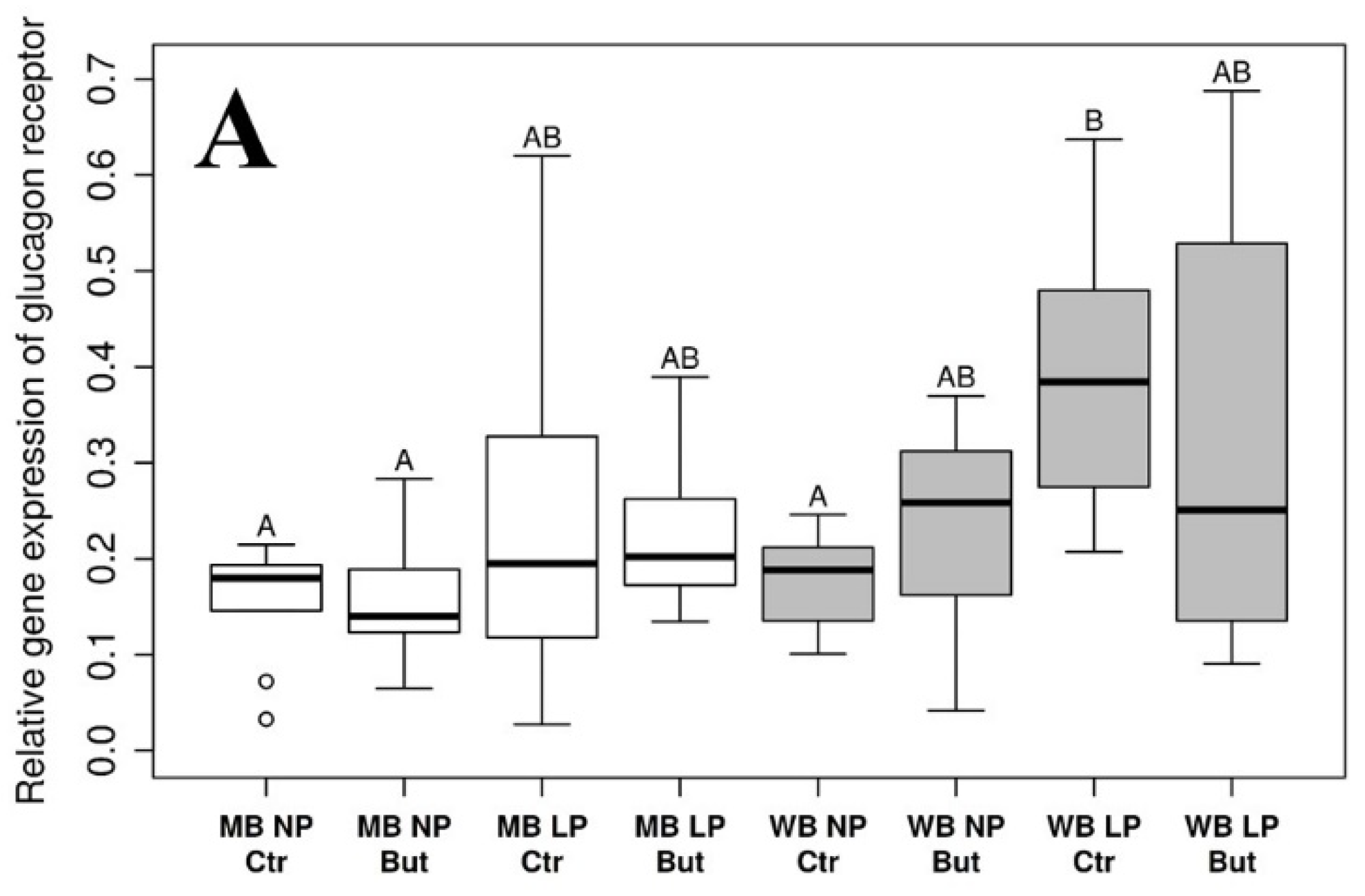
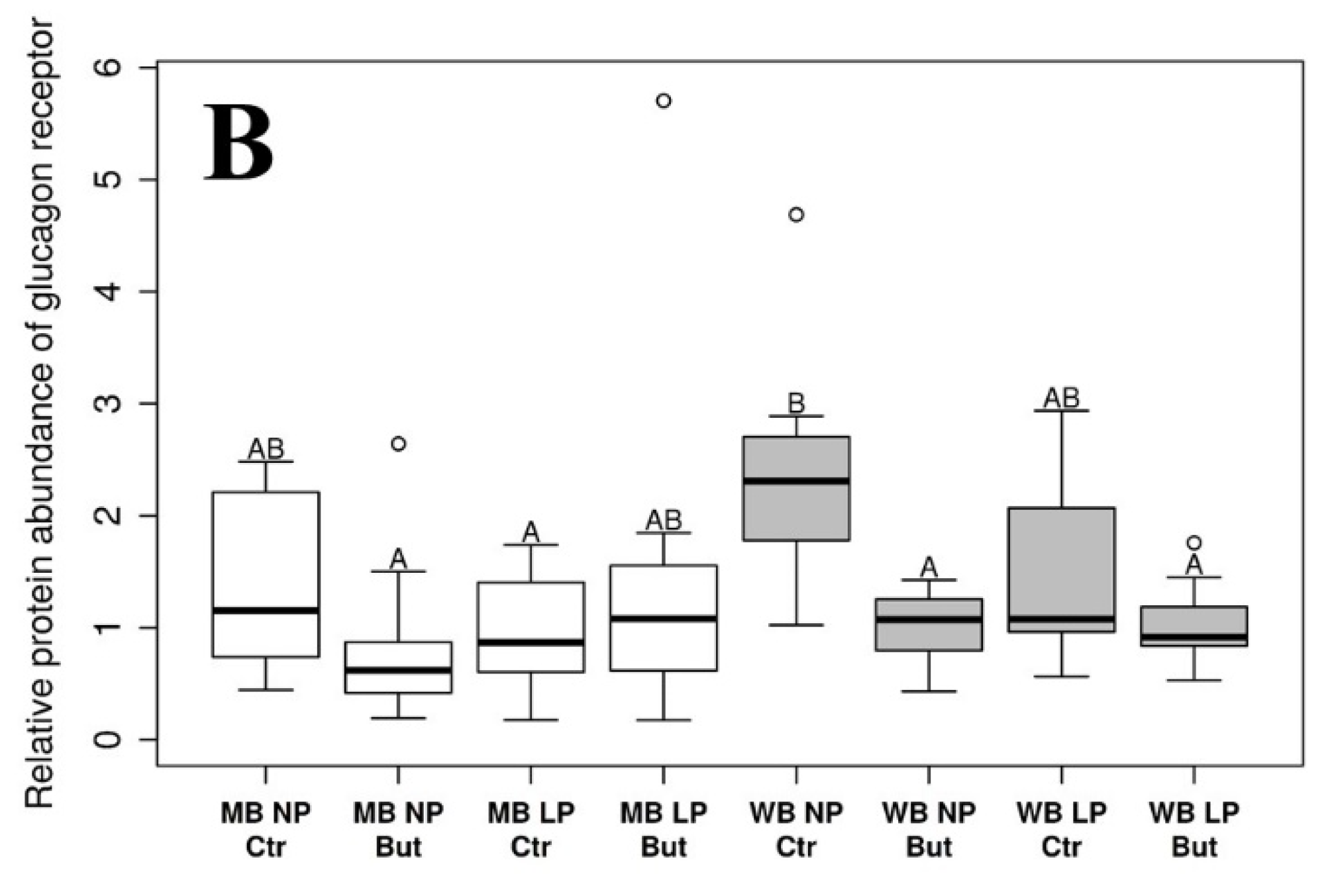
3.1. Glucagon Receptor
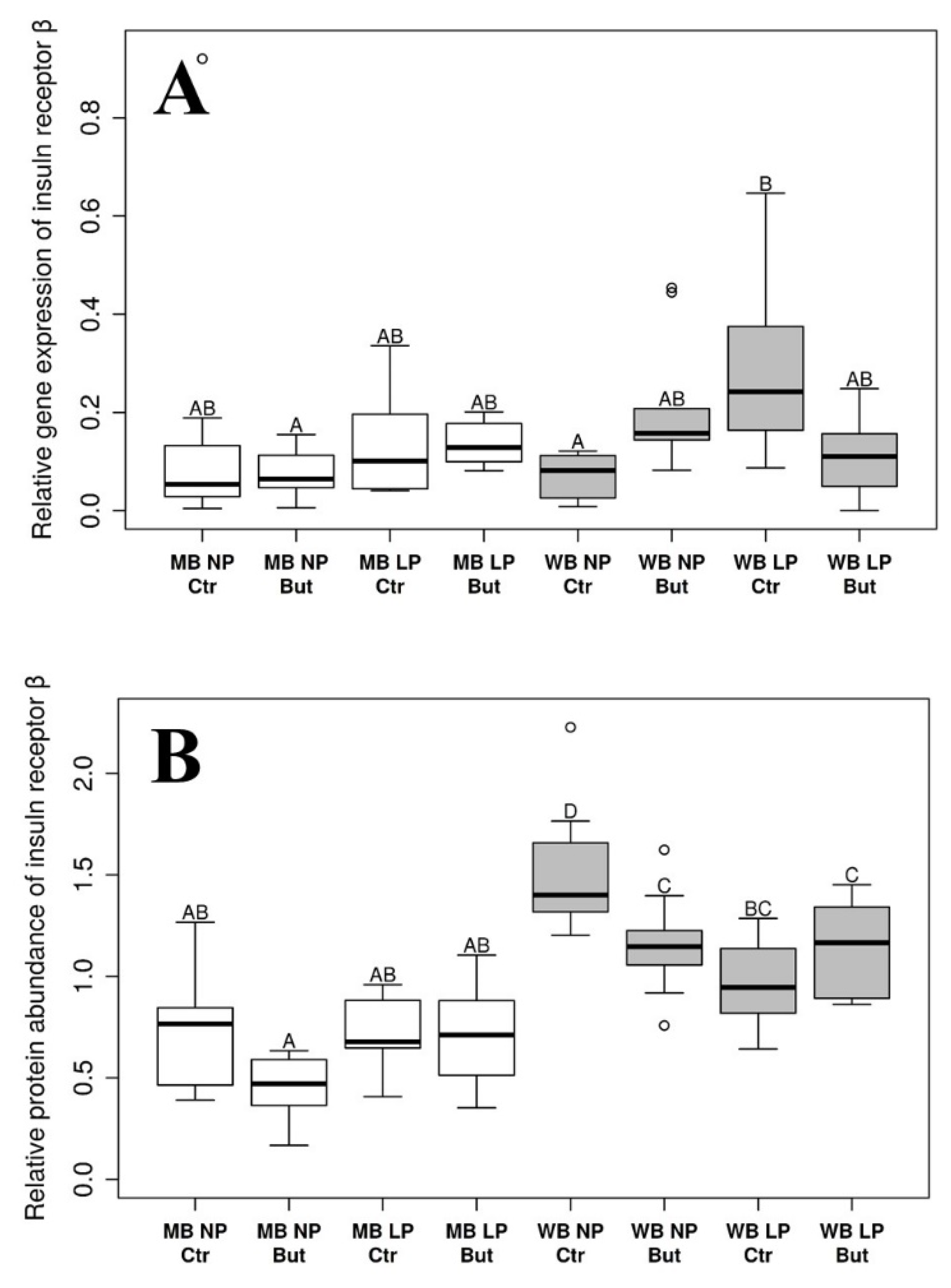
3.2. Insulin Receptor β
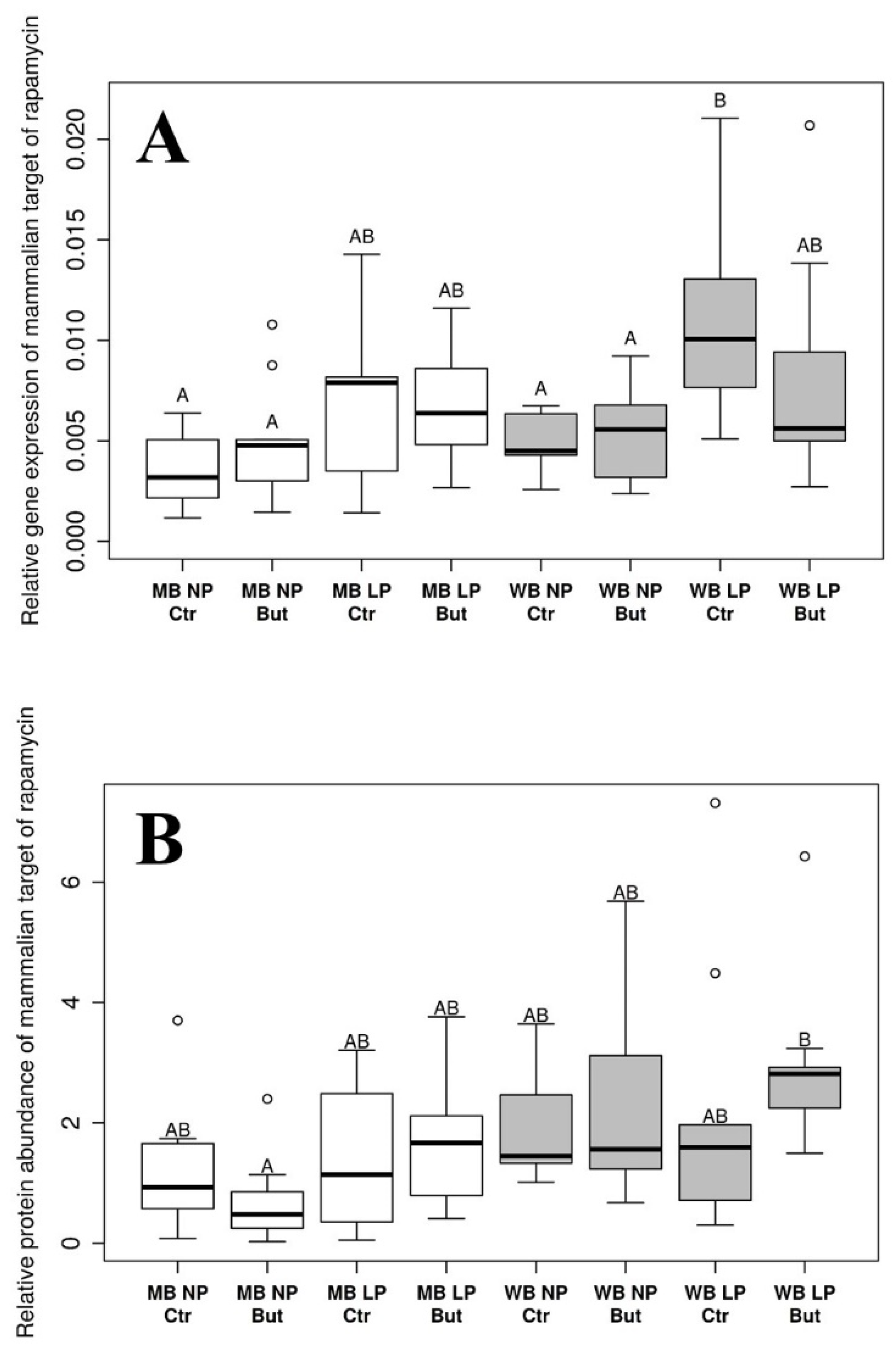
3.3. Mammalian Target of Rapamycin
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yadav, S.; Jha, R. Strategies to modulate the intestinal microbiota and their effects on nutrient utilization, performance, and health of poultry. J. Anim. Sci. Biotechnol. 2019, 10, 2. [Google Scholar] [CrossRef] [PubMed]
- Chamba, F.; Puyalto, M.; Ortiz, A.; Torrealba, H.; Mallo, J.J.; Riboty, R. Effect of partially protected sodium butyrate on performance, digestive organs, intestinal villi and E. coli development in broilers chickens. Int. J. Poult. Sci. 2014, 13, 390–396. [Google Scholar] [CrossRef] [Green Version]
- Svihus, B.; Choct, M.; Classen, H.L. Function and nutritional roles of the avian caeca: A review. World’s Poult. Sci. J. 2013, 69, 249–263. [Google Scholar] [CrossRef]
- Kiarie, E.; Romero, L.F.; Ravindran, V. Growth performance, nutrient utilization, and digesta characteristics in broiler chickens fed corn or wheat diets without or with supplemental xylanase. Poult. Sci. 2014, 93, 1186–1196. [Google Scholar] [CrossRef] [PubMed]
- Molnár, A.; Hess, C.; Pál, L.; Wágner, L.; Awad, W.A.; Husvéth, F.; Hess, M.; Dublecz, K. Composition of diet modifies colonization dynamics of Campylobacter jejuni in broiler chickens. J. Appl. Microbiol. 2015, 118, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Kulcsár, A.; Mátis, G.; Molnár, A.; Petrilla, J.; Husvéth, F.; Huber, K.; Dublecz, K.; Neogrády, Z. Effects of butyrate on the insulin homeostasis of chickens kept on maize- or wheat- based diets. Acta Vet. Hung. 2016, 64, 482–496. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, T.S.; Theil, P.K.; Purup, S.; Nørskov, N.; Knudsen, K.E.B. Effects of Resistant Starch and Arabinoxylan on Parameters Related to Large Intestinal and Metabolic Health in Pigs Fed Fat-Rich Diets. J. Agric. Food Chem. 2015, 63, 10418–10430. [Google Scholar] [CrossRef]
- Bergman, F.N. Energy contributions of volatile fatty acids from the gastrointestinal tract in various species. Physiol. Rev. 1990, 70, 567–590. [Google Scholar] [CrossRef] [Green Version]
- Hu, Z.; Guo, Y. Effects of dietary sodium butyrate supplementation on the intestinal morphological structure, absorptive function and gut flora in chickens. Anim. Feed Sci. Technol. 2007, 132, 240–249. [Google Scholar] [CrossRef]
- Borda-Molina, D.; Mátis, G.; Mackei, M.; Neogrády, Z.; Hubert, K.; Seifert, J.; Camarinha-Silva, A. Caeca Microbial Variation in Broiler Chickens as a Result of Dietary Combinations Using Two Cereal Types, Supplementation of Crude Protein and Sodium Butyrate. Front. Microbiol. 2021, 11, 617800. [Google Scholar] [CrossRef]
- Guilloteau, P.; Martin, L.; Eeckhaut, V.; Ducatelle, R.; Zabielski, R.; Van Immerseel, F. From the gut to the peripheral tissues: The multiple effects of butyrate. Nutr. Res. Rev. 2010, 23, 366–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrera, J.E.; Sakaguchi, K.; Bergel, M.; Trieschmann, L.; Nakatani, Y.; Bustin, M. Specific Acetylation of Chromosomal Protein HMG-17 by PCAF Alters Its Interaction with Nucleosomes. Mol. Cell. Biol. 1999, 19, 3466–3473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braun, H.; Koop, R.; Ertmer, A.; Nacht, S.; Suske, G. Transcription factor Sp3 is regulated by acetylation. Nucleic Acids Res. 2001, 29, 4994–5000. [Google Scholar] [CrossRef] [PubMed]
- Patel, P.; Nankova, B.B.; LaGamma, E.F. Butyrate, a gut-derived environmental signal, regulates tyrosine hydroxylase gene expression via a novel promoter element. Dev. Brain Res. 2005, 160, 53–62. [Google Scholar] [CrossRef]
- Mátis, G.; Neogrády, Z.; Csikó, G.; Gálfi, P.; Fébel, H.; Jemnitz, K.; Veres, Z.; Kulcsár, A.; Kenéz, Á.; Huber, K. Epigenetic effects of dietary butyrate on hepatic histone acetylation and enzymes of biotransformation in chicken. Acta Vet. Hung. 2013, 61, 477–499. [Google Scholar] [CrossRef]
- Mátis, G.; Neogrády, Z.; Csikó, G.; Kulcsár, A.; Kenéz, Á.; Huber, K. Effects of orally applied butyrate bolus on histone acetylation and cytochrome P450 enzyme activity in the liver of chicken–a randomized controlled trial. Nutr. Metab. 2013, 10, 12. [Google Scholar] [CrossRef] [Green Version]
- Thangaraju, M.; Cresci, G.A.; Liu, K.; Ananth, S.; Gnanaprakasam, J.P.; Browning, D.D.; Mellinger, J.D.; Smith, S.B.; Digby, G.J.; Lambert, N.A.; et al. GPR109A is a G-protein–coupled receptor for the bacterial fermentation product butyrate and functions as a tumor suppressor in colon. Cancer Res. 2009, 69, 2826–2832. [Google Scholar] [CrossRef] [Green Version]
- Braun, E.J.; Sweazea, K.L. Glucose regulation in birds. Comp. Biochem. Physiol. B 2008, 151, 1–9. [Google Scholar] [CrossRef]
- Dupont, J.; Tesseraud, S.; Simon, J. Insulin signaling in chicken liver and muscle. Gen. Comp. Endocr. 2009, 163, 52–57. [Google Scholar] [CrossRef]
- Dupont, J.; Dagou, C.; Derouet, M.; Simon, J.; Taouis, M. Early steps of insulin receptor signaling in chicken and rat: Apparent refractoriness in chicken muscle. Domest. Anim. Endocrinol. 2004, 26, 127–142. [Google Scholar] [CrossRef]
- Hazelwood, R.L. The avian endocrine pancreas. Am. Zool. 1973, 13, 699–709. [Google Scholar] [CrossRef]
- Mátis, G.; Kulcsár, A.; Turowski, V.; Fébel, H.; Neogrády, Z.; Huber, K. Effects of oral butyrate application on insulin signaling in various tissues of chickens. Domest. Anim. Endocrinol. 2015, 50, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Boll, E.V.J.; Ekström, L.M.N.K.; Courtin, C.M.; Delcour, J.A.; Nilsson, A.C.; Björck, I.M.E.; Östman, E.M. Effects of wheat bran extract rich in arabinoxylan oligosaccharides and resistant starch on overnight glucose tolerance and markers of gut fermentation in healthy young adults. Eur. J. Nutr. 2016, 55, 1661–1670. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, J.; Watanabe, K.; Taira, S.; Kasubuchi, M.; Li, X.; Irie, J.; Itoh, H.; Kimura, I. Barley β-glucan improves metabolic condition via short-chain fatty acids produced by gut microbial fermentation in high fat diet fed mice. PLoS ONE 2018, 13, e0196579. [Google Scholar] [CrossRef]
- Donsbough, A.L.; Powell, S.; Waguespack, A.; Bidner, T.D.; Southern, L.L. Uric Acid, Urea, and Ammonia Concentrations in Serum and Uric Acid Concentration in Excreta as Indicators of Amino Acid Utilization in Diets for Broilers. Poult. Sci. 2010, 89, 287–294. [Google Scholar] [CrossRef]
- Darsi, E.; Shivazad, M.; Zaghari, M.; Namroud, N.F.; Mohammadi, R. Effect of reduced dietary crude protein levels on growth performance, plasma uric acid and electrolyte concentration of male broiler chicks. J. Agric. Sci. Technol. 2012, 14, 789–797. [Google Scholar]
- Bregendahl, K.; Sell, J.L.; Zimmerman, D.R. Effect of lowprotein diets on growth performance and body composition of broiler chicks. Poult. Sci. 2002, 81, 1156–1167. [Google Scholar] [CrossRef]
- Mátis, G.; Petrilla, J.; Kulcsár, A.; van den Bighelaar, H.; Boomsma, B.; Neogrády, Z.; Fébel, H. Effects of dietary butyrate supplementation and crude protein level on carcass traits and meat composition of broiler chickens. Arch. Anim. Breed. 2019, 62, 527–536. [Google Scholar] [CrossRef] [Green Version]
- Aviagen. Broiler Management Handbook: Ross 308; Aviagen Ltd.: Newbridge, UK, 2018. [Google Scholar]
- Kulcsár, A.; Mátis, G.; Molnár, A.; Petrilla, J.; Wágner, L.; Fébel, H.; Husvéth, F.; Dublecz, K.; Neogrády, Z. Nutritional modulation of intestinal drug-metabolizing cytochrome P450 by butyrate of different origin in chicken. Res. Vet. Sci. 2017, 113, 25–32. [Google Scholar] [CrossRef]
- Kenéz, Á.; Locher, L.; Rehage, J.; Däniche, S.; Huber, K. Agonists of the G protein-coupled receptor 109A-mediated pathway promote antilipolysis by reducing serine residue 563 phosphorylation of hormone-sensitive lipase in bovine adipose tissue explants. J. Dairy Sci. 2014, 97, 3626–3634. [Google Scholar] [CrossRef] [Green Version]
- Douglas, S.G. A rapid method for the determination of pentosans in wheat flour. Food Chem. 1981, 7, 139–145. [Google Scholar] [CrossRef]
- Ingerslev, A.K.; Theil, P.K.; Hedemann, M.S.; Lærke, H.N.; Knudsen, K.E.B. Resistant starch and arabinoxylan augment SCFA absorption, but affect postprandial glucose and insulin responses differently. Br. J. Nutr. 2014, 111, 1564–1576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selvendran, R.R. The plant cell wall as a source of dietary fiber: Chemistry and structure. Am. J. Clin. Nutr. 1984, 39, 320–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hübener, K.; Vahjen, W.; Simon, O. Bacterial responses to different dietary cereal types and xylanase supplementation in the intestine of broiler chicken. Arch. Tierernahr. 2002, 56, 167–187. [Google Scholar] [CrossRef]
- Wang, Z.R.; Qiao, S.Y.; Lu, W.Q.; Li, D.F. Effects of Enzyme Supplementation on Performance, Nutrient Digestibility, Gastrointestinal Morphology, and Volatile Fatty Acid Profiles in the Hindgut of Broilers Fed Wheat-based Diets. Poult. Sci. 2005, 84, 875–881. [Google Scholar] [CrossRef] [PubMed]
- Wu, G. Amino acids: Metabolism, functions, and nutrition. Amino Acids 2009, 37, 1–17. [Google Scholar] [CrossRef]
- Tavares, F.B.; Santos, M.S.V.; Araújo, C.V.; Costa, H.S.; Loureiro, J.P.B.; Lima, E.M.; Lima, K.R.S. Performance, growth and carcass characteristics of alternatives lineages of broiler chickens created with access to paddock. Rev. Bras. Saúde Prod. Anim. 2015, 16, 420–429. [Google Scholar] [CrossRef] [Green Version]
- Murawska, D. Chapter 2: The Effect of Age on Growth Performance and Carcass Quality Parameters in Different Poultry Species. In Poultry Science; Manafi, M., Ed.; IntechOpen: London, UK, 2017; pp. 33–50. [Google Scholar] [CrossRef] [Green Version]
- Gupte, A.A.; Bomhoff, G.L.; Geiger, P.C. Age-related differences in skeletal muscle insulin signaling: The role of stress kinases and heat shock proteins. J. Appl. Physiol. 2008, 105, 839–848. [Google Scholar] [CrossRef] [Green Version]
- Joseph, J.; Dandekar, D.S.; Ramachandran, A.V. Dexamethasone-induced alterations in glucose tolerance and insulin, glucagon and adrenaline responses during the first month in White Leghorn chicks. Br. Poult. Sci. 1996, 37, 665–676. [Google Scholar] [CrossRef]
- Deng, H.; Zheng, A.; Liu, G.; Chang, W.; Zhang, S.; Cai, H. Activation of mammalian target of rapamycin signaling in skeletal muscle of neonatal chicks: Effects of dietary leucine and age. Poult. Sci. 2014, 93, 114–121. [Google Scholar] [CrossRef]
- Oscar, T.P. Down-Regulation of Glucagon Receptors on the Surface of Broiler Adipocytes. Poult. Sci. 1996, 75, 1027–1034. [Google Scholar] [CrossRef] [PubMed]
- Leeson, S.; Namkung, H.; Antongiovanni, M.; Lee, E.H. Effect of butyric acid on the performance and carcass yield of broiler chickens. Poult. Sci. 2005, 84, 1418–1422. [Google Scholar] [CrossRef] [PubMed]
- Dupont, J.; Métayer-Coustard, S.; Ji, B.; Ramé, C.; Gespach, C.; Voy, B.; Simon, J. Characterization of major elements of insulin signaling cascade in chicken adipose tissue: Apparent insulin refractoriness. Gen. Comp. Endocr. 2012, 176, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Petrilla, J.; Mátis, G.; Kulcsár, A.; Talapka, P.; Bíró, E.; Mackei, M.; Fébel, H.; Neogrády, Z. Effect of dietary cereal type, crude protein and butyrate supplementation on metabolic parameters of broilers. Acta Vet. Hung. 2018, 66, 408–425. [Google Scholar] [CrossRef] [PubMed]
- Ramiah, S.K.; Abdullah, N.; Akhmal, M.; Mookiah, S.; Farjam, A.S.; Li, C.W.; Boo, L.J.; Idrus, Z. Effect of feeding less shell, extruded and enzymatically treated palm kernel cake on expression growth-related genes in broiler chickens. Ital. J. Anim. Sci. 2019, 18, 997–1004. [Google Scholar] [CrossRef] [Green Version]
- Józefiak, D.; Ptak, A.; Kaczmarek, S.; Mackowiak, P.; Sassek, M.; Slominski, B.A. Multi-carbohydrase and phytase supplementation improves growth performance and liver insulin receptor sensitivity in broiler chickens fed diets containing full-fat rapeseed. Brit. Poult. Sci. 2010, 89, 1939–1946. [Google Scholar] [CrossRef]
- Yan, H.; Ajuwon, K.M. Butyrate modifies intestinal barrier function in IPEC-J2 cells through a selective upregulation of tight junction proteins and activation of the Akt signaling pathway. PLoS ONE 2017, 12, e0179586. [Google Scholar] [CrossRef]
- Mollica, M.P.; Raso, G.M.; Cavaliere, G.; Trinchese, G.; De Filippo, C.; Aceto, S.; Prisco, M.; Pirozzi, C.; Di Guida, F.; Lama, A.; et al. Butyrate Regulates Liver Mitochondrial Function, Efficiency, and Dynamics in Insulin-Resistant Obese Mice. Diabetes 2017, 66, 1405–1418. [Google Scholar] [CrossRef] [Green Version]
- Endo, H.; Niioka, M.; Kobayashi, N.; Tanaka, M.; Watanabe, T. Butyrate-Producing Probiotics Reduce Nonalcoholic Fatty Liver Disease Progression in Rats: New Insight into the Probiotics for the Gut-Liver Axis. PLoS ONE 2013, 8, e63388. [Google Scholar] [CrossRef] [Green Version]
- Jia, Y.; Gao, G.; Song, H.; Cai, D.; Yang, X.; Zhao, R. Low-protein diet fed to crossbred sows during pregnancy and lactation enhances myostatin gene expression through epigenetic regulation in skeletal muscle of weaning piglets. Eur. J. Nutr. 2016, 55, 1307–1314. [Google Scholar] [CrossRef]
- Pan, S.; Jia, Y.; Yang, X.; Cai, D.; Liu, Z.; Song, H.; Zhao, R. Amino acid starvation-induced autophagy is involved in reduced subcutaneous fat deposition in weaning piglets derived from sows fed low-protein diet during gestation and lactation. Eur. J. Nutr. 2018, 57, 991–1001. [Google Scholar] [CrossRef] [PubMed]
- Adams, K.A.; Davis, A.J. Dietary Protein Concentration Regulates the mRNA Concentration of Chicken Hepatic Malic Enzyme. J. Nutr. 2001, 131, 2269–2274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kita, K.; Nagao, K.; Taneda, N.; Inagaki, Y.; Hirano, K.; Shibata, T.; Yaman, M.A.; Conlon, M.A.; Okumura, J. Insulin-Like Growth Factor Binding Protein-2 Gene Expression Can Be Regulated by Diet Manipulation in Several Tissues of Young Chickens. J. Nutr. 2002, 132, 145–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Unit | Maize-Based Normal CP | Maize-Based Low CP | Wheat-Based Normal CP | Wheat-Based Low CP |
|---|---|---|---|---|---|
| Maize | % | 57.60 | 61.00 | 0 | 0 |
| Wheat | % | 0 | 0 | 54.79 | 62.60 |
| Solvent extr. soybean meal | % | 27.00 | 28.00 | 31.00 | 26.48 |
| PL-68 † | % | 6.50 | 0 | 3.00 | 0 |
| Sunflower oil | % | 3.50 | 3.50 | 6.00 | 5.30 |
| Wheat bran | % | 0 | 1.72 | 0 | 0 |
| Limestone | % | 1.70 | 1.60 | 1.70 | 1.70 |
| MCP | % | 1.80 | 2.00 | 1.70 | 1.70 |
| Salt (NaCl) | % | 0.40 | 0.40 | 0.40 | 0.40 |
| L-lysine hydrochloride | % | 0.44 | 0.58 | 0.38 | 0.60 |
| DL-methionine | % | 0.43 | 0.44 | 0.41 | 0.45 |
| L-threonine | % | 0.09 | 0.22 | 0.11 | 0.26 |
| L-tryptophan | % | 0.04 | 0.04 | 0 | 0 |
| Vitamin and mineral premix ‡ | % | 0.50 | 0.50 | 0.50 | 0.50 |
| Axtra XB 201 enzyme § | % | 0.015 | 0.015 | ||
| Calculated values | |||||
| Dry matter | % | 89.65 | 89.32 | 89.78 | 89.47 |
| Crude protein | % | 22.02 | 18.65 | 22.05 | 18.76 |
| Soluble NSP | Mg/kg | 506.88 | 536.80 | 5133.82 | 5865.62 |
| ME | MJ/kg | 12.65 | 12.61 | 12.63 | 12.62 |
| Ether extract | % | 6.54 | 6.30 | 7.49 | 6.62 |
| Crude fiber | % | 2.51 | 2.74 | 2.88 | 2.81 |
| Ash | % | 6.97 | 7.23 | 7.37 | 7.42 |
| Lysine | % | 1.43 | 1.43 | 1.44 | 1.43 |
| Methionine + Cystine | % | 1.07 | 1.05 | 1.08 | 1.07 |
| Threonine | % | 0.97 | 0.94 | 0.94 | 0.94 |
| Tryptophan | % | 0.23 | 0.25 | 0.26 | 0.24 |
| Arginine | % | 1.17 | 1.24 | 1.34 | 1.22 |
| Isoleucine | % | 0.74 | 0.78 | 0.85 | 0.78 |
| Leucine | % | 1.59 | 1.68 | 1.52 | 1.41 |
| Valine | % | 0.83 | 0.88 | 0.93 | 0.86 |
| Total Ca | % | 1.15 | 1.15 | 1.16 | 1.14 |
| Total P | % | 0.79 | 0.80 | 0.82 | 0.80 |
| Available P | % | 0.54 | 0.53 | 0.56 | 0.54 |
| Ingredients | Unit | Maize-Based Normal CP | Maize-Based Low CP | Wheat-Based Normal CP | Wheat-Based Low CP |
|---|---|---|---|---|---|
| Maize | % | 60.71 | 65.31 | 0 | 0 |
| Wheat | % | 0 | 0 | 61.30 | 66.56 |
| Solvent extr. soybean meal | % | 22.20 | 24.54 | 19.31 | 20.01 |
| PL-68 † | % | 8.00 | 1.00 | 8.50 | 2.50 |
| Sunflower oil | % | 4.80 | 4.50 | 6.70 | 6.50 |
| Wheat bran | % | 0 | 0 | 0 | 0 |
| Limestone | % | 1.30 | 1.20 | 1.35 | 1.35 |
| MCP | % | 1.35 | 1.60 | 1.15 | 1.15 |
| Salt (NaCl) | % | 0.40 | 0.40 | 0.40 | 0.40 |
| L-lysine hydrochloride | % | 0.34 | 0.41 | 0.38 | 0.48 |
| DL-methionine | % | 0.36 | 0.37 | 0.35 | 0.38 |
| L-threonine | % | 0 | 0.15 | 0.05 | 0.16 |
| L-tryptophan | % | 0.04 | 0.02 | 0 | 0 |
| Vitamin and mineral premix ‡ | % | 0.50 | 0.50 | 0.50 | 0.50 |
| Axtra XB 201 enzyme § | % | 0,015 | 0,015 | ||
| Calculated values | |||||
| Dry matter | % | 89.72 | 89.34 | 89.90 | 89.55 |
| Crude protein | % | 21.12 | 17.85 | 21.10 | 17.89 |
| Soluble NSP | Mg/kg | 534.25 | 574.73 | 5743.81 | 6236.67 |
| ME | MJ/kg | 13.27 | 13.24 | 13.24 | 13.24 |
| Ether extract | % | 7.96 | 7.39 | 8.45 | 7.92 |
| Crude fiber | % | 2.34 | 2.48 | 2.51 | 2.61 |
| Ash | % | 5.78 | 6.03 | 6.00 | 6.13 |
| Lysine | % | 1.25 | 1.22 | 1.25 | 1.22 |
| Methionine + Cystine | % | 0.96 | 0.95 | 0.94 | 0.95 |
| Threonine | % | 0.84 | 0.84 | 0.85 | 0.81 |
| Tryptophan | % | 0.21 | 0.20 | 0.20 | 0.21 |
| Arginine | % | 1.01 | 1.11 | 0.97 | 1.02 |
| Isoleucine | % | 0.65 | 0.72 | 0.62 | 0.65 |
| Leucine | % | 1.45 | 1.58 | 1.14 | 1.20 |
| Valine | % | 0.74 | 0.81 | 0.70 | 0.74 |
| Total Ca | % | 0.92 | 0.93 | 0.90 | 0.90 |
| Total P | % | 0.68 | 0.69 | 0.71 | 0.67 |
| Available P | % | 0.45 | 0.45 | 0.49 | 0.44 |
| Gene | Primer | Primer Sequence | Amplicon Size | NCBI Accession | Threshold |
|---|---|---|---|---|---|
| GAPDH | Forward (5′-3′) | GGGCACGCCATCACTATCTT | 187 | NM204305.1 | 0.03 |
| Reverse (5′-3′) | TCACAAACATGGGGGCATCA | ||||
| GCGR | Forward (5′-3′) | ATCCCGTGGGTTGTTGTGAA | 195 | NM001101035.1 | 0.02 |
| Reverse (5′-3′) | CTTGTAGTCGGTGTAGCGCA | ||||
| IRβ | Forward (5′-3′) | CAACCCACACTGGTGGTCAT | 134 | XM 001233398.5 | 0.0036 |
| Reverse (5′-3′) | GCAGCCATCTGGATCATTTCTC | ||||
| mTOR | Forward (5′-3′) | GTGGCGATCCTATGGCATGA | 276 | XM417614.6 | 0.05 |
| Reverse (5′-3′) | ACGCCTGAAAACGTGGTAGT |
| Parameter | MB NP Ctr | MB NP But | MB LP Ctr | MB LP But | WB NP Ctr | WB NP But | WB LP Ctr | WB LP But |
|---|---|---|---|---|---|---|---|---|
| GR |  |  |  |  |  |  |  |  |
| IRβ |  |  |  |  |  |  |  |  |
| mTOR |  |  |  |  |  |  |  |  |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petrilla, J.; Mátis, G.; Mackei, M.; Kulcsár, A.; Sebők, C.; Papp, M.; Gálfi, P.; Fébel, H.; Huber, K.; Neogrády, Z. Modulation of Hepatic Insulin and Glucagon Signaling by Nutritional Factors in Broiler Chicken. Vet. Sci. 2022, 9, 103. https://doi.org/10.3390/vetsci9030103
Petrilla J, Mátis G, Mackei M, Kulcsár A, Sebők C, Papp M, Gálfi P, Fébel H, Huber K, Neogrády Z. Modulation of Hepatic Insulin and Glucagon Signaling by Nutritional Factors in Broiler Chicken. Veterinary Sciences. 2022; 9(3):103. https://doi.org/10.3390/vetsci9030103
Chicago/Turabian StylePetrilla, Janka, Gábor Mátis, Máté Mackei, Anna Kulcsár, Csilla Sebők, Márton Papp, Péter Gálfi, Hedvig Fébel, Korinna Huber, and Zsuzsanna Neogrády. 2022. "Modulation of Hepatic Insulin and Glucagon Signaling by Nutritional Factors in Broiler Chicken" Veterinary Sciences 9, no. 3: 103. https://doi.org/10.3390/vetsci9030103
APA StylePetrilla, J., Mátis, G., Mackei, M., Kulcsár, A., Sebők, C., Papp, M., Gálfi, P., Fébel, H., Huber, K., & Neogrády, Z. (2022). Modulation of Hepatic Insulin and Glucagon Signaling by Nutritional Factors in Broiler Chicken. Veterinary Sciences, 9(3), 103. https://doi.org/10.3390/vetsci9030103