
ProAKAP4 Semen Concentrations as a Valuable Marker Protein of Post-Thawed Semen Quality and Bull Fertility: A Retrospective Study

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sperm Descriptors Analysis in Post-Thawed Straws
2.2. Semen Sample Preparation
2.3. Separation of Spermatozoa from the Cryopreservative
2.4. Western-Blotting
2.5. The Bull 4MID® ProAKAP4 ELISA Assay to Quantify Sperm proAKAP4
2.6. Calculation of Non-Return Rate at 90 Days after Artificial Insemination
2.7. Statistical Analysis
3. Results
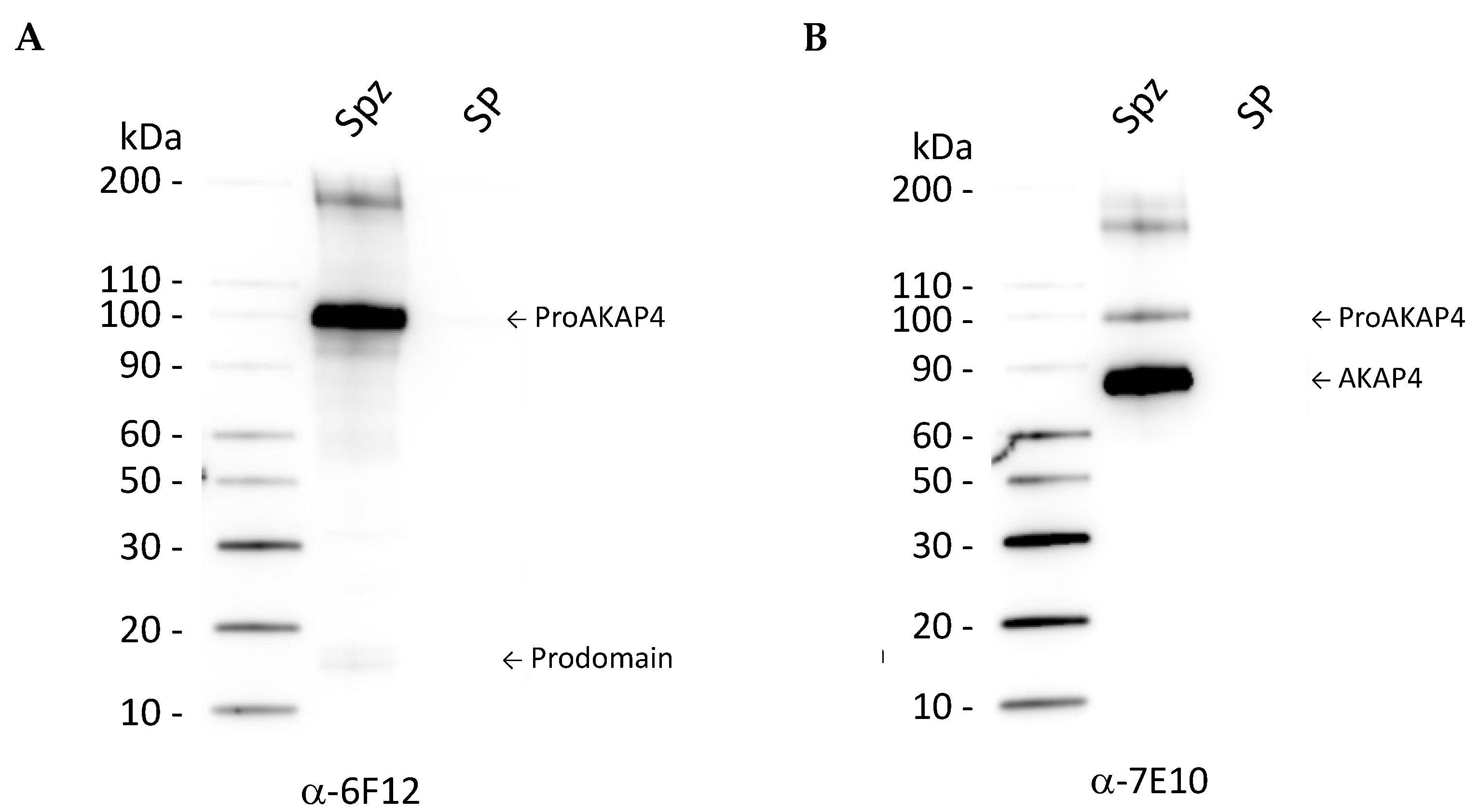
3.1. Analysis of AKAP4 and proAKAP4 Expression in Post-Thawed Isolated Spermatozoa
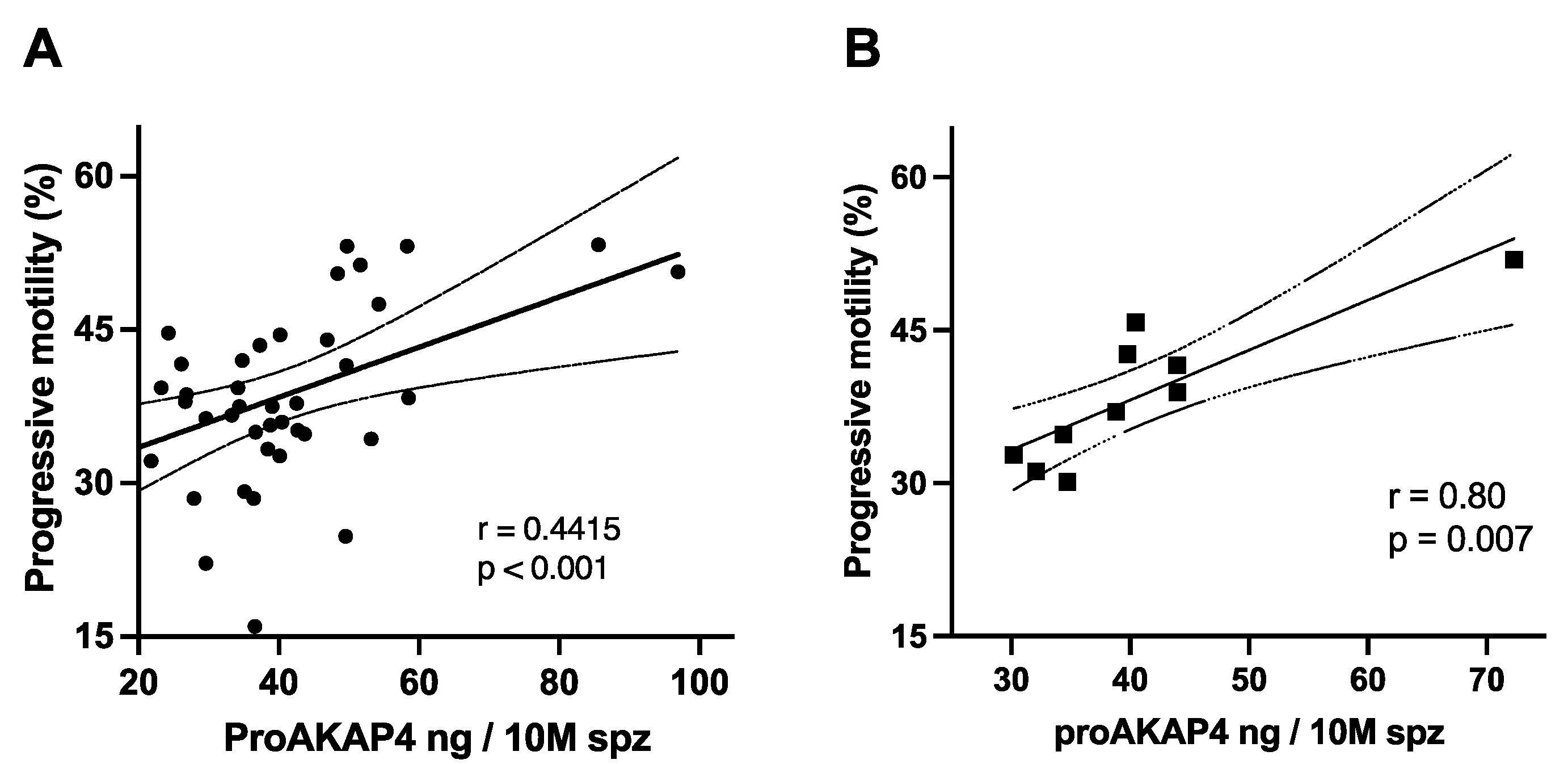
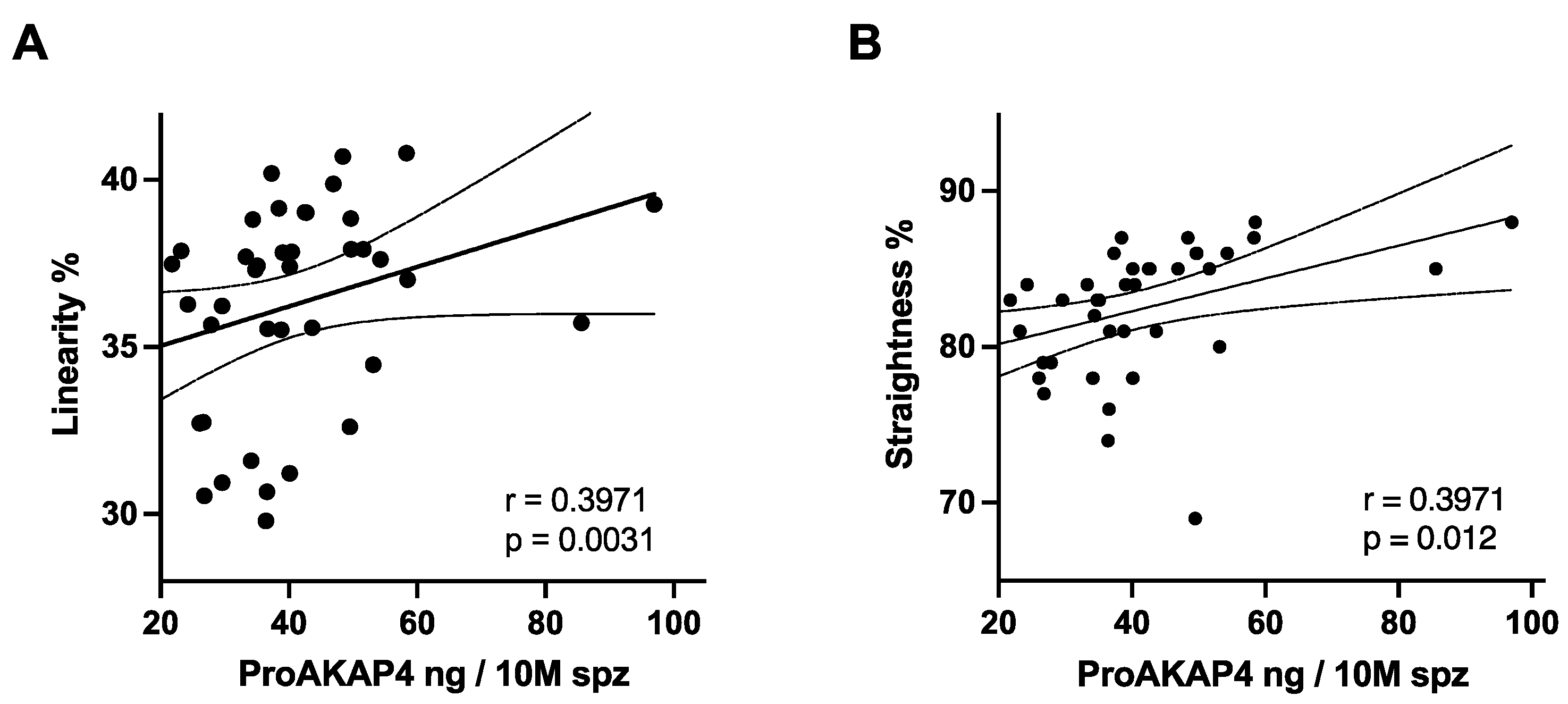
3.2. ProAKAP4 Concentrations and Total Sperm Motility
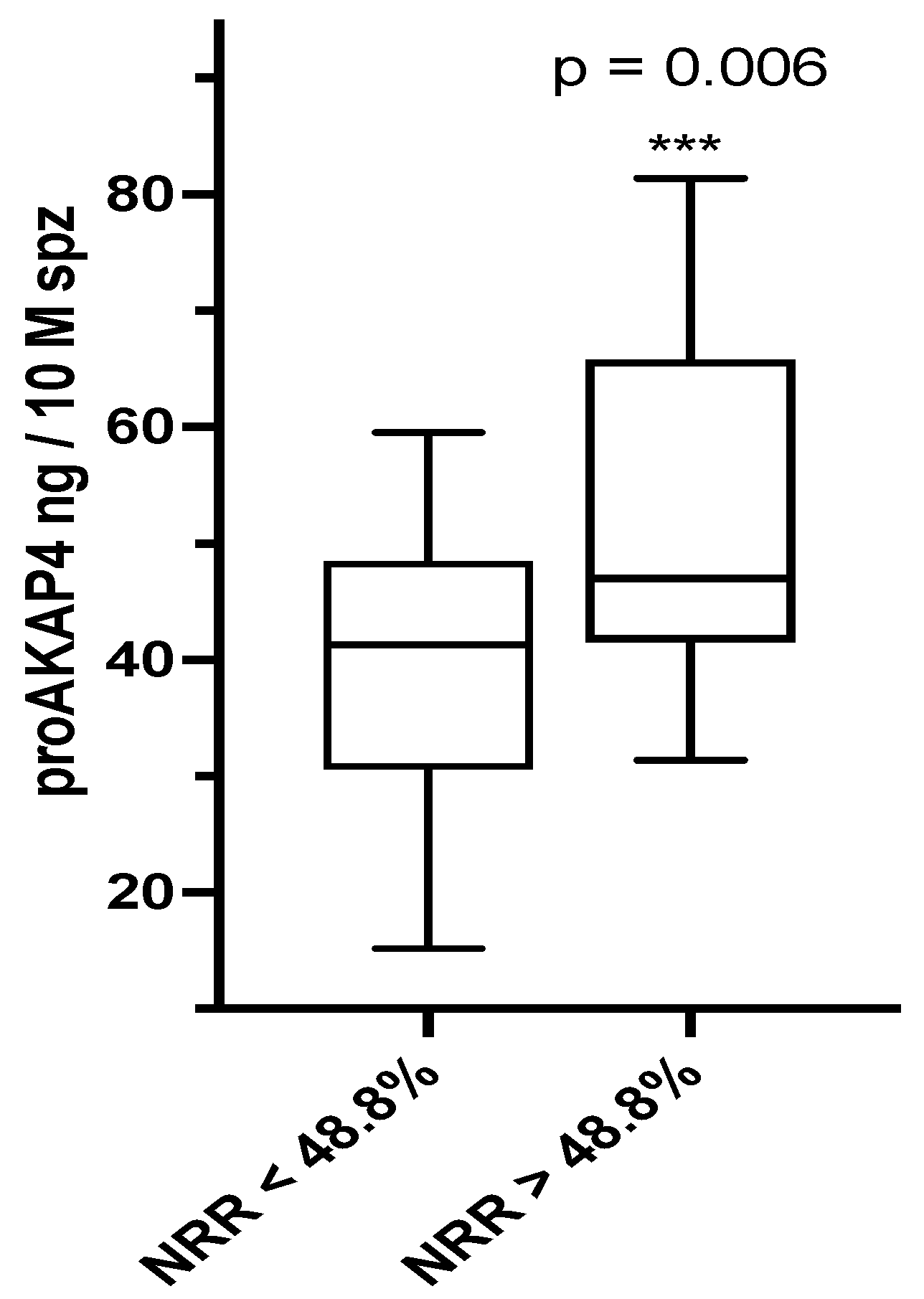
3.3. proAKAP4 Dosage and Fertility Rates
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Inchaisri, C.; Jorritsma, R.; Vos, P.L.A.M.; van der Weijden, G.C.; Hogeveen, H. Economic consequences of reproductive performance in dairy cattle. Theriogenology 2010, 74, 835–846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braundmeier, A.G.; Miller, D.J. The search is on: Finding accurate molecular markers of male fertility. J. Dairy Sci. 2001, 84, 1915–1925. [Google Scholar] [CrossRef]
- Mapel, X.M.; Hiltpold, M.; Kadri, N.K.; Witschi, U.; Pausch, H. Bull fertility and semen quality are not correlated with dairy and production traits in Brown Swiss cattle. JDS Commun. 2022, 3, 120–125. [Google Scholar] [CrossRef]
- Zhang, B.R.; Larsson, B.; Lundeheim, N.; Håård, M.G.; Rodriguez-Martinez, H. Prediction of bull fertility by combined in vitro assessments of frozen-thawed semen from young dairy bulls entering an AI-programme. Int. J. Androl. 1999, 22, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Kumaresan, A.; Johannisson, A.; Al-Essawe, E.M.; Morrell, J.M. Sperm viability, reactive oxygen species, and DNA fragmentation index combined can discriminate between above-and below-average fertility bulls. J. Dairy Sci. 2017, 100, 5824–5836. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, L.Z.; de Arruda, R.P.; de Andrade, A.F.C.; Celeghini, E.C.C.; dos Santos, R.M.; Beletti, M.E.; Peres, R.F.G.; Oliveira, C.S.; de Lima, V.F.M.H. Assessment of field fertility and several in vitro sperm characteristics following the use of different Angus sires in a timed-AI program with suckled Nelore cows. Livest. Sci. 2012, 146, 38–46. [Google Scholar] [CrossRef] [Green Version]
- Narud, B.; Khezri, A.; Nordborg, A.; Klinkenberg, G.; Zeremichael, T.T.; Stenseth, E.-B.; Heringstad, B.; Kommisrud, E.; Myromslien, F.D. Semen quality parameters including metabolites, sperm production traits and fertility in young Norwegian Red AI bulls. Livest. Sci. 2022, 255, 104803. [Google Scholar] [CrossRef]
- Amann, R.P.; DeJarnette, J.M. Impact of genomic selection of AI dairy sires on their likely utilization and methods to estimate fertility: A paradigm shift. Theriogenology 2012, 77, 795–817. [Google Scholar] [CrossRef]
- Donnellan, E.M.; Lonergan, P.; Meade, K.G.; Fair, S. An ex-vivo assessment of differential sperm transport in the female reproductive tract between high and low fertility bulls. Theriogenology 2022, 181, 42–49. [Google Scholar] [CrossRef]
- Schenk, J.L. Principles of maximizing bull semen production at genetic centers. Animal 2018, 12, s142–s147. [Google Scholar] [CrossRef] [Green Version]
- Berry, D.P.; Wall, E.; Pryce, J.E. Genetics and genomics of reproductive performance in dairy and beef cattle. Animal 2014, 8, 105–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butler, M.L.; Bormann, J.M.; Weaber, R.L.; Grieger, D.M.; Rolf, M.M. Selection for bull fertility: A review. Transl. Anim. Sci. 2019, 4, 423–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hopper, R.M. Evaluation of breeding soundness: Basic examination of the semen. In Bovine Reproduction; Wiley Blackwell: Oxford, UK, 2014; pp. 68–78. [Google Scholar]
- Parkinson, T.J. Evaluation of fertility and infertility in natural service bulls. Vet. J. 2004, 168, 215–229. [Google Scholar] [CrossRef] [PubMed]
- Toshimori, K. The spermatozoon. In Knobil and Neill’s Physiology of Reproduction, 4th ed.; Elsevier: New York, NY, USA, 2015; pp. 99–148. [Google Scholar]
- Sergeant, N.; Briand-Amirat, L.; Bencharif, D.; Delehedde, M. The sperm specific protein proAKAP4 as an innovative marker to evaluate sperm quality and fertility. J. Dairy Vet. Sci. 2019, 11, 555803. [Google Scholar] [CrossRef]
- Luconi, M.; Cantini, G.; Baldi, E.; Forti, G. Role of a-kinase anchoring proteins (AKAPs) in reproduction. Front Biosci. 2011, 16, 1315–1330. [Google Scholar] [CrossRef] [Green Version]
- Miki, K.; Willis, W.D.; Brown, P.R.; Goulding, E.H.; Fulcher, K.D.; Eddy, E.M. Targeted disruption of the Akap4 gene causes defects in sperm flagellum and motility. Dev. Biol. 2002, 248, 331–342. [Google Scholar] [CrossRef] [Green Version]
- Moss, S.; Turner, R.; Burkert, K.; Butt, H.; Gerton, G. Conservation and function of a bovine sperm A-kinase anchor protein homologous to mouse AKAP82. Biol. Reprod. 1999, 61, 335–342. [Google Scholar] [CrossRef] [Green Version]
- Fang, X.; Huang, L.L.; Xu, J.; Ma, C.Q.; Chen, Z.H.; Zhang, Z.; Liao, C.H.; Zheng, S.X.; Huang, P.; Xu, W.M.; et al. Proteomics and single-cell RNA analysis of Akap4-knockout mice model confirm indispensable role of Akap4 in spermatogenesis. Dev. Biol. 2019, 454, 118–127. [Google Scholar] [CrossRef]
- Xu, K.; Yang, L.; Zhang, L.; Qi, H. Lack of AKAP3 disrupts integrity of the subcellular structure and proteome of mouse sperm and causes male sterility. Development 2020, 147, dev181057. [Google Scholar] [CrossRef]
- Cui, Z.; Sharma, R.; Agarwal, A. Proteomic analysis of mature and immature ejaculated spermatozoa from fertile men. Asian J. Androl. 2016, 18, 735–746. [Google Scholar] [CrossRef]
- Bastan, I.; Akcay, E. Quality assessment of frozen bull semen with the precursor A-kinase anchor protein 4 biomarker. Andrologia 2021, 53, e14164. [Google Scholar] [CrossRef]
- Blommaert, D.; Sergeant, N.; Delehedde, M.; Jouy, N.; Mitchell, V.; Franck, T.; Donnay, I.; Lejeune, J.; Serteyn, D. Expression, localization, and concentration of A-kinase anchor protein 4 (AKAP4) and its precursor (proAKAP4) in equine semen: Promising marker correlated to the total and progressive motility in thawed spermatozoa. Theriogenology 2019, 131, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Malo, C.; Carracedo, S.; Delehedde, M.; Sergeant, N.; Skidmore, L. Identification of proAKAP4 in Dromedary sperm and their correlation with monthly semen parameters. Reprod. Fertil. 2021, 2, 268–279. [Google Scholar] [CrossRef] [PubMed]
- Jumeau, F.; Sigala, J.; Dossou-Gbete, F.; Frimat, K.; Barbotin, A.L.; Buée, L.; Béhal, H.; Sergeant, N.; Mitchell, V. A-kinase anchor protein 4 precursors (pro-AKAP4) in human sperm. Andrology 2018, 6, 854–859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delehedde, M.; Carracedo, S.; Selleslagh, M.; Eddarkaoui, S.; Amirat-Briand, L.; Sergeant, N. ProAKAP4 polypeptide as a biomarker of sperm functionality and male fertility disorders. Int. J. Gynecol. Reprod. Sci. 2019, 2, 3–19. [Google Scholar]
- Hashemitabar, M.; Sabbagh, S.; Orazizadeh, M.; Ghadiri, A.; Bahmanzadeh, M. A proteomic analysis on human sperm tail: Comparison between normozoospermia and asthenozoospermia. J. Assist. Reprod. Genet. 2015, 32, 853–863. [Google Scholar] [CrossRef] [Green Version]
- Peddinti, D.; Nanduri, B.; Kaya, A.; Feugang, J.M.; Burgess, S.C.; Memili, E. Comprehensive proteomic analysis of bovine spermatozoa of varying fertility rates and identification of biomarkers associated with fertility. BMC Syst. Biol. 2008, 2, 19. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.; Junghare, V.; Hazra, S.; Singh, U.; Sengar, G.S.; Raja, T.; Kumar, S.; Tyagi, S.; Das, A.; Kumar, A.; et al. Database on spermatozoa transcriptogram of catagorised Frieswal crossbred (Holstein Friesian X Sahiwal) bulls. Theriogenology 2019, 129, 130–145. [Google Scholar] [CrossRef]
- Wu, S.; Mipam, T.; Xu, C.; Zhao, W.; Shah, M.A.; Yi, C.; Luo, H.; Cai, X.; Zhong, J. Testis transcriptome profiling identified genes involved in spermatogenic arrest of cattleyak. PLoS ONE 2020, 15, e0229503. [Google Scholar] [CrossRef]
- Fu, Q.; Pan, L.; Huang, D.; Wang, Z.; Hou, Z.; Zhang, M. Proteomic profiles of buffalo spermatozoa and seminal plasma. Theriogenology 2019, 134, 74–82. [Google Scholar] [CrossRef]
- Légaré, C.; Akintayo, A.; Blondin, P.; Calvo, E.; Sullivan, R. Impact of male fertility status on the transcriptome of the bovine epididymis. Mol. Hum. Reprod. 2017, 23, 355–369. [Google Scholar] [CrossRef] [Green Version]
- Utt, M.D. Prediction of bull fertility. Anim. Reprod. Sci. 2016, 169, 37–44. [Google Scholar] [CrossRef]
- Clay, J.S.; McDaniel, B.T. Computing mating bull fertility from DHI nonreturn data. J. Dairy Sci. 2001, 84, 1238–1245. [Google Scholar] [CrossRef]
- Kuhn, M.T.; Hutchison, J.L. Prediction of dairy bull fertility from field data: Use of multiple services and identification and utilization of factors affecting bull fertility. J. Dairy Sci. 2008, 91, 2481–2492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carr, D.W.; Hanlon Newell, A.E. The role of A-kinase anchoringproteins (AKAPs) in regulating sperm function. Soc. Reprod. Fertil. Suppl. 2007, 63, 135–142. [Google Scholar] [PubMed]
- Farrell, P.B.; Presicce, G.A.; Brockett, C.C.; Foote, R.H. Quantification of bull sperm characteristics measured by computer-assisted sperm analysis (CASA) and the relationship to fertility. Theriogenology 1998, 49, 871–879. [Google Scholar] [CrossRef]
- Nagy, Á.; Polichronopoulos, T.; Gáspárdy, A.; Solti, L. Correlation between bull fertility and sperm cell velocity parameters generated by computer-assisted semen analysis. Acta Vet. Hung. 2015, 63, 370–381. [Google Scholar] [CrossRef] [Green Version]
- Puglisi, R.; Pozzi, A.; Foglio, L.; Spanò, M.; Grollino, M.G.; Bongioni, G. The usefulness of combining traditional sperm assessments with in vitro heterospermic insemination to identify bulls of low fertility as estimated in vivo. Anim. Reprod. Sci. 2012, 132, 17–28. [Google Scholar] [CrossRef]
- Campanholi, S.P.; Neto, S.G.; Basso, A.C.; Losano, J.D.D.A.; Siqueira, A.F.P.; Nichi, M.; Assumpção, M.E.O.D.; de Freitas, L.A.; de Paz, C.C.P.; Ferraudo, A.S.; et al. Estimate of in vitro embryo production based on sperm subpopulations in Senepol bulls. Theriogenology 2021, 161, 98–107. [Google Scholar] [CrossRef]
- Brown, P.R.; Miki, K.; Harper, D.B.; Eddy, E.M. A-kinase anchoring protein 4 binding proteins in the fibrous sheath of the sperm flagellum. Biol. Reprod. 2003, 68, 2241–2248. [Google Scholar] [CrossRef] [Green Version]
- Nixon, B.; Bernstein, I.; Café, S.L.; Delehedde, M.; Sergeant, S.; Eamens, A.L.; Lord, T.; Dun, M.D.; De Iuliis, G.N.; Bromfield, E.G. A Kinase Anchor Protein 4 is vulnerable to oxidative adduction in male germ cells. Front. Cell Dev. Biol. 2019, 7, 319. [Google Scholar] [CrossRef] [PubMed]
- Redgrove, K.A.; Nixon, B.; Baker, M.A.; Hetherington, L.; Baker, G.; Liu, D.-Y.; Aitken, R.J. The molecular chaperone HSPA2 plays a key role in regulating the expression of sperm surface receptors that mediate sperm-egg recognition. PLoS ONE 2012, 7, e50851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jagan Mojanarao, G.; Atreja, S.K. Identification of capacitation associated tyrosine phosphoproteins in buffalo and cattle spermatozoa. Anim. Reprod. Sci. 2011, 123, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Yu, H.; Pask, A.; O’Brien, D.; Shaw, G.; Renfree, M. A-kinase anchoring protein 4 has a conserved role in mammalian spermatogenesis. Reproduction 2009, 137, 645–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blommaert, D.; Sergeant, N.; Delehedde, M.; Donnay, I.; Lejeune, J.P.; Franck, T.; Serteyn, D. First results about ProAKAP4 concentration in stallion semen after cryopreservation in two different freezing media. Cryobiology 2021, 102, 133–135. [Google Scholar] [CrossRef] [PubMed]
- Riesco, M.; Anel-Lopez, L.; Neila-Montero, M.; Palacin-Martinez, C.; Montes-Garrido, R.; Alvarez, M.; de Paz, P.; Anel, L. ProAKAP4 as novel molecular marker of sperm quality in Ram: An integrative study in Fresh, cooled and cryopreserved Sperm. Biomolecules 2020, 10, 1046. [Google Scholar] [CrossRef] [PubMed]
- Riesco, M.; Neila-Montero, M.; Palacín-Martínez, C.; Montes-Garrido, R.; Álvarez, R.; Boixo, J.C.; De Paz, P.; Anel, L.; Anel-López, L. Establishment of innovative biomarkers to optimize cooling and cryopreservation protocols in ram sperm. Reprod. Domest. Anim. 2022, 57, 101. [Google Scholar]
- Kowalczyk, A.; Gałeska, E.; Bubel, A. The concentration of ProAKAP4 and other indicators of cryopotential of spermatozoa cryopreserved in extender with Holothuroidea extract addition. Animals 2022, 12, 521. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| proAKAP4 ng/10 M spz | PR (%) | VAP (μm/s) | VSL (μm/s) | VCL (μm/s) | BCF (Hz) | LIN (%) | STR (%) | |
|---|---|---|---|---|---|---|---|---|
| n | 40 | 40 | 40 | 40 | 40 | 40 | 40 | 40 |
| Min. | 19.96 | 16.00 | 77.65 | 53.78 | 142.5 | 29.80 | 32.83 | 69.00 |
| 25% | 30.51 | 34.46 | 92.24 | 77.57 | 162.6 | 34.73 | 42.30 | 80.00 |
| Median | 38.62 | 37.92 | 98.94 | 84.06 | 175.1 | 37.36 | 44.83 | 83.00 |
| 75% | 49.23 | 44.38 | 105.0 | 88.37 | 187.8 | 38.60 | 48.46 | 85.00 |
| Max | 96.95 | 53.33 | 131.5 | 103.5 | 251.7 | 40.80 | 53.17 | 88.00 |
| Mean | 41.07 | 38.67 | 100.9 | 83.20 | 181.6 | 36.28 | 44.87 | 82.40 |
| SD | 15.42 | 8.554 | 12.45 | 10.21 | 26.77 | 3.048 | 4.516 | 4.088 |
| Lower 95% CI | 36.14 | 35.93 | 96.96 | 79.93 | 173.0 | 35.30 | 43.42 | 81.09 |
| Upper 95% CI | 46.00 | 41.40 | 104.9 | 86.46 | 190.2 | 37.25 | 46.31 | 83.71 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dordas-Perpinyà, M.; Sergeant, N.; Ruelle, I.; Bruyas, J.-F.; Charreaux, F.; Michaud, S.; Carracedo, S.; Catalán, J.; Miró, J.; Delehedde, M.; et al. ProAKAP4 Semen Concentrations as a Valuable Marker Protein of Post-Thawed Semen Quality and Bull Fertility: A Retrospective Study. Vet. Sci. 2022, 9, 224. https://doi.org/10.3390/vetsci9050224
Dordas-Perpinyà M, Sergeant N, Ruelle I, Bruyas J-F, Charreaux F, Michaud S, Carracedo S, Catalán J, Miró J, Delehedde M, et al. ProAKAP4 Semen Concentrations as a Valuable Marker Protein of Post-Thawed Semen Quality and Bull Fertility: A Retrospective Study. Veterinary Sciences. 2022; 9(5):224. https://doi.org/10.3390/vetsci9050224
Chicago/Turabian StyleDordas-Perpinyà, Marta, Nicolas Sergeant, Isabelle Ruelle, Jean-François Bruyas, Frédéric Charreaux, Sandrine Michaud, Sara Carracedo, Jaime Catalán, Jordi Miró, Maryse Delehedde, and et al. 2022. "ProAKAP4 Semen Concentrations as a Valuable Marker Protein of Post-Thawed Semen Quality and Bull Fertility: A Retrospective Study" Veterinary Sciences 9, no. 5: 224. https://doi.org/10.3390/vetsci9050224
APA StyleDordas-Perpinyà, M., Sergeant, N., Ruelle, I., Bruyas, J. -F., Charreaux, F., Michaud, S., Carracedo, S., Catalán, J., Miró, J., Delehedde, M., & Briand-Amirat, L. (2022). ProAKAP4 Semen Concentrations as a Valuable Marker Protein of Post-Thawed Semen Quality and Bull Fertility: A Retrospective Study. Veterinary Sciences, 9(5), 224. https://doi.org/10.3390/vetsci9050224