1.2. Description of Stages
The descriptions are illustrated from caiman embryos (
Figure 1,
Figure 2,
Figure 3,
Figure 4,
Figure 5,
Figure 6 and
Figure 7); only
Figure 8 is from an alligator. We chose to start our study with stage 17, although several elements (endocardial cushions, parts of the muscular interventricular septum) already appear earlier. In St 17, the elements start to make up a comprehensible combination leading to complete septation.
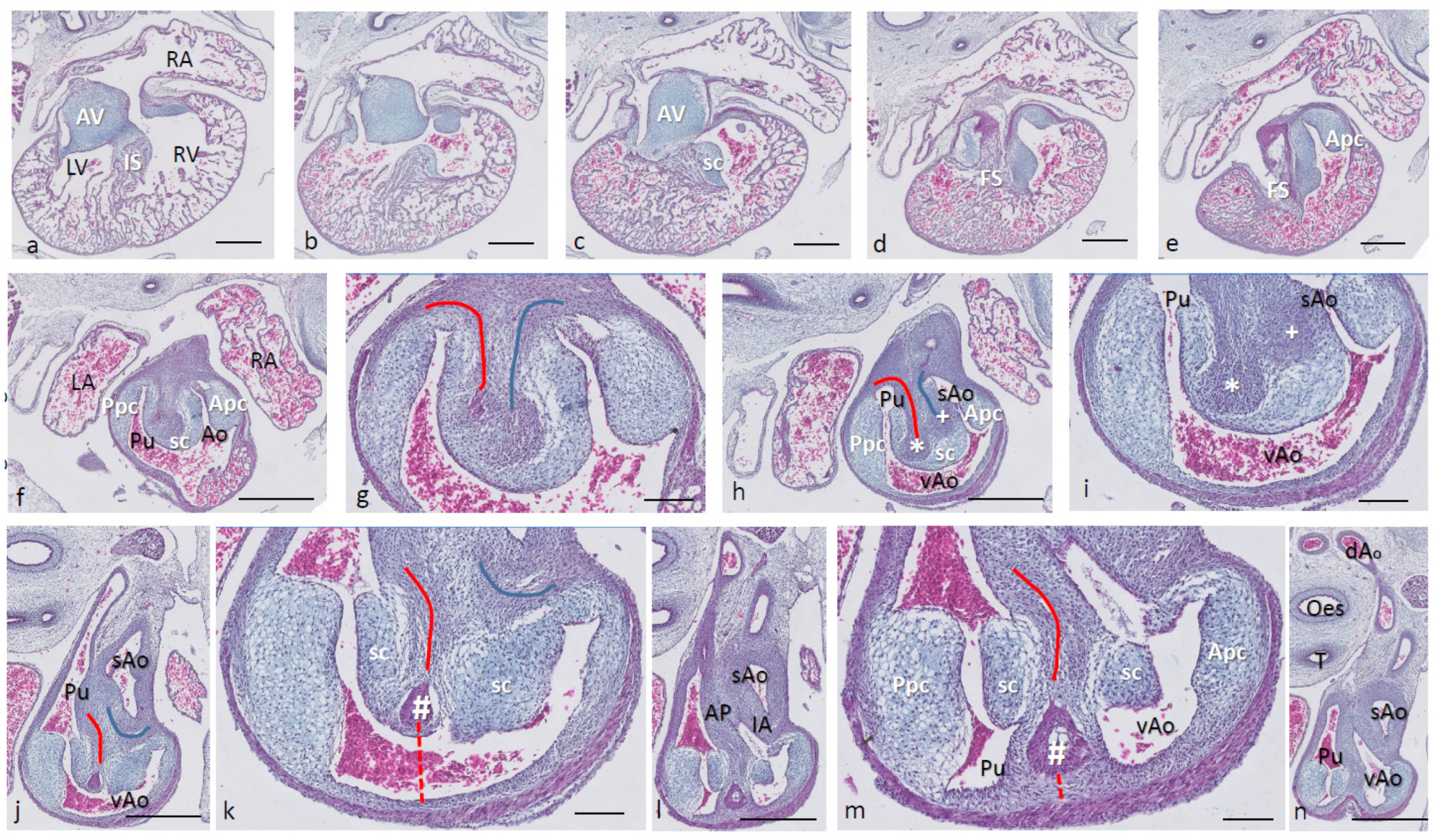
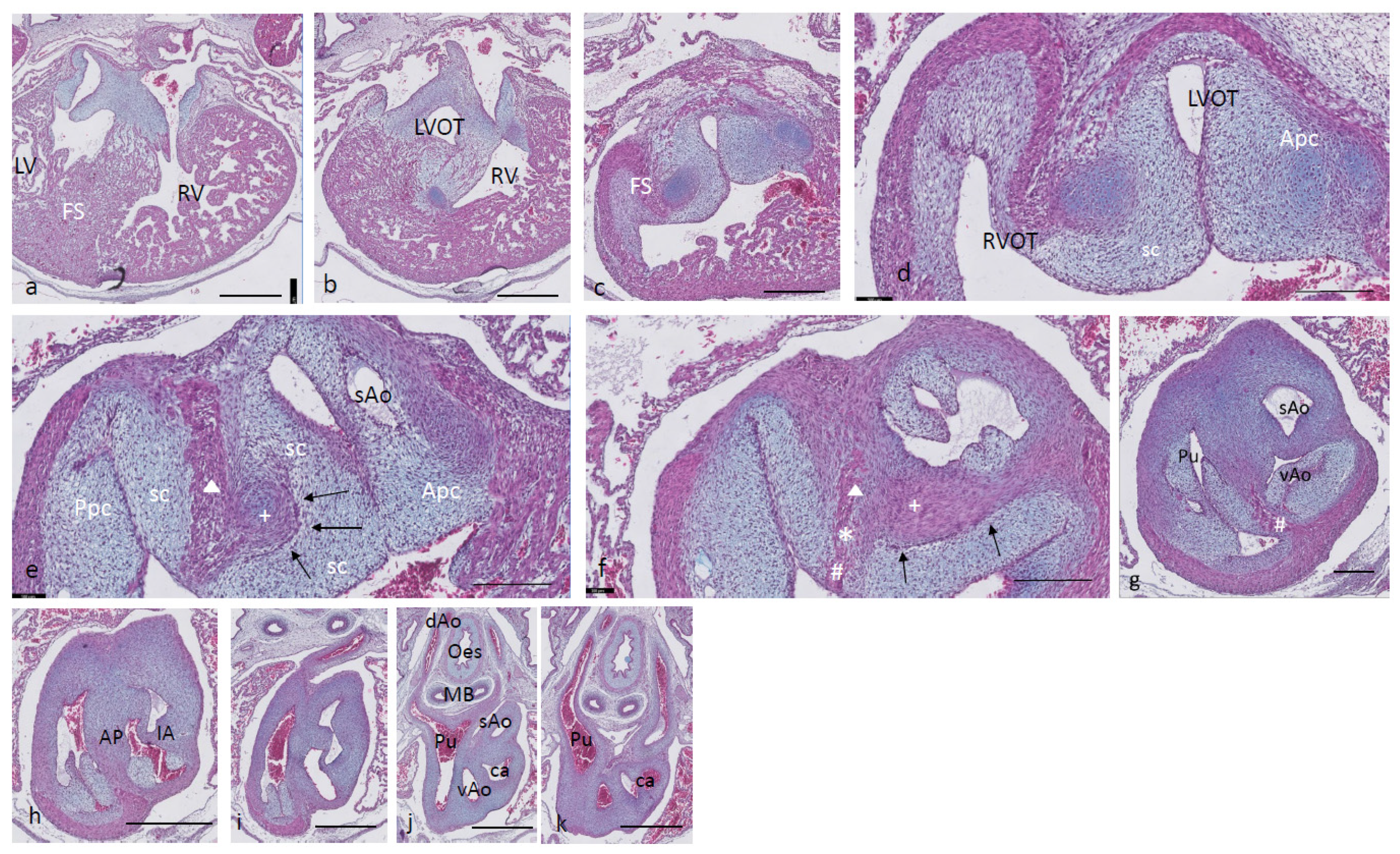
Ferguson stage 17. The right and left ventricles are dorsally intersected by the inlet septum (
Figure 1a), but otherwise not separated, as the interventricular communication presents itself clearly (
Figure 1b). The ventricular inlet septum is fused with the large central AV cushion complex, while the folding septum [
20] can hardly be discerned (
Figure 1a–c). We have defined the folding septum [
20] as a part of the muscular interventricular septum particularly located between the right and left ventricular outflow tracts. Slightly more downstream, the septal outflow tract cushion (sc) becomes apparent, together with the flanking aortic and pulmonary parietal outflow tract cushions (Apc and Ppc) (
Figure 1d–f). In the septal cushion, two streaks of condensed mesenchymal cells are evident, one on the pulmonary side (red curve in
Figure 1g–i), and the other on the aortic side (blue curve). At their tips (* and + in
Figure 1h,i), both contain a histologically very dense cluster of cells. In a recent study [
20], we showed the presence of neural crest cells in this dense cluster by the expression of AP2alpha, a known marker of NC. The pulmonary streak or aorto-pulmonary (AP) septum might contain left-sided NC-derived cells, while the aortic streak or interaortic (IA) septum probably contains right-sided NC cells, as it will be discussed later. The curved lumen of the outflow tract (
Figure 1h,i) can be divided into the pulmonary outflow tract (from the right ventricle), the visceral aorta (vAo, also from the right ventricle) and the systemic aorta (sAo, left ventricle). Separation of the outflow tracts (
Figure 1j) takes place by 1. the AP septum including a histologically distinguishable ventral myocardial spur (# in
Figure 1j–n) located between the pulmonary trunk and the aortas, and 2. the IA septum between sAo and vAo. In the thorax, no further branches will be found emanating from the vAo. The sAo, however, branches further in both carotid arteries (not depicted here).
Figure 1.
Ferguson stage 17. HE Alcian blue-stained serially sectioned embryo. Figure (a–n) from AV canal to pharyngeal arch arteries. Generally, the endocardial cushions stand out because of the Alcian blue staining. Figure (a), myocardium is spongious with a thin compact outer layer. The inlet septum is the most conspicuous part of the interventricular septum located between the left- and right-sided parts of the undivided common ventricle. Figure (b,c), the outflow tract septal cushion becomes visible. Figure (d,e), the folding septum only becomes clear more distally. Figure (f), pulmonary and aortic parietal OFT cushions become apparent, flanking the pulmonary channel and the yet common aortic channel. Figure (g), magnified Figure (f, in the large centrally located septal cushion, two streams of condensed mesenchyme, on the pulmonary side (in red) and the aortic side (in blue). The septal cushion becomes subdivided over the main arterial channels. Figure (h,i), both streams end separately in a bulbous structure as the basis for the AP septum (*) and IA septum (+). Figure (j), magnified in Figure (k), the AP stream meets the ventral myocardial spur (#). Figure (l), magnified in (m), the myocardial spur is part of the ventral myocardium indicated by the red dotted line. At this level, the AP septum is completed. Figure (n), pharyngeal arch arteries are separated, and the connection to the dorsal aorta is present. Key to the symbols # ventral myocardium, part of AP septum; * (presumably) left-sided neural crest cells, part of AP septum; + (presumably) right-sided neural crest, part of IA septum; Δ dorsal myocardium, part of AP septum. Magnification Figure (a–e,f,h,j,l,n), bar 500 µm; (g,i,k,m), 200 µm.
Figure 1.
Ferguson stage 17. HE Alcian blue-stained serially sectioned embryo. Figure (a–n) from AV canal to pharyngeal arch arteries. Generally, the endocardial cushions stand out because of the Alcian blue staining. Figure (a), myocardium is spongious with a thin compact outer layer. The inlet septum is the most conspicuous part of the interventricular septum located between the left- and right-sided parts of the undivided common ventricle. Figure (b,c), the outflow tract septal cushion becomes visible. Figure (d,e), the folding septum only becomes clear more distally. Figure (f), pulmonary and aortic parietal OFT cushions become apparent, flanking the pulmonary channel and the yet common aortic channel. Figure (g), magnified Figure (f, in the large centrally located septal cushion, two streams of condensed mesenchyme, on the pulmonary side (in red) and the aortic side (in blue). The septal cushion becomes subdivided over the main arterial channels. Figure (h,i), both streams end separately in a bulbous structure as the basis for the AP septum (*) and IA septum (+). Figure (j), magnified in Figure (k), the AP stream meets the ventral myocardial spur (#). Figure (l), magnified in (m), the myocardial spur is part of the ventral myocardium indicated by the red dotted line. At this level, the AP septum is completed. Figure (n), pharyngeal arch arteries are separated, and the connection to the dorsal aorta is present. Key to the symbols # ventral myocardium, part of AP septum; * (presumably) left-sided neural crest cells, part of AP septum; + (presumably) right-sided neural crest, part of IA septum; Δ dorsal myocardium, part of AP septum. Magnification Figure (a–e,f,h,j,l,n), bar 500 µm; (g,i,k,m), 200 µm.
![Jcdd 08 00132 g001]()
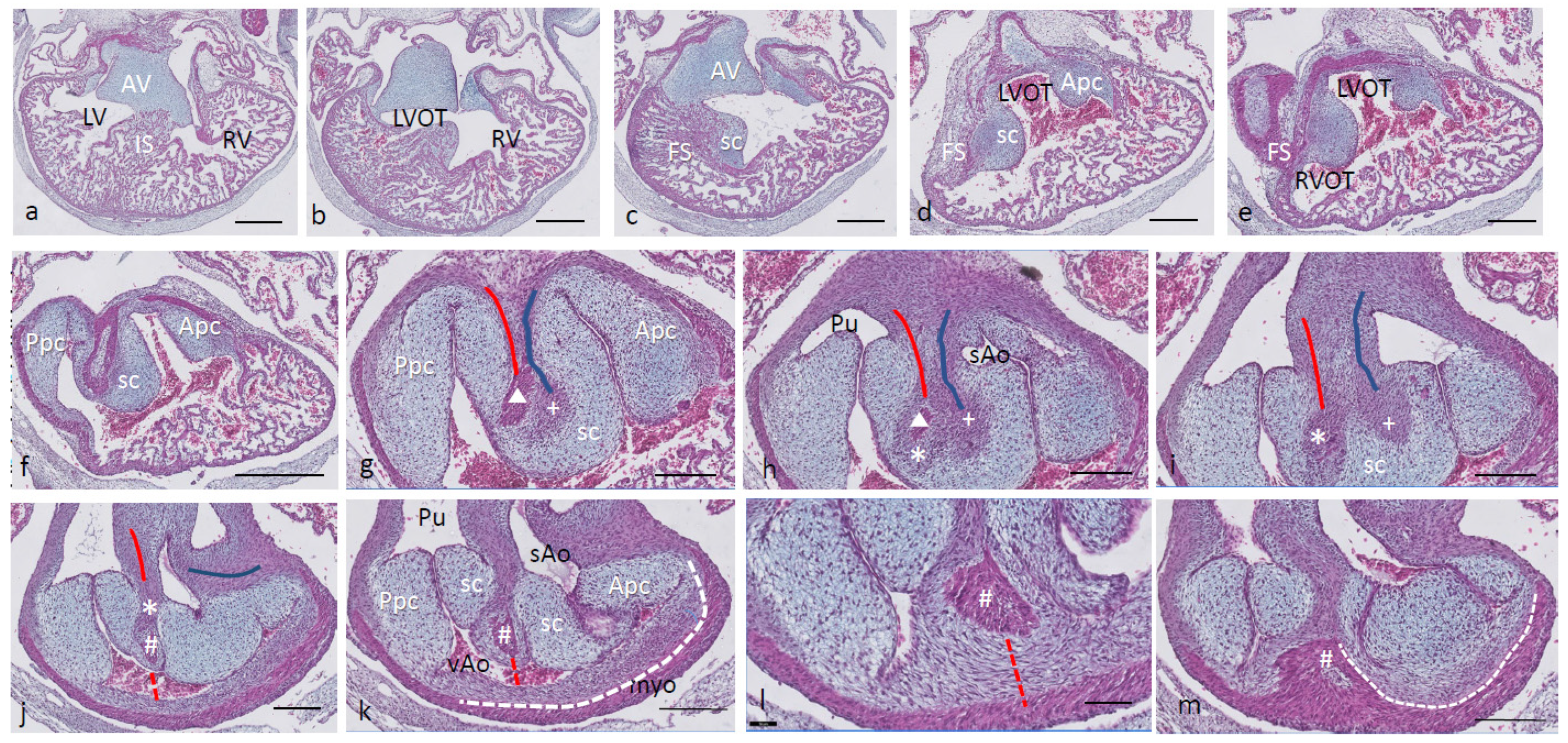
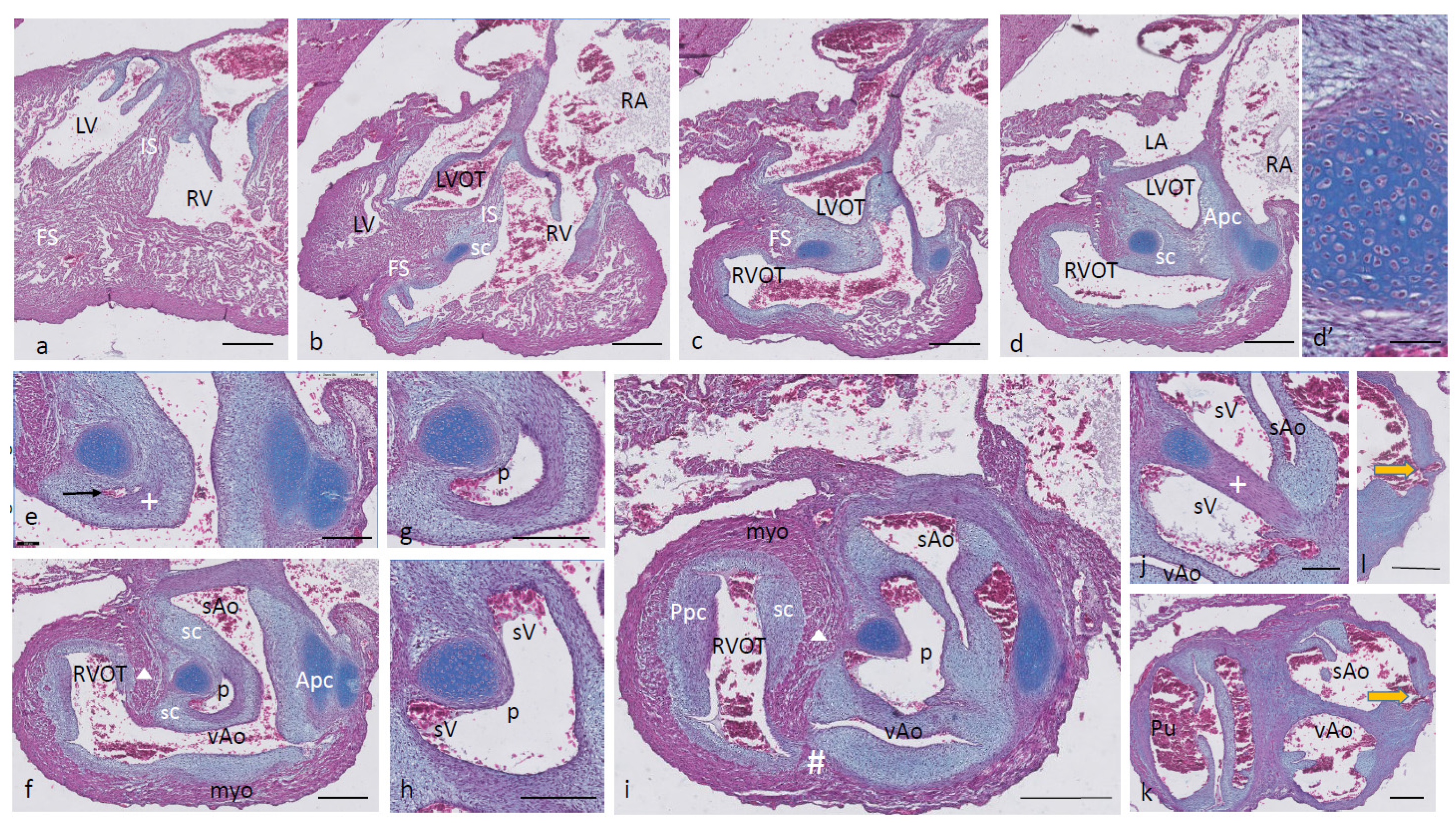
Ferguson stage 19. The interventricular situation (
Figure 2a,b) has not advanced very much compared to stage 17 as the interventricular communication is still open and the folding septum (FS) remains inconspicuous (
Figure 2c). The septal outflow tract cushion (sc) acquires a dense core (
Figure 2c) preceding the differentiation of cartilage in the next stage. The interconnected lumina of the LVOT and RVOT are easily recognized (
Figure 2d,e), interrupted by the large septal cushion (sc). The root of the Pu seems to be hconnected sideways (
Figure 2d–f) to the ventricular segment as a result of folding of this part of the interventricular septum. The septal cushion is located at the top of the folding septum (
Figure 2c,d) that becomes more clear in the next stage. In the as yet common outflow tract, the three arteries become separated by the AP septum and the IA septum. In the AP septum, the region of the originally dense cluster of cells (see
Figure 2h *) surrounds a dorsal spur of the myocardium (
in
Figure 2g,h), being the cranial tip of the folding septum. This dorsal myocardial spur has not yet fused with the ventral myocardial spur (#
Figure 2j–m and dashed red line (Figure l) to show the continuation with the ventral wall) as there is a “window”, free from the myocardium in between (* in
Figure 2h–j). AP separation and IA separation are seemingly spatially independent of each other (
Figure 2i,j). The septal cushion is distally completely bisected by the AP septum (
Figure 2k). The right-sided IA septum deviates between both aortas with a dense cell cluster at its tip (+ in
Figure 2h,i).
The transition of the cardiac tube to the arterial vascular wall deserves special attention. It is evident that the arterial wall extends inside the cardiac tube over a considerable distance estimated to be about 200 µm on the pulmonary side and even about 400 µm on the aortic side (white dashed line in
Figure 2k). Downstream, this appearance is even more substantiated (
Figure 2m), where nearly the complete ventral wall of the vAo seems to consist of a double layer of the vessel wall and myocardium (white dashed line) with hardly a thin endocardial cushion tissue. This continues to an estimated depth of 350–400 µm into the myocardial tube. As a consequence, we are dealing with two concentric tubes, a myocardial circumference surrounding an arterial wall.
Figure 2.
Ferguson stage 19. HE Alcian blue-stained serially sectioned embryo. Figure (a–m) from AV canal to OFT. Figure (a), ventricular myocardial inlet septum attached to the central AV cushions between the left and right ventricles. The myocardium is spongy but for the thin outer compact layer. Figure (b), more ventrally, the interventricular communication is visible, while the left ventricular outflow tract becomes apparent. Figure (c), muscular folding septum appears as continuation of the inlet septum. The septal OFT cushion is located at the tip of the folding septum. Figure (d), septal cushion with Alcian blue-stained condensed mesenchyme. The aortic parietal cushion appears on the other side of the LVOT. Figure (e), the septal cushion is located between both outflow tracts, while the folding septum has shifted to the right side. Figure (f), in the RVOT the pulmonary parietal cushion is present, and the folding septum including the septal cushion takes up a dorsal position. Figure (g), the most cranial remnant of the muscular folding septum is indicated (Δ). The adjacent condensed mesenchyme (+) belonging to the interaortic septum has appeared in the septal cushion. Figure (h), a second element of condensed mesenchyme (*) appears ventral to the disappearing myocardium (Δ). Figure (i), two streams of condensed mesenchyme as part of the aorto-pulmonary septum (*) and the interaortic septum (+) separate the septal cushion into three subcushions. Figure (j), more distally, only the AP septum continues and receives the myocardium (#) from the ventral wall indicated by the red dotted line. Figure (k), the condensed mesenchyme of the AP septum is not present anymore, but the myocardial component has enlarged (#). The pulmonary trunk, the visceral aorta and the systemic aorta are indicated, as are the endocardial cushion components. Only the visceral aorta is still encased in the myocardium, but note that the arterial wall of the visceral aorta continues within the myocardial tube (dashed line). Figure (l), the visceral and systemic aortas are separated at this level by the AP septum (red line). Figure (m), here, the AP septum is myocardialized (#). Key to the symbols # ventral myocardium, part of AP septum; * (presumably) left-sided neural crest cells, part of AP septum; + (presumably) right-sided neural crest, part of IA septum; Δ dorsal myocardium, part of AP septum. Magnification Figure (a–f), bar 500 µm; (g–m), bar 200 µm; (g–m), 200 µm; (l), 100 µm.
Figure 2.
Ferguson stage 19. HE Alcian blue-stained serially sectioned embryo. Figure (a–m) from AV canal to OFT. Figure (a), ventricular myocardial inlet septum attached to the central AV cushions between the left and right ventricles. The myocardium is spongy but for the thin outer compact layer. Figure (b), more ventrally, the interventricular communication is visible, while the left ventricular outflow tract becomes apparent. Figure (c), muscular folding septum appears as continuation of the inlet septum. The septal OFT cushion is located at the tip of the folding septum. Figure (d), septal cushion with Alcian blue-stained condensed mesenchyme. The aortic parietal cushion appears on the other side of the LVOT. Figure (e), the septal cushion is located between both outflow tracts, while the folding septum has shifted to the right side. Figure (f), in the RVOT the pulmonary parietal cushion is present, and the folding septum including the septal cushion takes up a dorsal position. Figure (g), the most cranial remnant of the muscular folding septum is indicated (Δ). The adjacent condensed mesenchyme (+) belonging to the interaortic septum has appeared in the septal cushion. Figure (h), a second element of condensed mesenchyme (*) appears ventral to the disappearing myocardium (Δ). Figure (i), two streams of condensed mesenchyme as part of the aorto-pulmonary septum (*) and the interaortic septum (+) separate the septal cushion into three subcushions. Figure (j), more distally, only the AP septum continues and receives the myocardium (#) from the ventral wall indicated by the red dotted line. Figure (k), the condensed mesenchyme of the AP septum is not present anymore, but the myocardial component has enlarged (#). The pulmonary trunk, the visceral aorta and the systemic aorta are indicated, as are the endocardial cushion components. Only the visceral aorta is still encased in the myocardium, but note that the arterial wall of the visceral aorta continues within the myocardial tube (dashed line). Figure (l), the visceral and systemic aortas are separated at this level by the AP septum (red line). Figure (m), here, the AP septum is myocardialized (#). Key to the symbols # ventral myocardium, part of AP septum; * (presumably) left-sided neural crest cells, part of AP septum; + (presumably) right-sided neural crest, part of IA septum; Δ dorsal myocardium, part of AP septum. Magnification Figure (a–f), bar 500 µm; (g–m), bar 200 µm; (g–m), 200 µm; (l), 100 µm.
![Jcdd 08 00132 g002]()
Ferguson stage 20. The interventricular communication between the LVOT and RV is almost closed (
Figure 3a). The dense core in the septal and in the aortic parietal cushions starts to differentiate into cartilage, as shown by intense Alcian blue staining (
Figure 3a–c), although more downstream, this differentiation is not apparent yet (
Figure 3d,e). Here, the IA septum consists of densely packed right-sided neural crest cells, as described in the earlier stages (+ in
Figure 3d). The AP septum now contains a clear core of the myocardium (Δ in
Figure 3d–f) as part of the incurving dorsal wall of the myocardial tube. Where the septal cushion has connected with the ventral wall, the ventral myocardial spur (# in
Figure 3f–h) is also present. The two spurs do not contact each other (* depicts the mesenchymal “window” between the myocardial spurs indicated by Δ and # in
Figure 3g). The AP and IA septa deviate (
Figure 3h,i) to continue outside the heart as the mesenchymal vessel walls of the respective arteries (
Figure 3j–l). The common stem of the carotid arteries (ca) branches from the systemic aorta (
Figure 3l). Two sinuses of Valsalva (white arrow) are present in the OFT cushions of the systemic aorta and the pulmonary trunk (
Figure 3h,i) as the first sign of the forming semilunar valve leaflets. The situation in the visceral aorta is slightly different as in the parietal cushion, a sinus of Valsalva is still lacking, but in the septal cushion, a very narrow slit is seen, representing the sinus of Valsalva that is connected to the main lumen of the vAo, probably being the first sign of the origin of the foramen of Panizza (see description in the next stage, stage 21).
Figure 3.
Ferguson stage 20, Figure (a–l) from AV canal to OFT. Figure (a), both in the septal and aortic parietal cushions, cartilage is forming (dark blue) which is less evident more distally (Figure (b)). Figure (c), the septal cushion containing condensed mesenchyme has a central position. Figure (d,e), the septal cushion becomes subdivided by both prongs of condensed mesenchyme, belonging to the IA septum (+) and AP septum (*), where the remnant of the folding septum is indicated (Δ). Figure (f), the condensed mesenchyme of the IA septum bridges the gap to the aortic parietal cushion whereby the systemic and visceral aortas become separated at this level. The AP septum now contains 3 elements. Enlargement in Figure (g), condensed mesenchyme (*), flanked by the myocardium of the folding septum (Δ) and the ventral myocardium (#). Figure (h), both IA and AP septa are fully developed at this level. The arrows indicate the sinus of Valsalva in the systemic and visceral aortas. Figure (i), the arrow indicates a sinus of Valsalva in the pulmonary trunk. Figure (j–l) illustrate the branching of the arterial tree, including the pulmonary arteries, and the carotid arteries from the systemic aorta. The visceral aorta does not branch further. Key to the symbols # ventral myocardium, part of AP septum; * (presumably) left-sided neural crest cells, part of AP septum; + (presumably) right-sided neural crest, part of IA septum; Δ dorsal myocardium, part of AP septum. Magnification Figure (a–f,h–l), 500 µm; (g), 100 µm.
Figure 3.
Ferguson stage 20, Figure (a–l) from AV canal to OFT. Figure (a), both in the septal and aortic parietal cushions, cartilage is forming (dark blue) which is less evident more distally (Figure (b)). Figure (c), the septal cushion containing condensed mesenchyme has a central position. Figure (d,e), the septal cushion becomes subdivided by both prongs of condensed mesenchyme, belonging to the IA septum (+) and AP septum (*), where the remnant of the folding septum is indicated (Δ). Figure (f), the condensed mesenchyme of the IA septum bridges the gap to the aortic parietal cushion whereby the systemic and visceral aortas become separated at this level. The AP septum now contains 3 elements. Enlargement in Figure (g), condensed mesenchyme (*), flanked by the myocardium of the folding septum (Δ) and the ventral myocardium (#). Figure (h), both IA and AP septa are fully developed at this level. The arrows indicate the sinus of Valsalva in the systemic and visceral aortas. Figure (i), the arrow indicates a sinus of Valsalva in the pulmonary trunk. Figure (j–l) illustrate the branching of the arterial tree, including the pulmonary arteries, and the carotid arteries from the systemic aorta. The visceral aorta does not branch further. Key to the symbols # ventral myocardium, part of AP septum; * (presumably) left-sided neural crest cells, part of AP septum; + (presumably) right-sided neural crest, part of IA septum; Δ dorsal myocardium, part of AP septum. Magnification Figure (a–f,h–l), 500 µm; (g), 100 µm.
![Jcdd 08 00132 g003]()
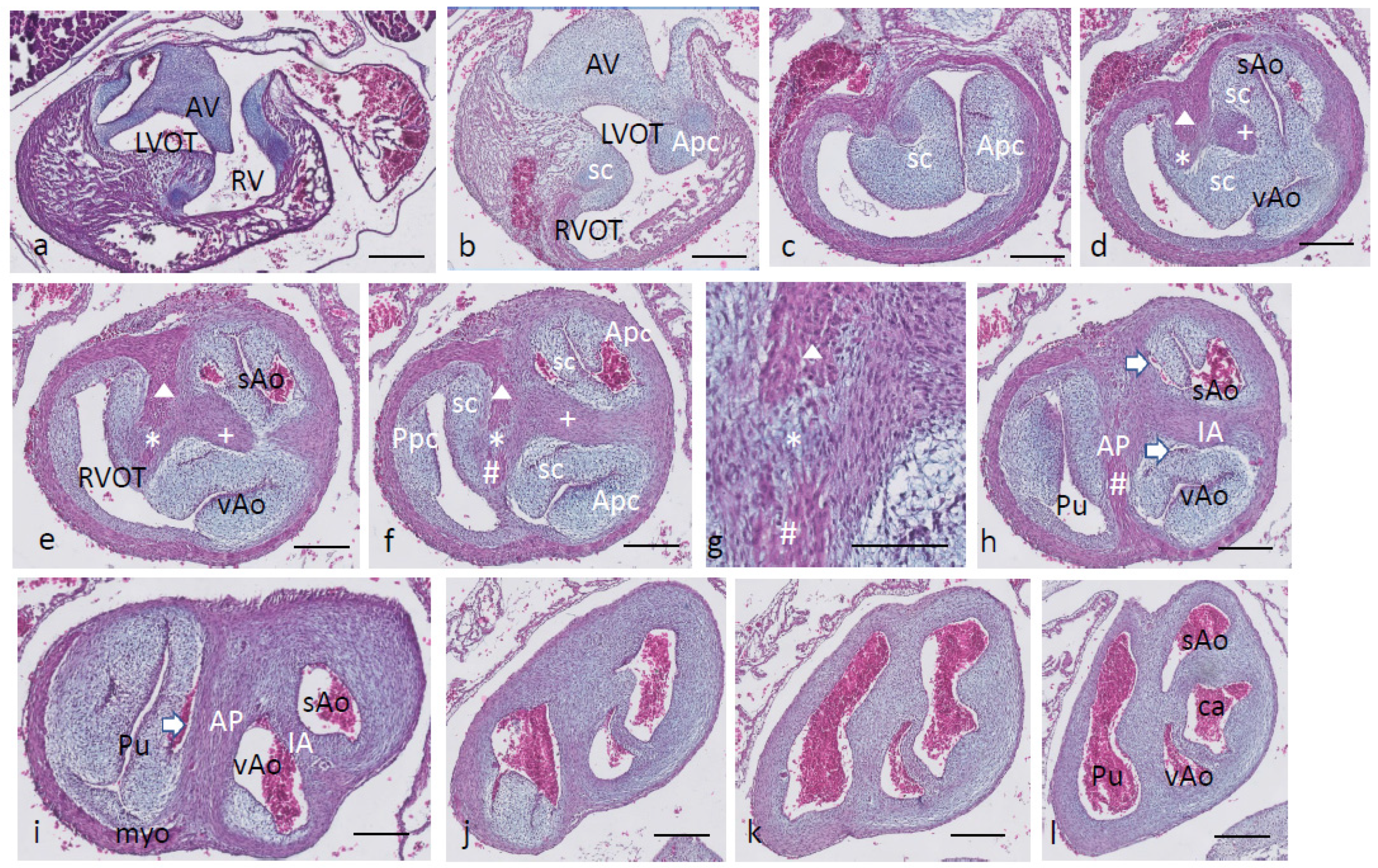
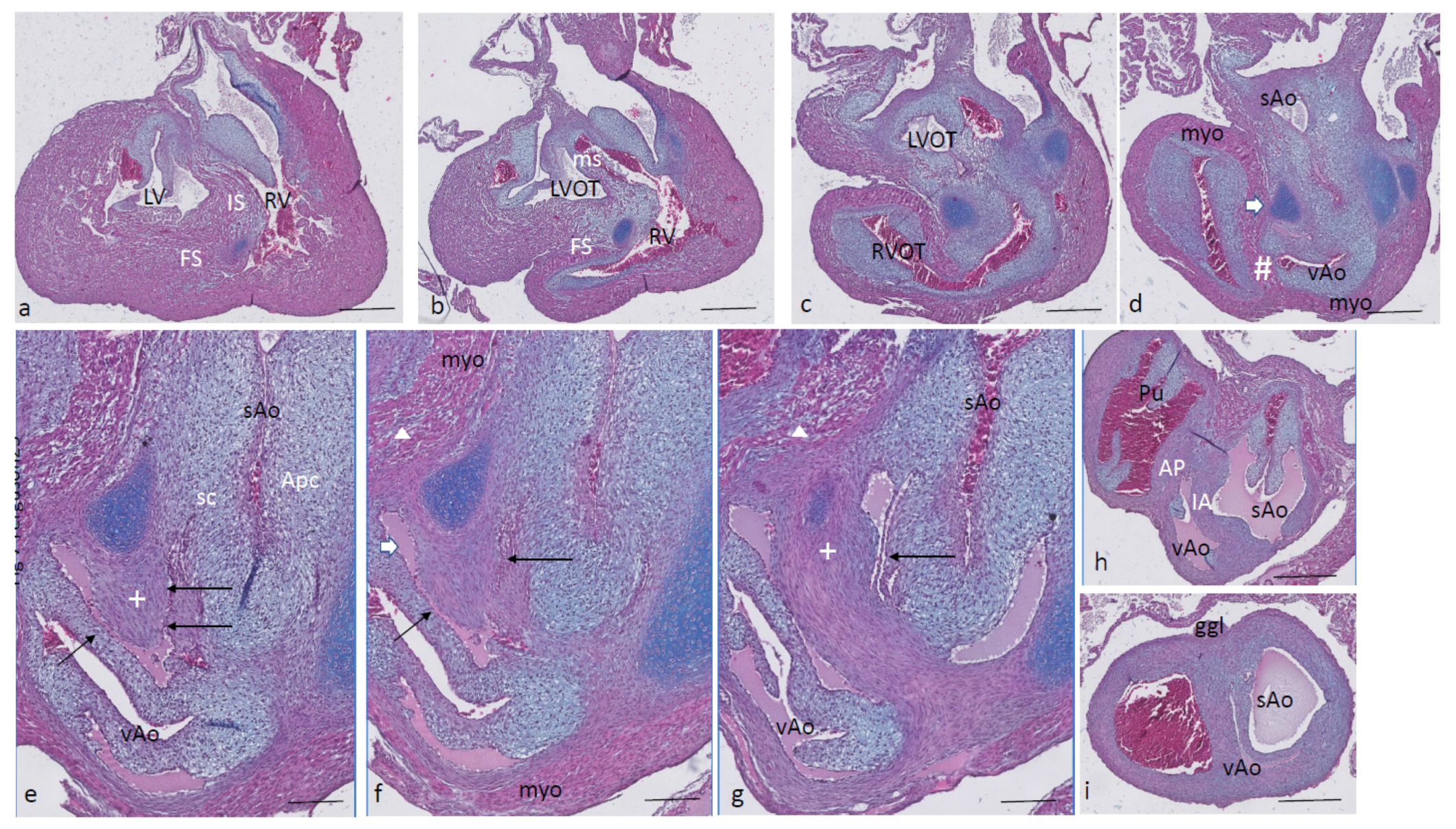
Ferguson stage 21. The ventricular inflow septum (IS) is seen as the continuation of the folding septum (
Figure 4a,b), beyond the topping cartilage of the septal cushion (
Figure 4b–e). Another smaller piece of cartilage is found in the aortic parietal cushion (
Figure 4c,d). In the outflow tract, the pulmonary side becomes separated from the aortic side by the large septal cushion, containing the elements of the AP septum (* and Δ in
Figure 4e–h) as well as the IA septum (+ in
Figure 4f–h). Here, an interesting feature becomes apparent, known as the foramen of Panizza (FOP, black arrows in
Figure 4f,g), connecting the two septal sinuses of Valsalva of the sAo (white arrow) and vAo. Actually, the FOP in this stadium is a very narrow channel, about 100 µm high in the cranio-caudal direction and about 200 µm between both sinuses. The channel surrounds the advancing tip of the IA (+ in
Figure 4f,g) that, at this level, is completed in the arterial direction only (
Figure 4h,i). Further downstream, the myocardial tube is replaced by the arterial vessel walls in the concentric manner described above. The stem of the carotid arteries branches from the sAo (
Figure 4j,k).
Figure 4.
Ferguson stage 21, from AV canal to the arterial tree. Figure (a), the ventricular septum shows its two parts: the inlet septum attached to the central AV cushions and the more ventro-distally located folding septum. Figure (b,c), the septal cushion already showing cartilage differentiation (dark blue) located on the tip of the folding septum. Figure (d,e), the right ventricular outflow tract rotates to the right, displacing the folding septum (Δ) to the dorsal wall. Figure (f,g), the AP (*) and IA (+) streams of condensed mesenchyme are visible. The sinus of Valsalva (white arrow) and the tunnel of the foramen of Panizza (black arrows) are indicated. Figure (h–j), more distally, the AP and IA septa become completed by fusion of the septal cushion with the parietal cushion. Figure (k), branching of the arterial tree. Key to the symbols # ventral myocardium, part of AP septum; * (presumably) left-sided neural crest cells, part of AP septum; + (presumably) right-sided neural crest, part of IA septum; Δ dorsal myocardium, part of AP septum. Magnification Figure (a–e,h–k), bar 500 µm; (f,g), bar 200 µm.
Figure 4.
Ferguson stage 21, from AV canal to the arterial tree. Figure (a), the ventricular septum shows its two parts: the inlet septum attached to the central AV cushions and the more ventro-distally located folding septum. Figure (b,c), the septal cushion already showing cartilage differentiation (dark blue) located on the tip of the folding septum. Figure (d,e), the right ventricular outflow tract rotates to the right, displacing the folding septum (Δ) to the dorsal wall. Figure (f,g), the AP (*) and IA (+) streams of condensed mesenchyme are visible. The sinus of Valsalva (white arrow) and the tunnel of the foramen of Panizza (black arrows) are indicated. Figure (h–j), more distally, the AP and IA septa become completed by fusion of the septal cushion with the parietal cushion. Figure (k), branching of the arterial tree. Key to the symbols # ventral myocardium, part of AP septum; * (presumably) left-sided neural crest cells, part of AP septum; + (presumably) right-sided neural crest, part of IA septum; Δ dorsal myocardium, part of AP septum. Magnification Figure (a–e,h–k), bar 500 µm; (f,g), bar 200 µm.
![Jcdd 08 00132 g004]()
Ferguson stage 22. Most of the features described for stage 21 apply to this stage. The folding septum is more advanced (
Figure 5a). The condensed mesenchyme in the septal cushion is further differentiated into cartilage (
Figure 5b–d), even reaching the level of the FOP (
Figure 5d,e, black arrows). The FOP is seen as a very narrow channel rounding the cartilaginous tip at the base of the IA (
Figure 5d,e). The condensed mesenchyme in the aortic parietal cushion is advanced and now contains two cartilaginous centers (
Figure 5c), one in the continuation of the aortic vessel wall, and the second one in the cushion mesenchyme. The ventral and dorsal myocardial spurs (# and Δ in
Figure 5e) have joined with no mesenchymal window in between anymore, implying that a small myocardial bridge has been established in the AP septum. The IA septum does not acquire a myocardial component in contrast to the AP septum (
Figure 5e–g). The branching of the main arterial stems is demonstrated in
Figure 5h–i. The pulmonary trunk is divided into the sixth left and right pulmonary arch arteries (PAA6,
Figure 5h), and the vAo, being the left fourth PAA, does not branch further, whereas the stem of the carotid arch arteries splits from the right fourth PAA, being the sAo (
Figure 5h–i).
Figure 5.
Ferguson stage 22, Figure (a–i), from AV canals to the arterial tree. Figure (a,b), septal cushion with cartilage between left and right ventricles. Figure (c), cartilage differentiation in both the septal and aortic parietal cushions. Figure (d), enlargement of Figure c, the septal cartilage related to the IA septum is located on the left aortic side of the folding septum. Figure (e,f), the AP (*), (Δ), (#) and the IA (+) septa are completely visible, containing their various elements. The tunnel of the foramen of Panizza is present (black arrows). Figure (g–k) demonstrate the branching pattern of the arterial tree. Key to the symbols # ventral myocardium, part of AP septum; * (presumably) left-sided neural crest cells, part of AP septum; + (presumably) right-sided neural crest, part of IA septum; Δ dorsal myocardium, part of AP septum. Magnification Figure (a–c,h–k), bar 500 µm; (d–g), 200 µm.
Figure 5.
Ferguson stage 22, Figure (a–i), from AV canals to the arterial tree. Figure (a,b), septal cushion with cartilage between left and right ventricles. Figure (c), cartilage differentiation in both the septal and aortic parietal cushions. Figure (d), enlargement of Figure c, the septal cartilage related to the IA septum is located on the left aortic side of the folding septum. Figure (e,f), the AP (*), (Δ), (#) and the IA (+) septa are completely visible, containing their various elements. The tunnel of the foramen of Panizza is present (black arrows). Figure (g–k) demonstrate the branching pattern of the arterial tree. Key to the symbols # ventral myocardium, part of AP septum; * (presumably) left-sided neural crest cells, part of AP septum; + (presumably) right-sided neural crest, part of IA septum; Δ dorsal myocardium, part of AP septum. Magnification Figure (a–c,h–k), bar 500 µm; (d–g), 200 µm.
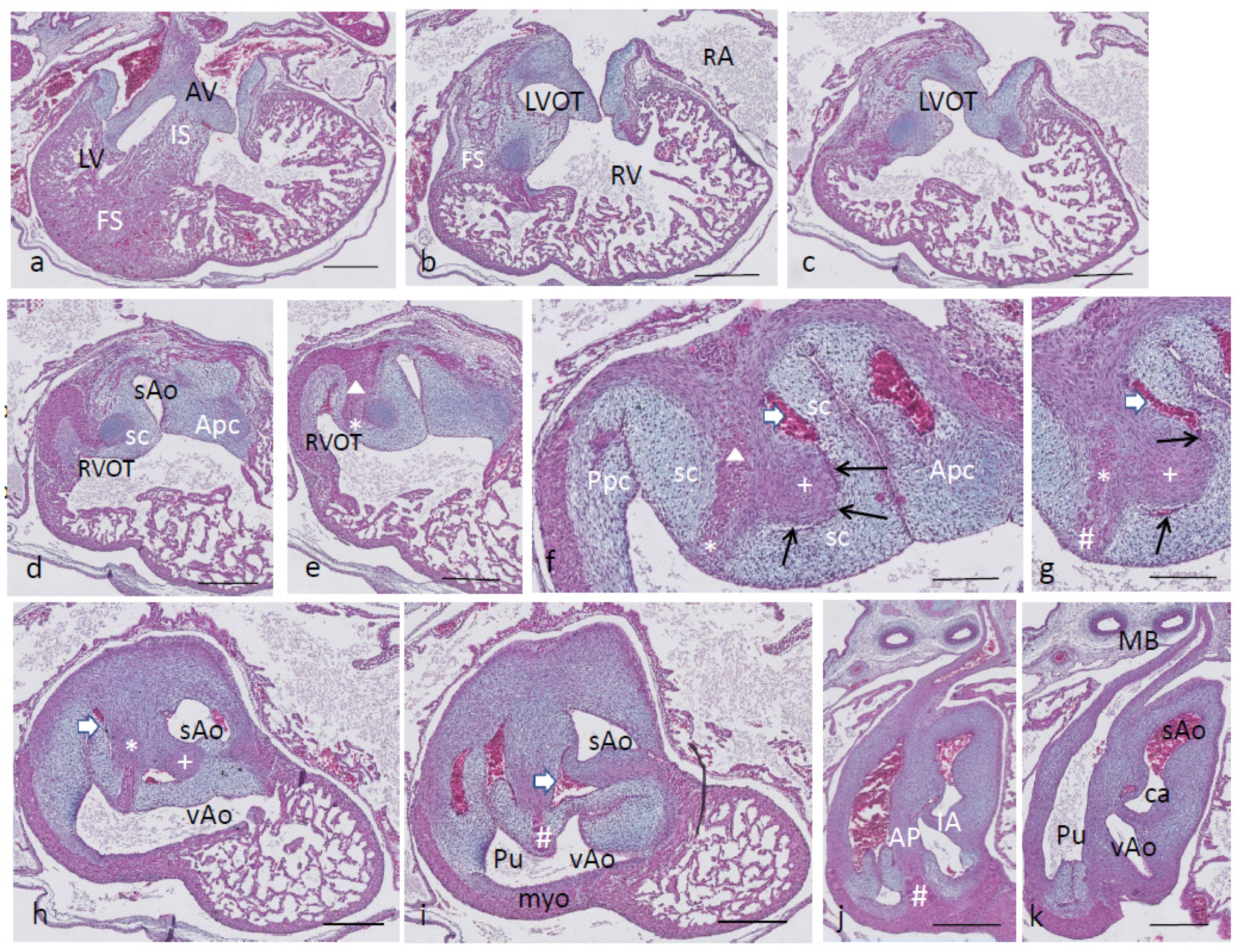
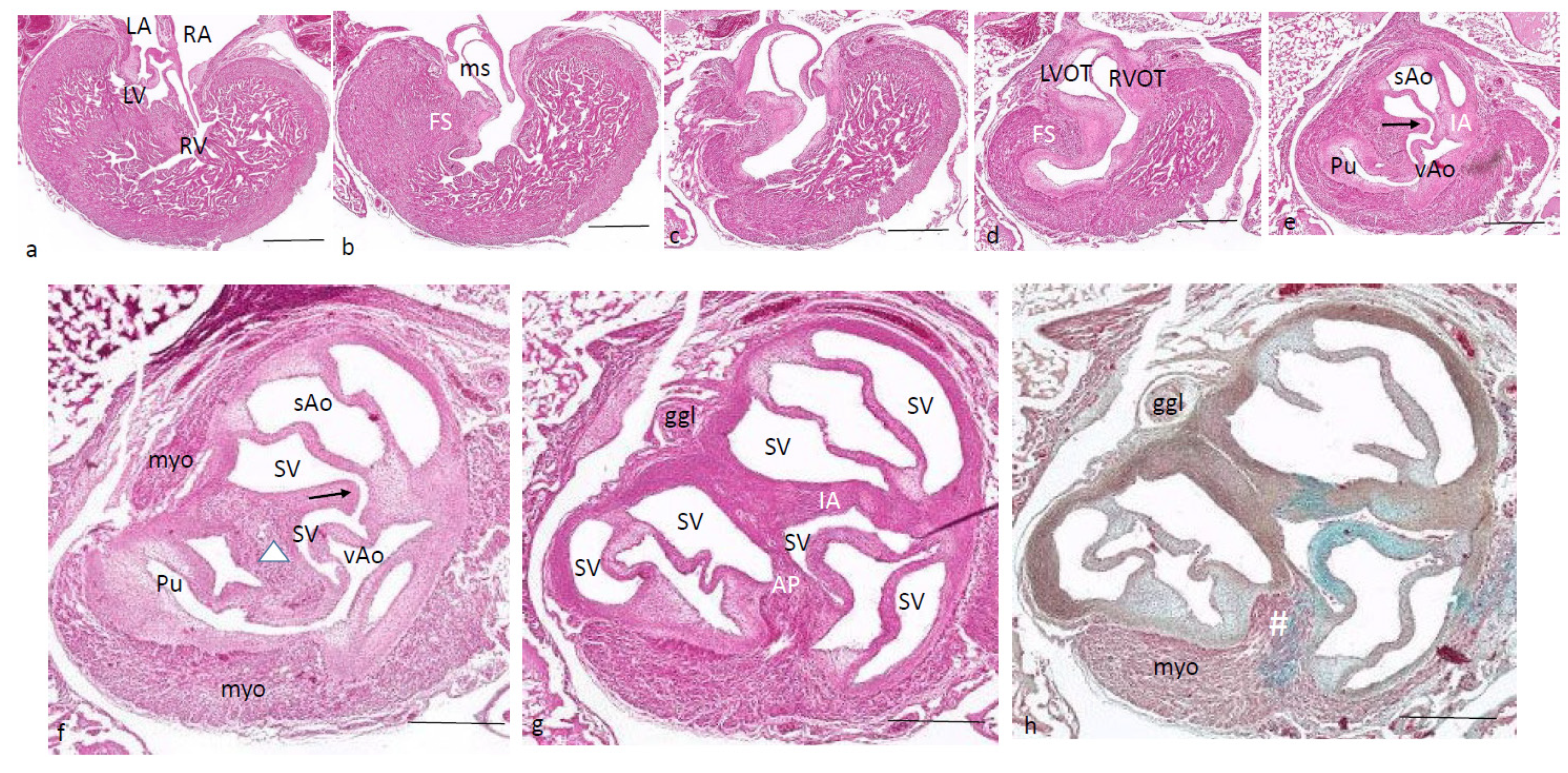
Ferguson stage 24. The folding and inlet septa meet each other at an angle with the septal outflow tract cushion as a hinge (
Figure 6a,b). The inlet septum continues between the LVOT and the right ventricle (
Figure 6b,c), while the septal and aortic parietal cushions have fused over a short distance (
Figure 6d) to reopen again further downstream. A mesenchymal interventricular septum is not apparent yet, in contrast to stage 25. The IA septum is likewise not fully developed, leaving a lumen contact between the sAo and vAo (
Figure 6e,f). Both imply that left–right separation has not been established yet. The cartilage prongs are well differentiated both in the septal cushion and aortic parietal cushion, and in the latter, two cartilage elements are present (
Figure 6e,f), as in earlier stages. The FOP in the septal cushion is wide open, connecting both the vAo’s and sAo’s sinus of Valsalva (
Figure 6f–i). Further downstream, the septal cartilage partly penetrates the interaortic septum (
Figure 6j). In the root of the sAo, the double orifice of the coronary artery is present (orange arrow in
Figure 6k,l), splitting immediately in a descendent and a circumflex branch (
Figure 6l).
Figure 6.
Ferguson stage 24, Figure (a–l), from AV valves to semilunar valve level. Figure (a,b), the AV cushions have elongated to form free-edged valve leaflets. Figure (c,d), folding septum including septal cushion is centrally located between left and right ventricular compartments. Cartilage-rich prongs in both OFT cushions. Note that the parietal cushion now contains two cartilaginous centers. Figure (d’) shows a higher magnification of the hypertrophic chondrocytes. Figure (e–h), the tunnel of the foramen of Panizza (p) is shown rounding the septal cartilage and joining the sinus of Valsalva (sV) of both aortas. Note that the cartilage is surrounded by a fibrous capsule. Figure i, the AP septum is completed. Figure (j), the IA septum is completed. Figure (k,l), coronary ostia (yellow arrows) are found in the parietal wall of the systemic aorta, but not the visceral aorta. Key to the symbols # ventral myocardium, part of AP septum; (presumably) left-sided neural crest cells, part of AP septum; + (presumably) right-sided neural crest, part of IA septum; Δ dorsal myocardium, part of AP septum. Magnification Figure (a), bar 1mm; (b–d,i), bar 500 µm; (e–h,j–l), bar 200 µm; (d’), bar 50 µm.
Figure 6.
Ferguson stage 24, Figure (a–l), from AV valves to semilunar valve level. Figure (a,b), the AV cushions have elongated to form free-edged valve leaflets. Figure (c,d), folding septum including septal cushion is centrally located between left and right ventricular compartments. Cartilage-rich prongs in both OFT cushions. Note that the parietal cushion now contains two cartilaginous centers. Figure (d’) shows a higher magnification of the hypertrophic chondrocytes. Figure (e–h), the tunnel of the foramen of Panizza (p) is shown rounding the septal cartilage and joining the sinus of Valsalva (sV) of both aortas. Note that the cartilage is surrounded by a fibrous capsule. Figure i, the AP septum is completed. Figure (j), the IA septum is completed. Figure (k,l), coronary ostia (yellow arrows) are found in the parietal wall of the systemic aorta, but not the visceral aorta. Key to the symbols # ventral myocardium, part of AP septum; (presumably) left-sided neural crest cells, part of AP septum; + (presumably) right-sided neural crest, part of IA septum; Δ dorsal myocardium, part of AP septum. Magnification Figure (a), bar 1mm; (b–d,i), bar 500 µm; (e–h,j–l), bar 200 µm; (d’), bar 50 µm.
![Jcdd 08 00132 g006]()
Ferguson stage 25. The spongious myocardia of the left and right ventricles have increased considerably in mass (
Figure 7a,b). The cartilage of the septal outflow tract cushion is embedded deep apically between the left and right ventricles squeezed between the inlet and folding segments (
Figure 7a). The presence of the mesenchymal septum (even more visible in the next stage) between the left and right ventricles shows that complete septation has taken place. Even in this stage, the complete separation of the sAo and vAo by the interaortic septum has not been established as there is a narrow lumen contact seen between the LVOT and RVOT (
Figure 7b–h) and, as a consequence, also between the (left ventricular) sAo and (right ventricular) vAo. This implies a complete interventricular septation from now on, but for the narrow connection between the sAo and vAo provided by the FOP (black arrows in
Figure 7e–g) and the narrow lumen contact between both aortas. Functionally, in this embryo, the FOP is closed as no red blood cells are found trapped in its lumen. The vAo appears very compressed in this particular embryo (
Figure 7i).
Figure 7.
Ferguson stage 25, Figure (a–i), from AV to semilunar valve leaflets. The myocardium of this specimen is more compact compared to the earlier stages described here. Figure (a,b), the left and right ventricular compartments are completely septated by the combination of the folding septum, inlet septum and mesenchymal septum. Figure (c,d), the left and right ventricular outflow tracts and the ensuing arterial trunks are separated by the AP septum and the IA septum. The aortic parietal cushion contains two cartilaginous elements. Figure (e,f) show the narrow tunnel of the foramen of Panizza (arrows) through the IA septum (+). Figure (g), the IA septum at this level is uninterrupted, with the flanking sinus of Valsalva of the visceral and systemic aortas. Figure (h,i), both AP and IA septa are obvious, and the lumen of the visceral aorta is very narrow in this specimen. Key to the symbols # ventral myocardium, part of AP septum; (presumably) left-sided neural crest cells, part of AP septum; + (presumably) right-sided neural crest, part of IA septum; Δ dorsal myocardium, part of AP septum. Magnification Figure (a–d,h,i), bar 500 µm; (e–g), bar 200 µm.
Figure 7.
Ferguson stage 25, Figure (a–i), from AV to semilunar valve leaflets. The myocardium of this specimen is more compact compared to the earlier stages described here. Figure (a,b), the left and right ventricular compartments are completely septated by the combination of the folding septum, inlet septum and mesenchymal septum. Figure (c,d), the left and right ventricular outflow tracts and the ensuing arterial trunks are separated by the AP septum and the IA septum. The aortic parietal cushion contains two cartilaginous elements. Figure (e,f) show the narrow tunnel of the foramen of Panizza (arrows) through the IA septum (+). Figure (g), the IA septum at this level is uninterrupted, with the flanking sinus of Valsalva of the visceral and systemic aortas. Figure (h,i), both AP and IA septa are obvious, and the lumen of the visceral aorta is very narrow in this specimen. Key to the symbols # ventral myocardium, part of AP septum; (presumably) left-sided neural crest cells, part of AP septum; + (presumably) right-sided neural crest, part of IA septum; Δ dorsal myocardium, part of AP septum. Magnification Figure (a–d,h,i), bar 500 µm; (e–g), bar 200 µm.
![Jcdd 08 00132 g007]()
Beyond Ferguson stage 25 (alligator). In this HE-stained series, the LV and RV are completely septated, and the hallmark of this process is the mesenchymal septum extending over a considerable distance (
Figure 8a–d), reaching as far as the IA septum (
Figure 8d,e). In the outflow tract, the septal cushion is penetrated by the FOP (black arrow), connecting both septal sinuses of Valsalva of the sAo and the vAo (
Figure 8e,f) at the level where the arterial wall is inserted into the myocardium. Slightly more downstream, the IA septum separates the two aortas, whereas the AP septum completely separates the pulmonary trunk from the vAo (
Figure 8g,h). Note that
Figure 8h is Alcian blue stained to demonstrate the myocardium versus the mesenchymal wall of the arteries.
Figure 8.
Alligator embryo older > than F stage 25. HE-stained Figure (h) is, in addition, Alcian blue stained. Figure (a–h) from AV valves to semilunar valves. Figure (a–d), the mesenchymal septum is inserted between the right and left ventricular compartments separating both ventricles. It reaches from the tip of the folding septum near the septal cushion (in Figure (a)) towards the IA septum (in Figure (d,e)). The communication between visceral and systemic aortas is still patent (Figure (d)). Figure (e,f), here, the IA septum is interrupted by the tunnel of the foramen of Panizza (arrow). Figure (g,h), both the AP septum and the IA septum are complete at this level. Key to the symbols # ventral myocardium, part of AP septum; (presumably) left-sided neural crest cells, part of AP septum; + (presumably) right-sided neural crest, part of IA septum; Δ dorsal myocardium, part of AP septum. Magnification Figure (a–e), bar 200 µm; (f–h), bar 500 µm.
Figure 8.
Alligator embryo older > than F stage 25. HE-stained Figure (h) is, in addition, Alcian blue stained. Figure (a–h) from AV valves to semilunar valves. Figure (a–d), the mesenchymal septum is inserted between the right and left ventricular compartments separating both ventricles. It reaches from the tip of the folding septum near the septal cushion (in Figure (a)) towards the IA septum (in Figure (d,e)). The communication between visceral and systemic aortas is still patent (Figure (d)). Figure (e,f), here, the IA septum is interrupted by the tunnel of the foramen of Panizza (arrow). Figure (g,h), both the AP septum and the IA septum are complete at this level. Key to the symbols # ventral myocardium, part of AP septum; (presumably) left-sided neural crest cells, part of AP septum; + (presumably) right-sided neural crest, part of IA septum; Δ dorsal myocardium, part of AP septum. Magnification Figure (a–e), bar 200 µm; (f–h), bar 500 µm.














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}