
Dissecting the Complexity of Early Heart Progenitor Cells


Abstract
:1. Introduction
- Raw and processed data https://www.ebi.ac.uk/arrayexpress/experiments/E-MTAB-6153/
- Raw and processed data https://www.ebi.ac.uk/ena/browser/view/PRJEB23303?show=reads
- Raw and processed data https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE169210; Browser https://tanaylab.weizmann.ac.il/embflow/
- Raw, processed data and Browser https://marionilab.cruk.cam.ac.uk/heartAtlas/;
- Raw and processed data https://www.ebi.ac.uk/ena/browser/view/PRJEB23303?show=reads;
- Raw and processed data https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE133244; Browser https://gottgens-lab.stemcells.cam.ac.uk/snATACseq_E825/
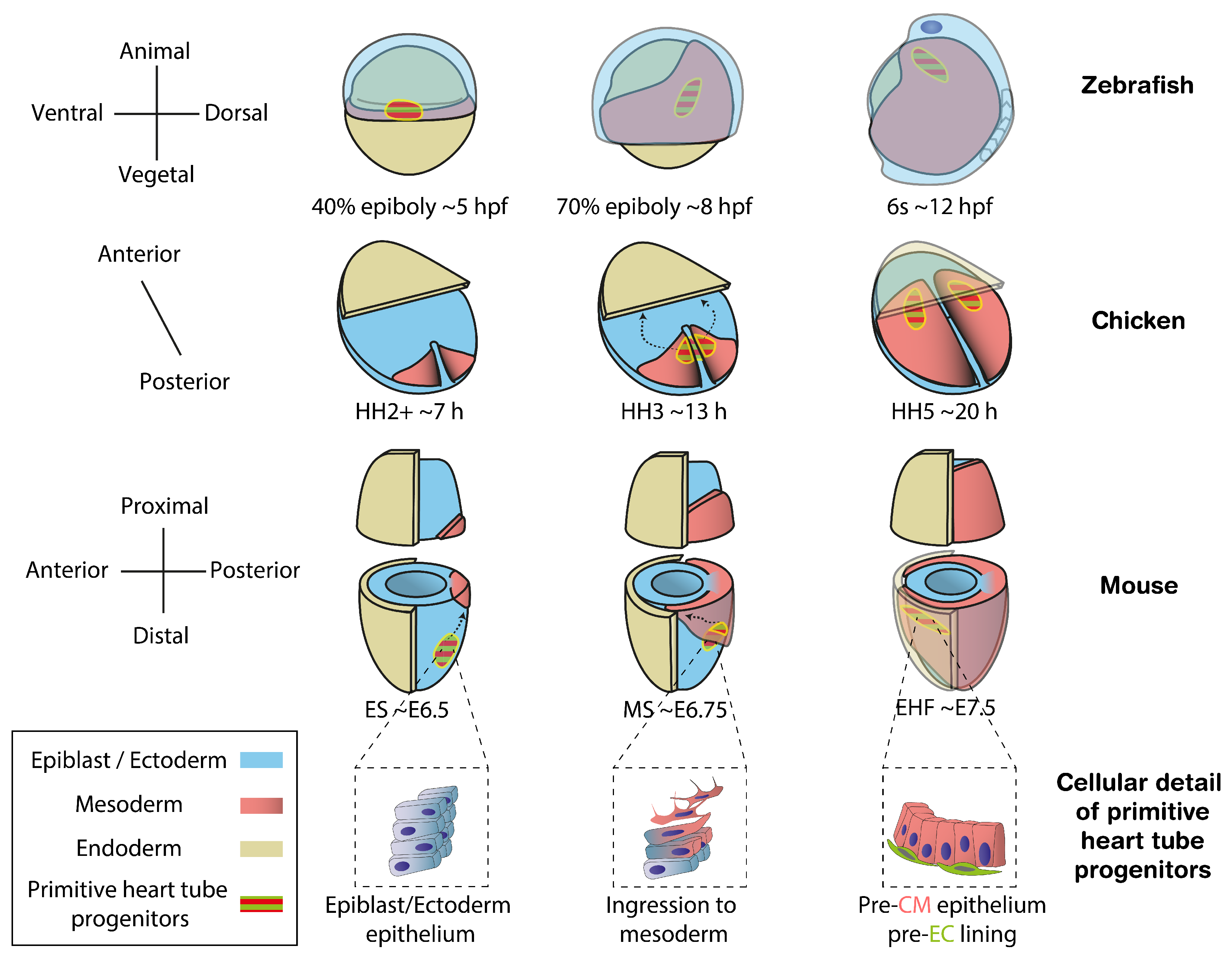
2. Cell Fate Specification Preceding Primitive Heart Tube Formation
2.1. Temporal Sequence of Fate Acquisition
2.2. Molecular Mechanisms of Specification
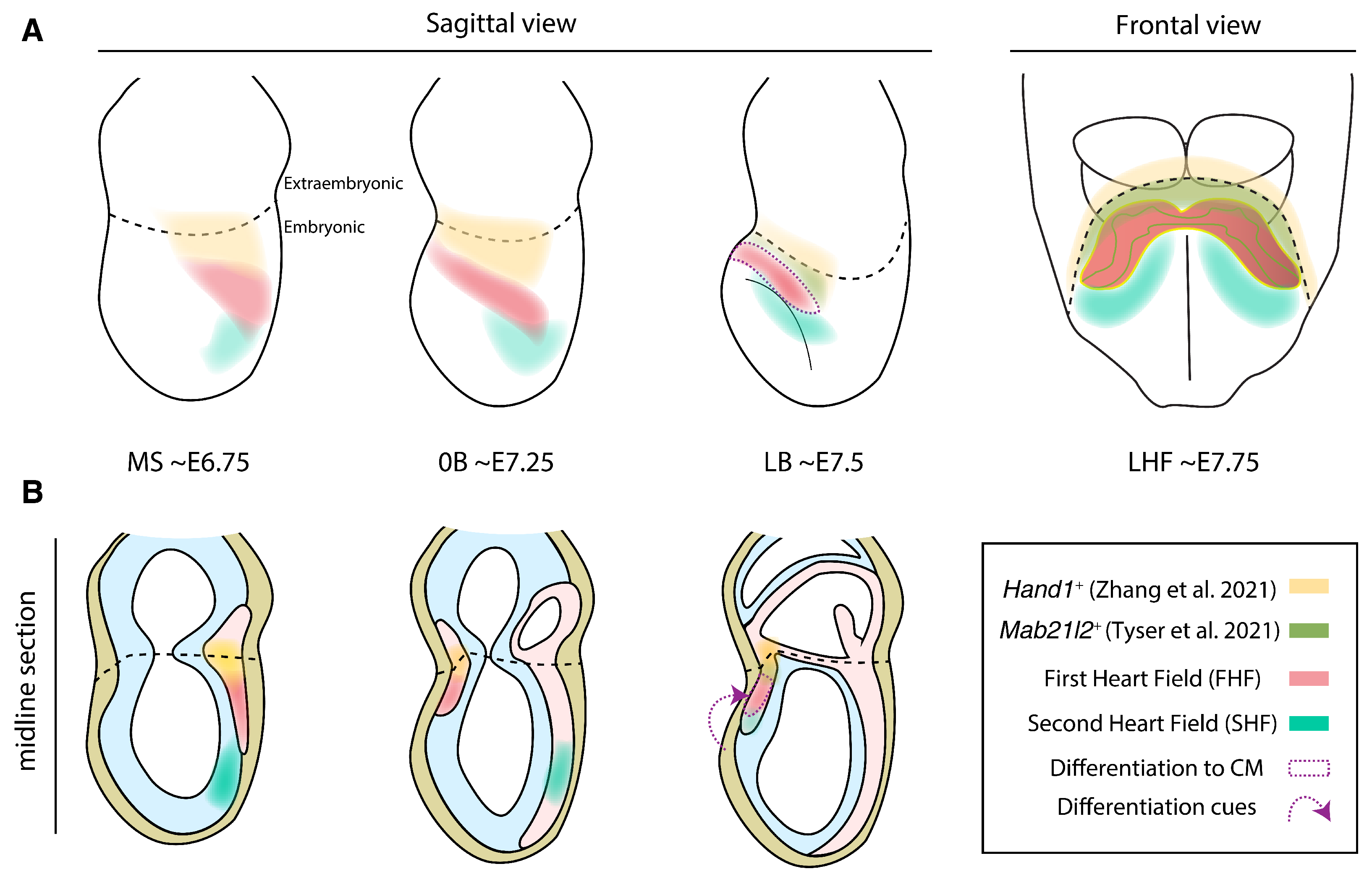
3. Differentiation of Primitive Heart Tube Progenitors
3.1. Signaling Cues Driving CM and EC Differentiation
3.2. Redirection of Cardiac Progenitor Differentiation upon Perturbation
3.3. Plasticity of Cardiac Progenitors
4. Molecular Heterogeneity of the Cardiomyocyte Sources within the Primitive Heart Tube
5. Discussion and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Meilhac, S.M.; Buckingham, M.E. The deployment of cell lineages that form the mammalian heart. Nat. Rev. Cardiol. 2018, 15, 705–724. [Google Scholar] [CrossRef]
- Zimmerman, M.S.; Smith, A.G.C.; Sable, C.A.; Echko, M.M.; Wilner, L.B.; Olsen, H.E.; Atalay, H.T.; Awasthi, A.; Bhutta, Z.A.; Boucher, J.L.A.; et al. Global, regional, and national burden of congenital heart disease, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet Child Adolesc. Health 2020, 4, 185–200. [Google Scholar] [CrossRef] [Green Version]
- Gittenberger-de Groot, A.C.; Winter, E.M.; Bartelings, M.M.; Jose Goumans, M.; DeRuiter, M.C.; Poelmann, R.E. The arterial and cardiac epicardium in development, disease and repair. Differentiation 2012, 84, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Gittenberger-De Groot, A.C.; Moulaert, A.J.; Harinck, E.; Becker, A.E. Histopathology of the ductus arteriosus after prostaglandin E1 administration in ductus dependent cardiac anomalies. Br. Heart J. 1978, 40, 215–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawson, K.A.; Pedersen, R.A. Clonal analysis of cell fate during gastrulation and early neurulation in the mouse. Ciba Foundation Symp. 1992, 165, 3–21. [Google Scholar] [CrossRef]
- Lawson, K.A.; Meneses, J.J.; Pedersen, R.A. Clonal analysis of epiblast fate during germ layer formation in the mouse embryo. Development 1991, 113, 891–911. [Google Scholar] [CrossRef]
- Tam, P.P.; Zhou, S.X. The allocation of epiblast cells to ectodermal and germ-line lineages is influenced by the position of the cells in the gastrulating mouse embryo. Dev. Biol. 1996, 178, 124–132. [Google Scholar] [CrossRef]
- Tam, P.P.; Behringer, R.R. Mouse gastrulation: The formation of a mammalian body plan. Mech. Dev. 1997, 68, 3–25. [Google Scholar] [CrossRef]
- Saga, Y.; Miyagawa-Tomita, S.; Takagi, A.; Kitajima, S.; Miyazaki, J.I.; Inoue, T. MesP1 is expressed in the heart precursor cells and required for the formation of a single heart tube. Development 1999, 126, 3437–3447. [Google Scholar] [CrossRef]
- Street, D. Early differentiation of the heart in mouse embryos. J. Anat. 1981, 133, 235–246. [Google Scholar]
- Harris, I.S.; Black, B.L. Development of the endocardium. Pediatr. Cardiol. 2010, 31, 391–399. [Google Scholar] [CrossRef] [Green Version]
- Buckingham, M.; Meilhac, S.; Zaffran, S. Building the mammalian heart from two sources of myocardial cells. Nat. Rev. Genet. 2005, 6, 826–835. [Google Scholar] [CrossRef]
- Ivanovitch, K.; Temiño, S.; Torres, M. Live imaging of heart tube development in mouse reveals alternating phases of cardiac differentiation and morphogenesis. eLife 2017, 6, e30668. [Google Scholar] [CrossRef]
- Downs, K.M.; Davies, T. Staging of gastrulating mouse embryos by morphological landmarks in the dissecting microscope. Development (Camb. Engl.) 1993, 118, 1255–1266. [Google Scholar] [CrossRef]
- Jensen, B.; Christoffels, V.M.; Moorman, A.F. An appreciation of anatomy in the molecular world. J. Cardiovasc. Dev. Dis. 2020, 7, 44. [Google Scholar] [CrossRef] [PubMed]
- Kelly, R.G.; Buckingham, M.E.; Moorman, A.F. Heart fields and cardiac morphogenesis. Cold Spring Harb. Perspect. Med. 2014, 4, a015750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kemmler, C.L.; Riemslagh, F.W.; Moran, H.R.; Mosimann, C. From stripes to a beating heart: Early cardiac development in zebrafish. J. Cardiovasc. Dev. Dis. 2021, 8, 17. [Google Scholar] [CrossRef] [PubMed]
- Tyser, R.C.; Ibarra-Soria, X.; McDole, K.; Jayaram, S.A.; Godwin, J.; Brand, T.A.D.; Miranda, A.M.; Scialdone, A.; Keller, P.J.; Marioni, J.C.; et al. Characterization of a common progenitor pool of the epicardium and myocardium. Science 2021, 371, eabb2986. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Carlin, D.; Zhu, F.; Cattaneo, P.; Ideker, T.; Evans, S.M.; Bloomekatz, J.; Chi, N.C. Unveiling Complexity and Multipotentiality of Early Heart Fields. Circ. Res. 2021, 129, 474–487. [Google Scholar] [CrossRef]
- Lee, R.K.; Stainier, D.Y.; Weinstein, B.M.; Fishman, M.C. Cardiovascular development in the zebrafish II. Endocardial progenitors are sequestered within the heart field. Development 1994, 120, 3361–3366. [Google Scholar] [CrossRef] [PubMed]
- Keegan, B.R. Organization of cardiac chamber progenitors in the zebrafish blastula. Development 2004, 131, 3081–3091. [Google Scholar] [CrossRef] [Green Version]
- Mao, L.M.; Boyle Anderson, E.A.; Ho, R.K. Anterior lateral plate mesoderm gives rise to multiple tissues and requires tbx5a function in left-right asymmetry, migration dynamics, and cell specification of late-addition cardiac cells. Dev. Biol. 2021, 472, 52–66. [Google Scholar] [CrossRef]
- Wei, Y.; Mikawa, T. Fate diversity of primitive streak cells during heart field formation in ovo. Dev. Dyn. 2000, 219, 505–513. [Google Scholar] [CrossRef]
- Lescroart, F.; Chabab, S.; Lin, X.; Rulands, S.; Paulissen, C.; Rodolosse, A.; Auer, H.; Achouri, Y.; Dubois, C.; Bondue, A.; et al. Early lineage restriction in temporally distinct populations of Mesp1 progenitors during mammalian heart development. Nat. Cell Biol. 2014, 16, 829–840. [Google Scholar] [CrossRef]
- Scialdone, A.; Tanaka, Y.; Jawaid, W.; Moignard, V.; Wilson, N.K.; Macaulay, I.C.; Marioni, J.C.; Göttgens, B. Resolving early mesoderm diversification through single-cell expression profiling. Nature 2016, 535, 289–293. [Google Scholar] [CrossRef] [Green Version]
- Lescroart, F.; Wang, X.; Lin, X.; Swedlund, B.; Gargouri, S.; Sànchez-dànes, A.; Dubois, C.; Paulissen, C.; Kinston, S.; Göttgens, B.; et al. Defining the earliest step of cardiovascular lineage segregation by single-cell RNA-seq. Science 2018, 4174, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibarra-Soria, X.; Jawaid, W.; Pijuan-Sala, B.; Ladopoulos, V.; Scialdone, A.; Jörg, D.J.; Tyser, R.C.; Calero-Nieto, F.J.; Mulas, C.; Nichols, J.; et al. Defining murine organogenesis at single-cell resolution reveals a role for the leukotriene pathway in regulating blood progenitor formation. Nat. Cell Biol. 2018, 20, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Pijuan-Sala, B.; Griffiths, J.A.; Guibentif, C.; Hiscock, T.W.; Jawaid, W.; Calero-Nieto, F.J.; Mulas, C.; Ibarra-Soria, X.; Tyser, R.C.; Ho, D.L.L.; et al. A single-cell molecular map of mouse gastrulation and early organogenesis. Nature 2019, 566, 490–495. [Google Scholar] [CrossRef] [PubMed]
- de Soysa, T.Y.; Ranade, S.S.; Okawa, S.; Ravichandran, S.; Huang, Y.; Salunga, H.T.; Schricker, A.; del Sol, A.; Gifford, C.A.; Srivastava, D. Single-cell analysis of cardiogenesis reveals basis for organ-level developmental defects. Nature 2019, 572, 120–124. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.; Pei, Y.; He, L.; Peng, G.; Reinius, B.; Tam, P.P.; Jing, N.; Deng, Q. Single-Cell RNA-Seq Reveals Cellular Heterogeneity of Pluripotency Transition and X Chromosome Dynamics during Early Mouse Development. Cell Rep. 2019, 26, 2593–2607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, H.; Luo, Y.; Yue, Y.; Zhang, J.; Ai, S.; Li, X.; Wang, X.; Zhang, Y.L.; Wei, Y.; Li, H.H.; et al. Single-Cell Transcriptomics Reveals Chemotaxis-Mediated Intraorgan Crosstalk during Cardiogenesis. Circ. Res. 2019, 125, 398–410. [Google Scholar] [CrossRef] [PubMed]
- Jia, G.; Preussner, J.; Chen, X.; Guenther, S.; Yuan, X.; Yekelchyk, M.; Kuenne, C.; Looso, M.; Zhou, Y.; Teichmann, S.; et al. Single cell RNA-seq and ATAC-seq analysis of cardiac progenitor cell transition states and lineage settlement. Nat. Commun. 2018, 9, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mittnenzweig, M.; Mayshar, Y.; Cheng, S.; Ben-Yair, R.; Hadas, R.; Rais, Y.; Chomsky, E.; Reines, N.; Uzonyi, A.; Lumerman, L.; et al. A single-embryo, single-cell time-resolved model for mouse gastrulation. Cell 2021, 184, 2825–2842. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Shi, J.; Tao, Y.; Zernicka-Goetz, M. Tracing the origin of heterogeneity and symmetry breaking in the early mammalian embryo. Nat. Commun. 2018, 9, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Preissl, S.; Fang, R.; Huang, H.; Zhao, Y.; Raviram, R.; Gorkin, D.U.; Zhang, Y.; Sos, B.C.; Afzal, V.; Dickel, D.E.; et al. Single-nucleus analysis of accessible chromatin in developing mouse forebrain reveals cell-type-specific transcriptional regulation. Nat. Neurosci. 2018, 21, 432–439. [Google Scholar] [CrossRef] [Green Version]
- Clark, S.J.; Argelaguet, R.; Kapourani, C.A.; Stubbs, T.M.; Lee, H.J.; Alda-Catalinas, C.; Krueger, F.; Sanguinetti, G.; Kelsey, G.; Marioni, J.C.; et al. ScNMT-seq enables joint profiling of chromatin accessibility DNA methylation and transcription in single cells. Nat. Commun. 2018, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Argelaguet, R.; Clark, S.J.; Mohammed, H.; Stapel, L.C.; Krueger, C.; Kapourani, C.A.; Imaz-Rosshandler, I.; Lohoff, T.; Xiang, Y.; Hanna, C.W.; et al. Multi-omics profiling of mouse gastrulation at single-cell resolution. Nature 2019, 576, 487–491. [Google Scholar] [CrossRef]
- McDole, K.; Guignard, L.; Amat, F.; Berger, A.; Malandain, G.; Royer, L.A.; Turaga, S.C.; Branson, K.; Keller, P.J. In Toto Imaging and Reconstruction of Post-Implantation Mouse Development at the Single-Cell Level. Cell 2018, 175, 859–876. [Google Scholar] [CrossRef] [Green Version]
- Saykali, B.; Mathiah, N.; Nahaboo, W.; Racu, M.L.; Hammou, L.; Defrance, M.; Migeotte, I. Distinct mesoderm migration phenotypes in extra-embryonic and embryonic regions of the early mouse embryo. eLife 2019, 8, e42434. [Google Scholar] [CrossRef]
- Luecken, M.D.; Theis, F.J. Current best practices in single-cell RNA-seq analysis: A tutorial. Mol. Syst. Biol. 2019, 15, e8746. [Google Scholar] [CrossRef]
- Rosenquist, G.C. Location and movements of cardiogenic cells in the chick embryo: The heart-forming portion of the primitive streak. Dev. Biol. 1970, 22, 461–475. [Google Scholar] [CrossRef]
- Stainier, D.Y.; Weinstein, B.M.; Detrich, H.W.; Zon, L.I.; Fishman, M.C. Cloche, an early acting zebrafish gene, is required by both the endothelial and hematopoietic lineages. Development 1995, 121, 3141–3150. [Google Scholar] [CrossRef] [PubMed]
- Gassmann, M.; Casagranda, F.; Orloli, D.; Simon, H.; Lai, C.; Kleint, R.; Lemke, G. Aberrant neural and cardiac development in mice lacking the ErbB4 neuregulin receptor. Nature 1995, 378, 390–394. [Google Scholar] [CrossRef] [PubMed]
- Peshkovsky, C.; Totong, R.; Yelon, D. Dependence of cardiac trabeculation on neuregulin signaling and blood flow in zebrafish. Dev. Dyn. 2011, 240, 446–456. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, Y.; Mironov, V.; Yamagishi, T.; Nakamura, H.; Markwald, R.R. Expression of smooth muscle alpha-actin in mesenchymal cells during formation of avian endocardial cushion tissue: A role for transforming growth factor β3. Dev. Dyn. 1997, 209, 296–309. [Google Scholar] [CrossRef]
- Brown, C.B.; Boyer, A.S.; Runyan, R.B.; Barnett, J.V. Antibodies to the type II TGFβ receptor block cell activation and migration during atrioventricular cushion transformation in the heart. Dev. Biol. 1996, 174, 248–257. [Google Scholar] [CrossRef] [Green Version]
- Feng, W.; Chen, L.; Nguyen, P.K.; Wu, S.M.; Li, G. Single Cell Analysis of Endothelial Cells Identified Organ-Specific Molecular Signatures and Heart-Specific Cell Populations and Molecular Features. Front. Cardiovasc. Med. 2019, 6, 165. [Google Scholar] [CrossRef]
- Drake, C.J.; Fleming, P.A. Vasculogenesis in the day 6.5 to 9.5 mouse embryo. Blood 2000, 95, 1671–1679. [Google Scholar] [CrossRef]
- De La Pompa, J.L.; Timmerman, L.A.; Takimoto, H.; Yoshida, H.; Elia, A.J.; Samper, E.; Potter, J.; Wakeham, A.; Marengere, L.; Langille, B.L.; et al. Role of the NF-ATc transcription factor in morphogenesis of cardiac valves and septum. Nature 1998, 392, 182–186. [Google Scholar] [CrossRef]
- Poelmann, R.E.; Gittenberger-de Groot, A.C. Development and evolution of the metazoan heart. Dev. Dyn. 2019, 248, 634–656. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Chápuli, R.; Carmona, R.; Guadix, J.A.; Macías, D.; Pérez-Pomares, J.M. The origin of the endothelial cells: An evo-devo approach for the invertebrate/vertebrate transition of the circulatory system. Evol. Dev. 2005, 7, 351–358. [Google Scholar] [CrossRef]
- Nakano, A.; Nakano, H.; Smith, K.A.; Palpant, N.J. The developmental origins and lineage contributions of endocardial endothelium. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2016, 1863, 1937–1947. [Google Scholar] [CrossRef] [PubMed]
- Conklin, E.G. Mosaic development in ascidian eggs. J. Exp. Zool. 1905, 2, 145–223. [Google Scholar] [CrossRef]
- Petit, A.C.; Legué, E.; Nicolas, J.F. Methods in clonal analysis and applications. Reprod. Nutr. Dev. 2005, 45, 321–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Martinez, V.; Schoenwolf, G.C. Primitive-streak origin of the cardiovascular system in avian embryos. Dev. Biol. 1993, 159, 706–719. [Google Scholar] [CrossRef] [PubMed]
- Stalsberg, H.; DeHaan, R.L. The precardiac areas and formation of the tubular heart in the chick embryo. Dev. Biol. 1969, 19, 128–159. [Google Scholar] [CrossRef]
- Mikawa, T.; Borisov, A.; Brown, A.M.; Fischman, D.A. Clonal analysis of cardiac morphogenesis in the chicken embryo using a replication-defective retrovirus: I. Formation of the ventricular myocardium. Dev. Dyn. 1992, 193, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Devine, W.P.; Wythe, J.D.; George, M.; Koshiba-Takeuchi, K.; Bruneau, B.G. Early patterning and specification of cardiac progenitors in gastrulating mesoderm. eLife 2014, 3, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Kattman, S.J.; Huber, T.L.; Keller, G.M. Multipotent Flk-1+ Cardiovascular Progenitor Cells Give Rise to the Cardiomyocyte, Endothelial, and Vascular Smooth Muscle Lineages. Dev. Cell 2006, 11, 723–732. [Google Scholar] [CrossRef] [Green Version]
- Milgrom-Hoffman, M.; Harrelson, Z.; Ferrara, N.; Zelzer, E.; Evans, S.M.; Tzahor, E. The heart endocardium is derived from vascular endothelial progenitors. Development 2011, 138, 4777–4787. [Google Scholar] [CrossRef] [Green Version]
- Misfeldt, A.M.; Boyle, S.C.; Tompkins, K.L.; Bautch, V.L.; Labosky, P.A.; Baldwin, H.S. Endocardial cells are a distinct endothelial lineage derived from Flk1+ multipotent cardiovascular progenitors. Dev. Biol. 2009, 333, 78–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Motoike, T.; Markham, D.W.; Rossant, J.; Sato, T.N. Evidence for novel fate of Flk1+ progenitor: Contribution to muscle lineage. Genesis 2003, 35, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Bu, L.; Jiang, X.; Martin-Puig, S.; Caron, L.; Zhu, S.; Shao, Y.; Roberts, D.J.; Huang, P.L.; Domian, I.J.; Chien, K.R. Human ISL1 heart progenitors generate diverse multipotent cardiovascular cell lineages. Nature 2009, 460, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Tyser, R.C.; Mahammadov, E.; Nakanoh, S.; Vallier, L.; Scialdone, A.; Srinivas, S. Single-cell transcriptomic characterization of a gastrulating human embryo. Nature 2021, 600, 285–289. [Google Scholar] [CrossRef]
- Moretti, A.; Caron, L.; Nakano, A.; Lam, J.T.; Bernshausen, A.; Chen, Y.; Qyang, Y.; Bu, L.; Sasaki, M.; Martin-Puig, S.; et al. Multipotent Embryonic Isl1+Progenitor Cells Lead to Cardiac, Smooth Muscle, and Endothelial Cell Diversification. Cell 2006, 127, 1151–1165. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.M.; Fujiwara, Y.; Cibulsky, S.M.; Clapham, D.E.; Ling Lien, C.; Schultheiss, T.M.; Orkin, S.H. Developmental Origin of a Bipotential Myocardial and Smooth Muscle Cell Precursor in the Mammalian Heart. Cell 2006, 127, 1137–1150. [Google Scholar] [CrossRef] [Green Version]
- Stanley, E.G.; Biben, C.; Elefanty, A.; Barnett, L.; Koentgen, F.; Robb, L.; Harvey, R.P. Efficient cre-mediated deletion in cardiac progenitor cells conferred by a 3’UTR-ires-Cre allele of the homeobox gene Nkx2-5. Int. J. Dev. Biol. 2002, 46, 431–439. [Google Scholar] [CrossRef]
- Bussmann, J.; Bakkers, J.; Schulte-Merker, S. Early endocardial morphogenesis requires Scl/Tal1. PLoS Genet. 2007, 3, 1425–1437. [Google Scholar] [CrossRef]
- Schoenebeck, J.J.; Keegan, B.R.; Yelon, D. Vessel and Blood Specification Override Cardiac Potential in Anterior Mesoderm. Dev. Cell 2007, 13, 254–267. [Google Scholar] [CrossRef] [Green Version]
- Chan, M.M.; Smith, Z.D.; Grosswendt, S.; Kretzmer, H.; Norman, T.M.; Adamson, B.; Jost, M.; Quinn, J.J.; Yang, D.; Jones, M.G.; et al. Molecular recording of mammalian embryogenesis. Nature 2019, 570, 77–82. [Google Scholar] [CrossRef] [Green Version]
- McKenna, A.; Findlay, G.M.; Gagnon, J.A.; Horwitz, M.S.; Schier, A.F.; Shendure, J. Whole-organism lineage tracing by combinatorial and cumulative genome editing. Science 2016, 353, aaf7907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elsasser, W.M. Outline of a theory of cellular heterogeneity. Proc. Natl. Acad. Sci. USA 1984, 81, 5126–5129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, J.; Spielmann, M.; Qiu, X.; Huang, X.; Ibrahim, D.M.; Hill, A.J.; Zhang, F.; Mundlos, S.; Christiansen, L.; Steemers, F.J.; et al. The single-cell transcriptional landscape of mammalian organogenesis. Nature 2019, 566, 496–502. [Google Scholar] [CrossRef] [PubMed]
- Haghverdi, L.; Büttner, M.; Wolf, F.A.; Buettner, F.; Theis, F.J. Diffusion pseudotime robustly reconstructs lineage branching. Nat. Methods 2016, 13, 845–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cusanovich, D.A.; Reddington, J.P.; Garfield, D.A.; Daza, R.M.; Aghamirzaie, D.; Marco-Ferreres, R.; Pliner, H.A.; Christiansen, L.; Qiu, X.; Steemers, F.J.; et al. The cis-regulatory dynamics of embryonic development at single-cell resolution. Nature 2018, 555, 538–542. [Google Scholar] [CrossRef] [PubMed]
- Pijuan-Sala, B.; Wilson, N.K.; Xia, J.; Hou, X.; Hannah, R.L.; Kinston, S.; Calero-Nieto, F.J.; Poirion, O.; Preissl, S.; Liu, F.; et al. Single-cell chromatin accessibility maps reveal regulatory programs driving early mouse organogenesis. Nat. Cell Biol. 2020, 22, 487–497. [Google Scholar] [CrossRef] [PubMed]
- Crispatzu, G.; Rehimi, R.; Pachano, T.; Bleckwehl, T.; Cruz-Molina, S.; Xiao, C.; Mahabir, E.; Bazzi, H.; Rada-Iglesias, A. The chromatin, topological and regulatory properties of pluripotency-associated poised enhancers are conserved in vivo. Nat. Commun. 2021, 12, 4344. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Yue, F.; Li, Y.; Xie, R.; Harper, T.; Patel, N.A.; Muth, K.; Palmer, J.; Qiu, Y.; Wang, J.; et al. Epigenetic priming of enhancers predicts developmental competence of hESC-derived endodermal lineage intermediates. Cell Stem Cell 2015, 16, 386–399. [Google Scholar] [CrossRef] [Green Version]
- Grosswendt, S.; Kretzmer, H.; Smith, Z.D.; Kumar, A.S.; Hetzel, S.; Wittler, L.; Klages, S.; Timmermann, B.; Mukherji, S.; Meissner, A. Epigenetic regulator function through mouse gastrulation. Nature 2020, 584, 102–108. [Google Scholar] [CrossRef]
- Takeuchi, J.K.; Bruneau, B.G. Directed transdifferentiation of mouse mesoderm to heart tissue by defined factors. Nature 2009, 459, 708–711. [Google Scholar] [CrossRef] [Green Version]
- Arai, A.; Yamamoto, K.; Toyama, J. Murine cardiac progenitor cells require visceral embryonic endoderm and primitive streak for terminal differentiation. Dev. Dyn. 1997, 210, 344–353. [Google Scholar] [CrossRef]
- Varner, V.D.; Taber, L.A. Not just inductive: A crucial mechanical role for the endoderm during heart tube assembly. Development 2012, 139, 1680–1690. [Google Scholar] [CrossRef] [Green Version]
- Rowton, M.; Guzzetta, A.; Rydeen, A.B.; Moskowitz, I.P. Control of cardiomyocyte differentiation timing by intercellular signaling pathways. Semin. Cell Dev. Biol. 2021, 118, 94–106. [Google Scholar] [CrossRef]
- Alsan, B.H.; Schultheiss, T.M. Regulation of avian cardiogenesis by Fgf8 signaling. Development 2002, 129, 1935–1943. [Google Scholar] [CrossRef]
- Brown, K.; Doss, M.X.; Legros, S.; Artus, J.; Hadjantonakis, A.K.; Foley, A.C. Extraembryonic endoderm (XEN) stem cells producefactors that activate heart formation. PLoS ONE 2010, 5, e13446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tzahor, E. Wnt/β-Catenin Signaling and Cardiogenesis: Timing Does Matter. Dev. Cell 2007, 13, 10–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, M.; Tang, Y.; Zhou, Y.; Zhang, J. Deciphering Role of Wnt Signalling in Cardiac Mesoderm and Cardiomyocyte Differentiation from Human iPSCs: Four-dimensional control of Wnt pathway for hiPSC-CMs differentiation. Sci. Rep. 2019, 9, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Lewis-Israeli, Y.R.; Wasserman, A.H.; Aguirre, A. Heart organoids and engineered heart tissues: Novel tools for modeling human cardiac biology and disease. Biomolecules 2021, 11, 1277. [Google Scholar] [CrossRef]
- Reichman, D.E.; Park, L.; Man, L.; Redmond, D.; Chao, K.; Harvey, R.P.; Taketo, M.M.; Rosenwaks, Z.; James, D. Wnt inhibition promotes vascular specification of embryonic cardiac progenitors. Development 2018, 145, dev159905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, C.; Khan, M.A.; Wong, F.; Solovieva, T.; Oliveira, N.M.; Baldock, R.A.; Tickle, C.; Burt, D.W.; Stern, C.D. A strategy to discover new organizers identifies a putative heart organizer. Nat. Commun. 2016, 7, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Nemir, M.; Croquelois, A.; Pedrazzini, T.; Radtke, F. Induction of cardiogenesis in embryonic stem cells via downregulation of Notch1 signaling. Circ. Res. 2006, 98, 1471–1478. [Google Scholar] [CrossRef] [Green Version]
- Schroeder, T.; Meier-Stiegen, F.; Schwanbeck, R.; Eilken, H.; Nishikawa, S.; Häsler, R.; Schreiber, S.; Bornkamm, G.W.; Nishikawa, S.I.; Just, U. Activated Notch1 alters differentiation of embryonic stem cells into mesodermal cell lineages at multiple stages of development. Mech. Dev. 2006, 123, 570–579. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Bodmer, R. Myogenic cells fates are antagonized by Notch only in asymmetric lineages of the Drosophila heart, with or without cell division. Development 2003, 130, 3039–3051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rones, M.S.; McLaughlin, K.A.; Raffin, M.; Mercola, M. Serrate and Notch specify cell fates in the heart field by suppressing cardiomyogenesis. Development 2000, 127, 3865–3876. [Google Scholar] [CrossRef] [PubMed]
- Chau, M.D.; Tuft, R.; Fogarty, K.; Bao, Z.Z. Notch signaling plays a key role in cardiac cell differentiation. Mech. Dev. 2006. [Google Scholar] [CrossRef]
- Watanabe, Y.; Kokubo, H.; Miyagawa-Tomita, S.; Endo, M.; Igarashi, K.; Aisaki, K.I.; Kanno, J.; Saga, Y. Activation of Notch1 signaling in cardiogenic mesoderm induces abnormal heart morphogenesis in mouse. Development 2006, 133, 1625–1634. [Google Scholar] [CrossRef] [Green Version]
- Gale, N.W.; Dominguez, M.G.; Noguera, I.; Pan, L.; Hughes, V.; Valenzuela, D.M.; Murphy, A.J.; Adams, N.C.; Lin, H.C.; Holash, J.; et al. Haploinsufficiency of delta-like 4 ligand results in embryonic lethality due to major defects in arterial and vascular development. Proc. Natl. Acad. Sci. USA 2004, 101, 15949–15954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcelo, K.L.; Sills, T.M.; Coskun, S.; Vasavada, H.; Sanglikar, S.; Goldie, L.C.; Hirschi, K.K. Hemogenic endothelial cell specification requires c-Kit, notch signaling, and p27-mediated cell-cycle control. Dev. Cell 2013, 27, 504–515. [Google Scholar] [CrossRef] [Green Version]
- Ferdous, A.; Caprioli, A.; Iacovino, M.; Martin, C.M.; Morris, J.; Richardson, J.A.; Latif, S.; Hammer, R.E.; Harvey, R.P.; Olson, E.N.; et al. Nkx2-5 transactivates the Ets-related protein 71 gene and specifies an endothelial/endocardial fate in the developing embryo. Proc. Natl. Acad. Sci. USA 2009, 106, 814–819. [Google Scholar] [CrossRef] [Green Version]
- Reischauer, S.; Stone, O.A.; Villasenor, A.; Chi, N.; Jin, S.W.; Martin, M.; Lee, M.T.; Fukuda, N.; Marass, M.; Witty, A.; et al. Cloche is a bHLH-PAS transcription factor that drives haemato-vascular specification. Nature 2016, 535, 294–298. [Google Scholar] [CrossRef]
- Haraguchi, S.; Kitajima, S.; Takagi, A.; Takeda, H.; Inoue, T.; Saga, Y. Transcriptional regulation of Mesp1 and Mesp2 genes: Differential usage of enhancers during development. Mech. Dev. 2001, 108, 59–69. [Google Scholar] [CrossRef]
- Matsuo, I.; Kuratani, S.; Kimura, C.; Takeda, N.; Aizawa, S. Mouse Otx2 functions in the formation and patterning of rostral head. Genes Dev. 1995, 9, 2646–2658. [Google Scholar] [CrossRef] [Green Version]
- Tellier, A.P.; Archambault, D.; Tremblay, K.D.; Mager, J. The elongation factor Elof1 is required for mammalian gastrulation. PLoS ONE 2019, 14, e0219410. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, W.; Nakanishi, H.; Miyoshi, J.; Mandai, K.; Ishizaki, H.; Tanaka, M.; Togawa, A.; Takahashi, K.; Nishioka, H.; Yoshida, H.; et al. Afadin: A key molecule essential for structural organization of cell- cell junctions of polarized epithelia during embryogenesis. J. Cell Biol. 1999, 146, 1117–1132. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Meyers, E.N.; Lewandoski, M.; Martin, G.R. Targeted disruption of Fgf8 causes failure of cell migration in the gastrulating mouse embryo. Genes Dev. 1999, 13, 1834–1846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, R.; Watt, A.J.; Battle, M.A.; Li, J.; Bondow, B.J.; Duncan, S.A. Loss of both GATA4 and GATA6 blocks cardiac myocyte differentiation and results in acardia in mice. Dev. Biol. 2008, 317, 614–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gannon, M.; Bader, D. Initiation of cardiac differentiation occurs in the absence of anterior endoderm. Development 1995, 121, 2439–2450. [Google Scholar] [CrossRef]
- Palencia-Desai, S.; Kohli, V.; Kang, J.; Chi, N.C.; Black, B.L.; Sumanas, S. Vascular endothelial and endocardial progenitors differentiate as cardiomyocytes in the absence of Etsrp/Etv2 function. Development 2011, 138, 4721–4732. [Google Scholar] [CrossRef] [Green Version]
- Tzahor, E.; Lassar, A.B. Wnt signals from the neural tube block ectopic cardiogenesis. Genes Dev. 2001, 15, 255–260. [Google Scholar] [CrossRef] [Green Version]
- Marvin, M.J.; Di Rocco, G.; Gardiner, A.; Bush, S.M.; Lassar, A.B. Inhibition of Wnt activity induces heart formation from posterior mesoderm. Genes Dev. 2001, 15, 316–327. [Google Scholar] [CrossRef] [Green Version]
- Chestnut, B.; Casie Chetty, S.; Koenig, A.L.; Sumanas, S. Single-cell transcriptomic analysis identifies the conversion of zebrafish Etv2-deficient vascular progenitors into skeletal muscle. Nat. Commun. 2020, 11, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Van Handel, B.; Montel-Hagen, A.; Sasidharan, R.; Nakano, H.; Ferrari, R.; Boogerd, C.J.; Schredelseker, J.; Wang, Y.; Hunter, S.; Org, T.; et al. Scl represses cardiomyogenesis in prospective hemogenic endothelium and endocardium. Cell 2012, 150, 590–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saba, R.; Kitajima, K.; Rainbow, L.; Engert, S.; Uemura, M.; Ishida, H.; Kokkinopoulos, I.; Shintani, Y.; Miyagawa, S.; Kanai, Y.; et al. Endocardium differentiation through Sox17 expression in endocardium precursor cells regulates heart development in mice. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kouskoff, V.; Lacaud, G.; Schwantz, S.; Fehling, H.J.; Keller, G. Sequential development of hematopoietic and cardiac mesoderm during embryonic stem cell differentiation. Proc. Natl. Acad. Sci. USA 2005, 102, 13170–13175. [Google Scholar] [CrossRef] [Green Version]
- Nostro, M.C.; Cheng, X.; Keller, G.M.; Gadue, P. Wnt, Activin, and BMP Signaling Regulate Distinct Stages in the Developmental Pathway from Embryonic Stem Cells to Blood. Cell Stem Cell 2008, 2, 60–71. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Soonpaa, M.H.; Adler, E.D.; Roepke, T.K.; Kattman, S.J.; Kennedy, M.; Henckaerts, E.; Bonham, K.; Abbott, G.W.; Linden, R.M.; et al. Human cardiovascular progenitor cells develop from a KDR+ embryonic-stem-cell-derived population. Nature 2008, 453, 524–528. [Google Scholar] [CrossRef]
- Santini, M.P.; Forte, E.; Harvey, R.P.; Kovacic, J.C. Developmental origin and lineage plasticity of endogenous cardiac stem cells. Development 2016, 143, 1242–1258. [Google Scholar] [CrossRef] [Green Version]
- Van Vliet, P.; Wu, S.M.; Zaffran, S.; Pucéat, M. Early cardiac development: A view from stem cells to embryos. Cardiovasc. Res. 2012, 96, 352–362. [Google Scholar] [CrossRef] [Green Version]
- Rosenquist, G.C.; DeHaan, R.L. Contributions to Embryology; Carnegie Institution of Washington: Washington, DC, USA, 1966; Volume 38, pp. 111–121. [Google Scholar]
- Cui, C.; Cheuvront, T.J.; Lansford, R.D.; Moreno-Rodriguez, R.A.; Schultheiss, T.M.; Rongish, B.J. Dynamic positional fate map of the primary heart-forming region. Dev. Biol. 2009, 332, 212–222. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Sanchez, C.; Garcia-Masa, N.; Gañan, C.M.; Garcia-Martinez, V. Movement and commitment of primitive streak precardiac cells during cardiogenesis. Int. J. Dev. Biol. 2009, 53, 1445–1455. [Google Scholar] [CrossRef] [Green Version]
- Redkar, A.; Montgomery, M.; Litvin, J. Fate map of early avian cardiac progenitor cells. Development 2001, 128, 2269–2279. [Google Scholar] [CrossRef]
- Satin, J.; Fujii, S.; DeHaan, R.L. Development of cardiac beat rate in early chick embryos is regulated by regional cues. Dev. Biol. 1988, 129, 103–113. [Google Scholar] [CrossRef]
- Wu, S.P.; Cheng, C.M.; Lanz, R.B.; Wang, T.; Respress, J.L.; Ather, S.; Chen, W.; Tsai, S.J.; Wehrens, X.H.; Tsai, M.J.; et al. Atrial Identity Is Determined by a COUP-TFII Regulatory Network. Dev. Cell 2013, 25, 417–426. [Google Scholar] [CrossRef] [Green Version]
- Joubin, K.; Stern, C.D. Molecular interactions continuously define the organizer during the cell movements of gastrulation. Cell 1999, 98, 559–571. [Google Scholar] [CrossRef] [Green Version]
- Saito, Y.; Kojima, T.; Takahashi, N. Mab21l2 is essential for embryonic heart and liver development. PLoS ONE 2012, 7, e32991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldessari, D.; Badaloni, A.; Longhi, R.; Zappavigna, V.; Consalez, G.G. MAB21L2, a vertebrate member of the Male-abnormal 21 family, modulates BMP signaling and interacts with SMAD1. BMC Cell Biol. 2004, 5, 48. [Google Scholar] [CrossRef] [Green Version]
- Domínguez, J.N.; Meilhac, S.M.; Bland, Y.S.; Buckingham, M.E.; Brown, N.A. Asymmetric fate of the posterior part of the second heart field results in unexpected left/right contributions to both poles of the heart. Circ. Res. 2012, 111, 1323–1335. [Google Scholar] [CrossRef] [Green Version]
- Zaffran, S.; Kelly, R.G.; Meilhac, S.M.; Buckingham, M.E.; Brown, N.A. Right ventricular myocardium derives from the anterior heart field. Circ. Res. 2004, 95, 261–268. [Google Scholar] [CrossRef] [Green Version]
- Mommersteeg, M.T.; Domínguez, J.N.; Wiese, C.; Norden, J.; De Gier-De Vries, C.; Burch, J.B.; Kispert, A.; Brown, N.A.; Moorman, A.F.; Christoffels, V.M. The sinus venosus progenitors separate and diversify from the first and second heart fields early in development. Cardiovasc. Res. 2010, 87, 92–101. [Google Scholar] [CrossRef] [Green Version]
- Bruneau, B.G.; Nemer, G.; Schmitt, J.P.; Charron, F.; Robitaille, L.; Caron, S.; Conner, D.A.; Gessler, M.; Nemer, M.; Seidman, C.E.; et al. A murine model of Holt-Oram syndrome defines roles of the T-Box transcription factor Tbx5 in cardiogenesis and disease. Cell 2001, 106, 709–721. [Google Scholar] [CrossRef] [Green Version]
- Waddington, C.H. The Strategy of the Genes. A Discussion of Some Aspects of Theoretical Biology. With an Appendix by H. Kacser; CAB Direct: London, UK, 1957; pp. 152–169. [Google Scholar]
- Sáez, M.; Blassberg, R.; Camacho-Aguilar, E.; Siggia, E.D.; Rand, D.A.; Briscoe, J. Statistically derived geometrical landscapes capture principles of decision-making dynamics during cell fate transitions. Cell Syst. 2021. (In Press, Corrected Proof). [Google Scholar] [CrossRef] [PubMed]
- Gilbert, S.F. Developmental Biology, 8th ed.; Sinauer Associates: Sunderland, MA, USA, 2006; p. 751. [Google Scholar]
- Guignard, L.; Fiúza, U.M.; Leggio, B.; Laussu, J.; Faure, E.; Michelin, G.; Biasuz, K.; Hufnagel, L.; Malandain, G.; Godin, C.; et al. Contact area-dependent cell communication and the morphological invariance of ascidian embryogenesis. Science 2020, 369, eaar5663. [Google Scholar] [CrossRef] [PubMed]
- Wiegner, O.; Schierenberg, E. Regulative development in a nematode embryo: A hierarchy of cell fate transformations. Dev. Biol. 1999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossant, J. Stem Cells and Early Lineage Development. Cell 2008, 132, 527–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moorman, A.F.; Christoffels, V.M.; Anderson, R.H.; Van Den Hoff, M.J. The heart-forming fields: One or multiple? Philos. Trans. R. Soc. Biol. Sci. 2007, 362, 1257–1265. [Google Scholar] [CrossRef] [Green Version]
- Cai, C.L.; Liang, X.; Shi, Y.; Chu, P.H.; Pfaff, S.L.; Chen, J.; Evans, S. Isl1 identifies a cardiac progenitor population that proliferates prior to differentiation and contributes a majority of cells to the heart. Dev. Cell 2003, 5, 877–889. [Google Scholar] [CrossRef] [Green Version]
- Noden, D.M.; Poelmann, R.E.; Gittenberger-de Groot, A.C. Cell origins and tissue boundaries during outflow tract development. Trends Cardiovasc. Med. 1995, 5, 69–75. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Model | Methodology | Stage | Location | N | Progeny (% from Total) | Reference | |||
|---|---|---|---|---|---|---|---|---|---|
| CM | EC | CM + EC | CM + EC + Non Cardiac | ||||||
| Zebrafish | Single-cell dextran microinjection | Early blastula (∼2.75 hpf) | lateral-marginal zone | 29 | 4 | 10 | 21 | 65 | [20] |
| Zebrafish | Single-cell dextran microinjection | Midblastula (∼3 hpf ) | lateral-marginal zone | 41 | 18 | 7 | 0 | 75 | [20] |
| Zebrafish | 2–3 cells fluorescein activation | 40% epiboly (∼5 hpf) | ventral-marginal zone | 69 | 44 | 52 | 4 | ND | [21] |
| Zebrafish | Kaede photoconversion | 14-somite (∼18 hpf) | anterior lateral plate mesoderm | 39 | 27 | 0 | 2 | ND | [22] |
| Chicken | Replication-defective retrovirus | HH3 (∼14 h) | anterior lateral plate mesoderm | 82 | 95 | 5 | 0 | ND | [23] |
| Chicken | Replication-defective retrovirus | HH4 (∼18 h) | rostral portion primitive streak | 36 | 55 * | 45 * | 0 | ND | [23] |
| Mouse | Doxycyclin induced transgene expression | Early primitive streak (∼E6.5) ** | Mesp1-expressing cells | 13 | 85 | 15 | 0 | ND | [24] |
| Mouse | Doxycyclin induced transgene expression | Late primitive streak (∼E7.0) ** | Mesp1-expressing cells | 6 | 100 | 0 | 0 | ND | [24] |
| Mouse | Doxycyclin induced transgene expression | Late bud (∼E7.5) ** | Mesp1-expressing cells | 17 | 70 | 6 | 24 *** | ND | [24] |
| Resource | Stage | Selection (N Cells) | Method | Depth * | Data | Reference |
|---|---|---|---|---|---|---|
| scRNAseq | ES to LHF ∼E6.5, E7.5, E7.75 | ∼E6.5 epiblast (501) ∼E7.5 Flk1+ (704) | Smart-seq2 | ∼1 × 106 reads/cell | link list 1 | [25] |
| scRNAseq | ∼E6.75, E7.25 | Mesp1+ ∼E6.75 (83 WT 85 Mesp1−/−) ∼E7.25 (173) | Smart-seq2 | ∼1 × 106 reads/cell ** | link list 2 | [26] |
| scRNAseq | ∼E8.25 | Whole embryo (19,396) | 10× genomics | ∼2 × 104 UMIs/cell | link list 3 | [27] |
| scRNAseq | ES to 7 s ∼E6.5, E6.75, E7.25, E7.5, E7.75, E8.0, E8.25, E8.5 | Whole embryo (116,312) Per stage: supplementary | 10× genomics | ∼2 × 104 UMIs/cell | link list 4 | [28] |
| scRNAseq | 4 s, 8 s, 21 s ∼E7.75, E8.25, E9.25 | Dissected cardiac region E7.75 (4326 WT 3535 Hand2−/−) E8.25 (5664 WT 4112 Hand2−/−) E9.25 (11,376 WT) | 10× genomics | ∼2 × 104 UMIs/cell | link list 5 | [29] |
| scRNAseq | LHF, 8 s, 13 s, 20 s ∼E7.75, E8.25, E8.75, E9.25 | Nkx2-5+ (690), Isl1+ (640) Per stage: supplementary 1 | Modified Smart-seq2 | ∼1 × 106 reads/cell | link list 6 | [30] |
| scRNAseq | Pre-Streak stages ∼E5.25, E5.5, E6.25, E6.5 | Whole embryo E5.25 (331), E5.5 (269) E6.25 (321), E6.5 (803) | Smart-seq2 | ∼1 × 106 reads/cell ** | link list 7 | [31] |
| scRNAseq | ∼E7.5, E8.5, E9.5 | Nkx2-5+ E7.5 (61), E8.5 (58) E9.5 (81) Isl1+ E7.5 (30), E8.5 (167) E9.5 (348 WT 50 Isl1−/−) | Smart-seq2 | ∼1 × 106 reads/cell ** | link list 8 | [32] |
| scRNAseq | PrS to Presomitic ∼E6.5 to E8.25 | Whole embryo (33,700 from 153 embryos) | MARS-seq | ∼4 × 103 UMIs/cell | link list 9 | [33] |
| scRNAseq | LHF to 4 s ∼E7.75 to E8.25 | dissected cardiac region (3105) | Smart-seq2 | ∼1 × 106 reads/cell ** | link list 10 | [18] |
| scRNAseq | 0B to somite stage ∼E7.25 to E8.25 | Mesp1+ (9072) | 10× genomics | 60,450 UMIs/cell | link list 11 | [19] |
| snATACseq | ∼E8.5, E9.5 | Isl1+ (695) | [34] | ∼1.5 × 104 reads/nucleus * | link list 12 | [32] |
| snATACseq | ∼E8.25 | Whole embryo (19,453) | [35] | ∼2 × 104 reads/nucleus * | link list 13 | [28] |
| scNMTseq | ∼4.5, E5.5, E6.5, E7.5 | Whole embryo (856) | [36] | ∼1 × 106 reads/cell *** | link list 14 | [37] |
| Live imaging | LB to 4 s ∼E7.5 to E8.5 | Cardiac region: 4 embryos Tdtomato mosaic Nkx2-5:GFP | Two-photon microscopy | 10 min 5 m | NA | [13] |
| Live imaging | LS to 2 s ∼E7.0 to E8.25 | 4 Whole embryos, H2B:eGFP | Adaptative light-sheet microscopy | 4 min 2 m | link list 15 | [38] |
| Live imaging | MS to LB ∼E6.75 to 7.5 | 4 Whole embryo: T-cre mT/mG mosaic | Two-photon microscopy | 20 min 3 m | NA | [39] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sendra, M.; Domínguez, J.N.; Torres, M.; Ocaña, O.H. Dissecting the Complexity of Early Heart Progenitor Cells. J. Cardiovasc. Dev. Dis. 2022, 9, 5. https://doi.org/10.3390/jcdd9010005
Sendra M, Domínguez JN, Torres M, Ocaña OH. Dissecting the Complexity of Early Heart Progenitor Cells. Journal of Cardiovascular Development and Disease. 2022; 9(1):5. https://doi.org/10.3390/jcdd9010005
Chicago/Turabian StyleSendra, Miquel, Jorge N. Domínguez, Miguel Torres, and Oscar H. Ocaña. 2022. "Dissecting the Complexity of Early Heart Progenitor Cells" Journal of Cardiovascular Development and Disease 9, no. 1: 5. https://doi.org/10.3390/jcdd9010005
APA StyleSendra, M., Domínguez, J. N., Torres, M., & Ocaña, O. H. (2022). Dissecting the Complexity of Early Heart Progenitor Cells. Journal of Cardiovascular Development and Disease, 9(1), 5. https://doi.org/10.3390/jcdd9010005