
High Soluble Endoglin Levels Affect Aortic Vascular Function during Mice Aging

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Biochemical Analysis
2.3. Histological Analysis
2.4. ELISA
2.5. Functional Analysis of Vascular Reactivity Ex Vivo
2.6. Western Blot Analysis
2.7. Statistical Analysis
3. Results
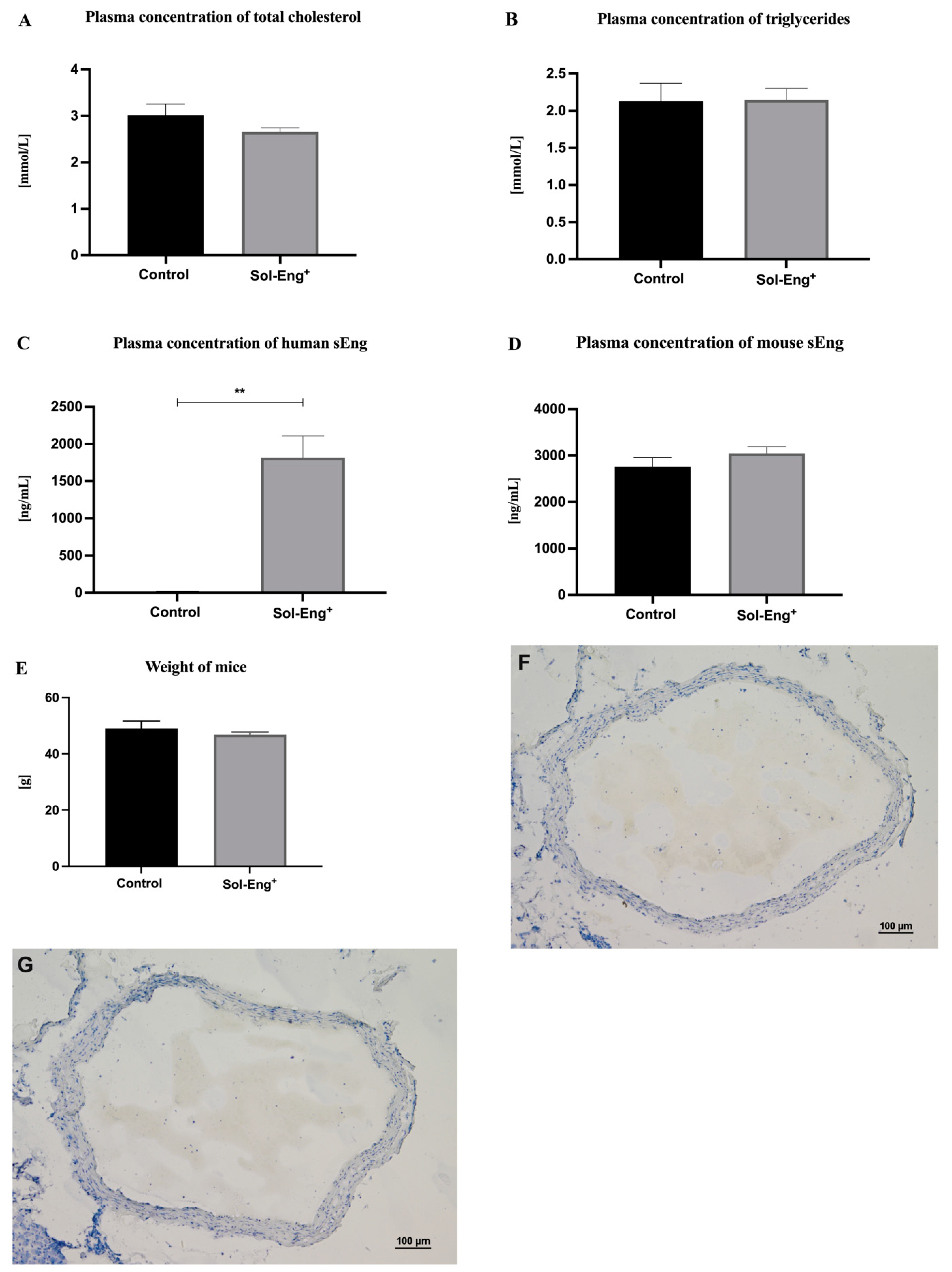
3.1. Lipid Spectrum, Human and Mouse sEng Levels in Plasma
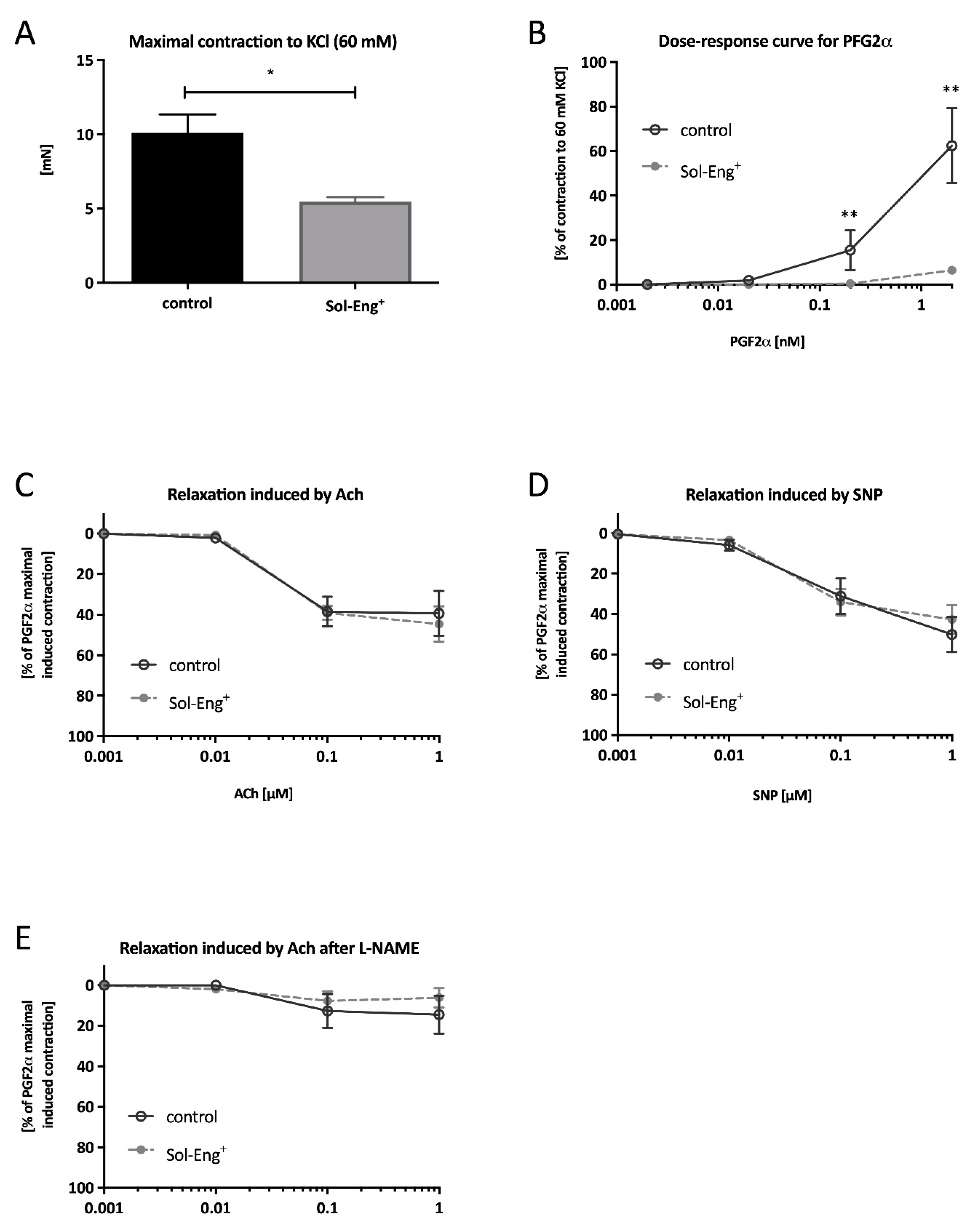
3.2. Impaired Vascular Contractility in Aortas of Sol-Eng+ Mice
3.3. High sEng Levels Do Not Affect the Expression of Biomarkers of Endothelial Dysfunction
3.4. High sEng Levels Inhibit Membrane Endoglin Downstream Signaling
3.5. High Levels of sEng Affect the Expression of Markers Related to Vascular Contractility
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bot, P.T.; Hoefer, I.E.; Sluijter, J.P.; van Vliet, P.; Smits, A.M.; Lebrin, F.; Moll, F.; de Vries, J.P.; Doevendans, P.; Piek, J.J.; et al. Increased expression of the transforming growth factor-beta signaling pathway, endoglin, and early growth response-1 in stable plaques. Stroke 2009, 40, 439–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- St-Jacques, S.; Cymerman, U.; Pece, N.; Letarte, M. Molecular characterization and in situ localization of murine endoglin reveal that it is a transforming growth factor-beta binding protein of endothelial and stromal cells. Endocrinology 1994, 134, 2645–2657. [Google Scholar] [CrossRef] [PubMed]
- Meurer, S.; Wimmer, A.E.; Leur, E.V.; Weiskirchen, R. Endoglin Trafficking/Exosomal Targeting in Liver Cells Depends on N-Glycosylation. Cells 2019, 8, 997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lastres, P.; Bellon, T.; Cabanas, C.; Sanchez-Madrid, F.; Acevedo, A.; Gougos, A.; Letarte, M.; Bernabeu, C. Regulated expression on human macrophages of endoglin, an Arg-Gly-Asp-containing surface antigen. Eur. J. Immunol. 1992, 22, 393–397. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Nunez, M.; Munoz-Felix, J.M.; Lopez-Novoa, J.M. The ALK-1/Smad1 pathway in cardiovascular physiopathology. A new target for therapy? Biochim. Biophys. Acta 2013, 1832, 1492–1510. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Vanhoutte, P.M.; Leung, S.W. Vascular nitric oxide: Beyond eNOS. J. Pharm. Sci. 2015, 129, 83–94. [Google Scholar] [CrossRef] [Green Version]
- Palmer, R.M.; Ashton, D.S.; Moncada, S. Vascular endothelial cells synthesize nitric oxide from l-arginine. Nature 1988, 333, 664–666. [Google Scholar] [CrossRef] [PubMed]
- Moncada, S.; Higgs, E.A. Nitric oxide and the vascular endothelium. Handb Exp. Pharm. 2006, 176, 213–254. [Google Scholar] [CrossRef]
- Lloyd-Jones, D.M.; Bloch, K.D. The vascular biology of nitric oxide and its role in atherogenesis. Annu. Rev. Med. 1996, 47, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Nachtigal, P.; Zemankova Vecerova, L.; Rathouska, J.; Strasky, Z. The role of endoglin in atherosclerosis. Atherosclerosis 2012, 224, 4–11. [Google Scholar] [CrossRef]
- Zhang, X.H.; Zhang, H.Y.; Lu, S.; Jiang, L.L.; Wu, J.; Yang, Y.L.; Zhang, S.A. MMP-14 aggravates onset of severe preeclampsia by mediating soluble endoglin release. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 1209–1215. [Google Scholar]
- Vitverova, B.; Blazickova, K.; Najmanova, I.; Vicen, M.; Hyspler, R.; Dolezelova, E.; Nemeckova, I.; Tebbens, J.D.; Bernabeu, C.; Pericacho, M.; et al. Soluble endoglin and hypercholesterolemia aggravate endothelial and vessel wall dysfunction in mouse aorta. Atherosclerosis 2018, 271, 15–25. [Google Scholar] [CrossRef]
- Blazquez-Medela, A.M.; Garcia-Ortiz, L.; Gomez-Marcos, M.A.; Recio-Rodriguez, J.I.; Sanchez-Rodriguez, A.; Lopez-Novoa, J.M.; Martinez-Salgado, C. Increased plasma soluble endoglin levels as an indicator of cardiovascular alterations in hypertensive and diabetic patients. BMC Med. 2010, 8, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallardo-Vara, E.; Gamella-Pozuelo, L.; Perez-Roque, L.; Bartha, J.L.; Garcia-Palmero, I.; Casal, J.I.; Lopez-Novoa, J.M.; Pericacho, M.; Bernabeu, C. Potential Role of Circulating Endoglin in Hypertension via the Upregulated Expression of BMP4. Cells 2020, 9, 988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, W.T.; Dong, D.L. Bone morphogenetic protein-4: A novel therapeutic target for pathological cardiac hypertrophy/heart failure. Heart Fail. Rev. 2014, 19, 781–788. [Google Scholar] [CrossRef] [PubMed]
- Nemeckova, I.; Serwadczak, A.; Oujo, B.; Jezkova, K.; Rathouska, J.; Fikrova, P.; Varejckova, M.; Bernabeu, C.; Lopez-Novoa, J.M.; Chlopicki, S.; et al. High soluble endoglin levels do not induce endothelial dysfunction in mouse aorta. PLoS ONE 2015, 10, e0119665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jezkova, K.; Rathouska, J.; Nemeckova, I.; Fikrova, P.; Dolezelova, E.; Varejckova, M.; Vitverova, B.; Tysonova, K.; Serwadczak, A.; Buczek, E.; et al. High Levels of Soluble Endoglin Induce a Proinflammatory and Oxidative-Stress Phenotype Associated with Preserved NO-Dependent Vasodilatation in Aortas from Mice Fed a High-Fat Diet. J. Vasc. Res. 2016, 53, 149–162. [Google Scholar] [CrossRef] [PubMed]
- Vanhooren, V.; Libert, C. The mouse as a model organism in aging research: Usefulness, pitfalls and possibilities. Ageing Res. Rev. 2013, 12, 8–21. [Google Scholar] [CrossRef]
- Ackert-Bicknell, C.L.; Anderson, L.C.; Sheehan, S.; Hill, W.G.; Chang, B.; Churchill, G.A.; Chesler, E.J.; Korstanje, R.; Peters, L.L. Aging Research Using Mouse Models. Curr. Protoc. Mouse Biol. 2015, 5, 95–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harkema, L.; Youssef, S.A.; de Bruin, A. Pathology of Mouse Models of Accelerated Aging. Vet. Pathol. 2016, 53, 366–389. [Google Scholar] [CrossRef]
- Jackson, S.J.; Andrews, N.; Ball, D.; Bellantuono, I.; Gray, J.; Hachoumi, L.; Holmes, A.; Latcham, J.; Petrie, A.; Potter, P.; et al. Does age matter? The impact of rodent age on study outcomes. Lab. Anim. 2017, 51, 160–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valbuena-Diez, A.C.; Blanco, F.J.; Oujo, B.; Langa, C.; Gonzalez-Nunez, M.; Llano, E.; Pendas, A.M.; Diaz, M.; Castrillo, A.; Lopez-Novoa, J.M.; et al. Oxysterol-induced soluble endoglin release and its involvement in hypertension. Circulation 2012, 126, 2612–2624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vicen, M.; Igreja Sa, I.C.; Tripska, K.; Vitverova, B.; Najmanova, I.; Eissazadeh, S.; Micuda, S.; Nachtigal, P. Membrane and soluble endoglin role in cardiovascular and metabolic disorders related to metabolic syndrome. Cell Mol. Life Sci. 2020, 78, 2405–2418. [Google Scholar] [CrossRef]
- Dolezelova, E.; Sa, I.C.I.; Prasnicka, A.; Hroch, M.; Hyspler, R.; Ticha, A.; Lastuvkova, H.; Cermanova, J.; Pericacho, M.; Visek, J.; et al. High soluble endoglin levels regulate cholesterol homeostasis and bile acids turnover in the liver of transgenic mice. Life Sci. 2019, 232, 116643. [Google Scholar] [CrossRef]
- Igreja Sa, I.C.; Tripska, K.; Hroch, M.; Hyspler, R.; Ticha, A.; Lastuvkova, H.; Schreiberova, J.; Dolezelova, E.; Eissazadeh, S.; Vitverova, B.; et al. Soluble Endoglin as a Potential Biomarker of Nonalcoholic Steatohepatitis (NASH) Development, Participating in Aggravation of NASH-Related Changes in Mouse Liver. Int. J. Mol. Sci. 2020, 21, 9021. [Google Scholar] [CrossRef] [PubMed]
- Venkatesha, S.; Toporsian, M.; Lam, C.; Hanai, J.; Mammoto, T.; Kim, Y.M.; Bdolah, Y.; Lim, K.H.; Yuan, H.T.; Libermann, T.A.; et al. Soluble endoglin contributes to the pathogenesis of preeclampsia. Nat. Med. 2006, 12, 642–649. [Google Scholar] [CrossRef]
- Varejckova, M.; Gallardo-Vara, E.; Vicen, M.; Vitverova, B.; Fikrova, P.; Dolezelova, E.; Rathouska, J.; Prasnicka, A.; Blazickova, K.; Micuda, S.; et al. Soluble endoglin modulates the pro-inflammatory mediators NF-kappaB and IL-6 in cultured human endothelial cells. Life Sci. 2017, 175, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Perez-Roque, L.; Nunez-Gomez, E.; Rodriguez-Barbero, A.; Bernabeu, C.; Lopez-Novoa, J.M.; Pericacho, M. Pregnancy-Induced High Plasma Levels of Soluble Endoglin in Mice Lead to Preeclampsia Symptoms and Placental Abnormalities. Int. J. Mol. Sci. 2020, 22, 165. [Google Scholar] [CrossRef]
- Shao, C.; Wang, J.; Tian, J.; Tang, Y.D. Coronary Artery Disease: From Mechanism to Clinical Practice. Adv. Exp. Med. Biol. 2020, 1177, 1–36. [Google Scholar] [CrossRef] [PubMed]
- Ratz, P.H.; Berg, K.M.; Urban, N.H.; Miner, A.S. Regulation of smooth muscle calcium sensitivity: KCl as a calcium-sensitizing stimulus. Am. J. Physiol. Cell Physiol. 2005, 288, C769–C783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Himpens, B.; Missiaen, L.; Casteels, R. Ca2+ homeostasis in vascular smooth muscle. J. Vasc. Res. 1995, 32, 207–219. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, S.; Koh, E.; Kim, S.; Morita, R.; Fukuo, K.; Ogihara, T. Effects of prostaglandin F2 alpha on the mobilization of cytosolic free calcium in vascular smooth muscle cells and on the tension of aortic strips from rats. Am. J. Hypertens. 1990, 3, 241S–244S. [Google Scholar] [CrossRef]
- Kishi, H.; Ye, L.H.; Nakamura, A.; Okagaki, T.; Iwata, A.; Tanaka, T.; Kohama, K. Structure and function of smooth muscle myosin light chain kinase. Adv. Exp. Med. Biol. 1998, 453, 229–234. [Google Scholar] [CrossRef]
- Stull, J.T.; Tansey, M.G.; Tang, D.C.; Word, R.A.; Kamm, K.E. Phosphorylation of myosin light chain kinase: A cellular mechanism for Ca2+ desensitization. Mol. Cell Biochem. 1993, 127-128, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Eglen, R.M.; Whiting, R.L. Heterogeneity of vascular muscarinic receptors. J. Auton. Pharm. 1990, 10, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Tangsucharit, P.; Takatori, S.; Zamami, Y.; Goda, M.; Pakdeechote, P.; Kawasaki, H.; Takayama, F. Muscarinic acetylcholine receptor M1 and M3 subtypes mediate acetylcholine-induced endothelium-independent vasodilatation in rat mesenteric arteries. J. Pharm. Sci. 2016, 130, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Vanhoutte, P.M.; De Mey, J. Control of vascular smooth muscle function by the endothelial cells. Gen. Pharm. 1983, 14, 39–41. [Google Scholar] [CrossRef]
- Vicen, M.; Vitverova, B.; Havelek, R.; Blazickova, K.; Machacek, M.; Rathouska, J.; Najmanova, I.; Dolezelova, E.; Prasnicka, A.; Sternak, M.; et al. Regulation and role of endoglin in cholesterol-induced endothelial and vascular dysfunction in vivo and in vitro. FASEB J. 2019, 33, 6099–6114. [Google Scholar] [CrossRef]
- Santibanez, J.F.; Letamendia, A.; Perez-Barriocanal, F.; Silvestri, C.; Saura, M.; Vary, C.P.; Lopez-Novoa, J.M.; Attisano, L.; Bernabeu, C. Endoglin increases eNOS expression by modulating Smad2 protein levels and Smad2-dependent TGF-beta signaling. J. Cell Physiol. 2007, 210, 456–468. [Google Scholar] [CrossRef] [PubMed]
- Toporsian, M.; Gros, R.; Kabir, M.G.; Vera, S.; Govindaraju, K.; Eidelman, D.H.; Husain, M.; Letarte, M. A role for endoglin in coupling eNOS activity and regulating vascular tone revealed in hereditary hemorrhagic telangiectasia. Circ. Res. 2005, 96, 684–692. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.A.; Druhan, L.J.; Varadharaj, S.; Chen, Y.R.; Zweier, J.L. Phosphorylation of endothelial nitric-oxide synthase regulates superoxide generation from the enzyme. J. Biol. Chem. 2008, 283, 27038–27047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goumans, M.J.; Valdimarsdottir, G.; Itoh, S.; Rosendahl, A.; Sideras, P.; ten Dijke, P. Balancing the activation state of the endothelium via two distinct TGF-beta type I receptors. EMBO J. 2002, 21, 1743–1753. [Google Scholar] [CrossRef]
- Galkina, E.; Ley, K. Vascular adhesion molecules in atherosclerosis. Arter. Thromb. Vasc. Biol. 2007, 27, 2292–2301. [Google Scholar] [CrossRef] [PubMed]
- Sumagin, R.; Lomakina, E.; Sarelius, I.H. Leukocyte-endothelial cell interactions are linked to vascular permeability via ICAM-1-mediated signaling. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, H969–H977. [Google Scholar] [CrossRef] [Green Version]
- Habas, K.; Shang, L. Alterations in intercellular adhesion molecule 1 (ICAM-1) and vascular cell adhesion molecule 1 (VCAM-1) in human endothelial cells. Tissue Cell 2018, 54, 139–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, J.K. Linking endothelial dysfunction with endothelial cell activation. J. Clin. Investig. 2013, 123, 540–541. [Google Scholar] [CrossRef] [PubMed]
- Alt, A.; Miguel-Romero, L.; Donderis, J.; Aristorena, M.; Blanco, F.J.; Round, A.; Rubio, V.; Bernabeu, C.; Marina, A. Structural and functional insights into endoglin ligand recognition and binding. PLoS ONE 2012, 7, e29948. [Google Scholar] [CrossRef] [Green Version]
- Van Le, B.; Franke, D.; Svergun, D.I.; Han, T.; Hwang, H.Y.; Kim, K.K. Structural and functional characterization of soluble endoglin receptor. Biochem. Biophys. Res. Commun. 2009, 383, 386–391. [Google Scholar] [CrossRef]
- Rossi, E.; Sanz-Rodriguez, F.; Eleno, N.; Duwell, A.; Blanco, F.J.; Langa, C.; Botella, L.M.; Cabanas, C.; Lopez-Novoa, J.M.; Bernabeu, C. Endothelial endoglin is involved in inflammation: Role in leukocyte adhesion and transmigration. Blood 2013, 121, 403–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, E.; Smadja, D.M.; Boscolo, E.; Langa, C.; Arevalo, M.A.; Pericacho, M.; Gamella-Pozuelo, L.; Kauskot, A.; Botella, L.M.; Gaussem, P.; et al. Endoglin regulates mural cell adhesion in the circulatory system. Cell Mol. Life Sci. 2016, 73, 1715–1739. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primary Antibody | Secondary Antibody | |||
|---|---|---|---|---|
| Protein | Source | Dilution | Source | Dilution |
| Endoglin | (sc-19793) (Santa Cruz Biotechnology, Inc. Dallas, TX, USA | 1:200 | Sigma-Aldrich (A5420) | 1:5000 |
| eNOS | (sc-654) (Santa Cruz Biotechnology, Inc. Dallas, TX, USA | 1:200 | Abcam (ab6112) | 1:2000 |
| p-eNOS | (sc-21871-R) (Santa Cruz Biotechnology, Inc. Dallas, TX, USA | 1:200 | Abcam (ab6112) | 1:2000 |
| ID1 | (ab134163), (Abcam, Cambridge, UK) | 1:1000 | Abcam (ab6112) | 1:2000 |
| VCAM-1 | (32653S), (Cell Signaling Technology, Inc., Danvers, MA, USA) | 1:1000 | Abcam (ab6112) | 1:2000 |
| MLCK | (ab76092), (Abcam, Cambridge, UK) | 1:200 | Abcam (ab6112) | 1:1000 |
| p-MLCK | (44-1085G), (Invitrogen, Waltham, MA, USA) | 1:1000 | Abcam (ab6112) | 1:2000 |
| GAPDH | (G8795) (Sigma-Aldrich, St. Louis, MO, USA) | 1:10,000 | Sigma-Aldrich (A9917) | 1:20,000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nejmanová, I.; Vitverová, B.; Eissazadeh, S.; Tripská, K.; Igreja Sa, I.C.; Hyšpler, R.; Němečkova, I.; Pericacho, M.; Nachtigal, P. High Soluble Endoglin Levels Affect Aortic Vascular Function during Mice Aging. J. Cardiovasc. Dev. Dis. 2021, 8, 173. https://doi.org/10.3390/jcdd8120173
Nejmanová I, Vitverová B, Eissazadeh S, Tripská K, Igreja Sa IC, Hyšpler R, Němečkova I, Pericacho M, Nachtigal P. High Soluble Endoglin Levels Affect Aortic Vascular Function during Mice Aging. Journal of Cardiovascular Development and Disease. 2021; 8(12):173. https://doi.org/10.3390/jcdd8120173
Chicago/Turabian StyleNejmanová, Iveta, Barbora Vitverová, Samira Eissazadeh, Katarina Tripská, Ivone Cristina Igreja Sa, Radomír Hyšpler, Ivana Němečkova, Miguel Pericacho, and Petr Nachtigal. 2021. "High Soluble Endoglin Levels Affect Aortic Vascular Function during Mice Aging" Journal of Cardiovascular Development and Disease 8, no. 12: 173. https://doi.org/10.3390/jcdd8120173
APA StyleNejmanová, I., Vitverová, B., Eissazadeh, S., Tripská, K., Igreja Sa, I. C., Hyšpler, R., Němečkova, I., Pericacho, M., & Nachtigal, P. (2021). High Soluble Endoglin Levels Affect Aortic Vascular Function during Mice Aging. Journal of Cardiovascular Development and Disease, 8(12), 173. https://doi.org/10.3390/jcdd8120173