
Neonatal Deletion of Hand1 and Hand2 within Murine Cardiac Conduction System Reveals a Novel Role for HAND2 in Rhythm Homeostasis

 , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mouse Lines and Breeding
2.2. Surface ECG Recording and Analysis
2.3. Optical Mapping
2.4. Statistical Analysis
3. Results
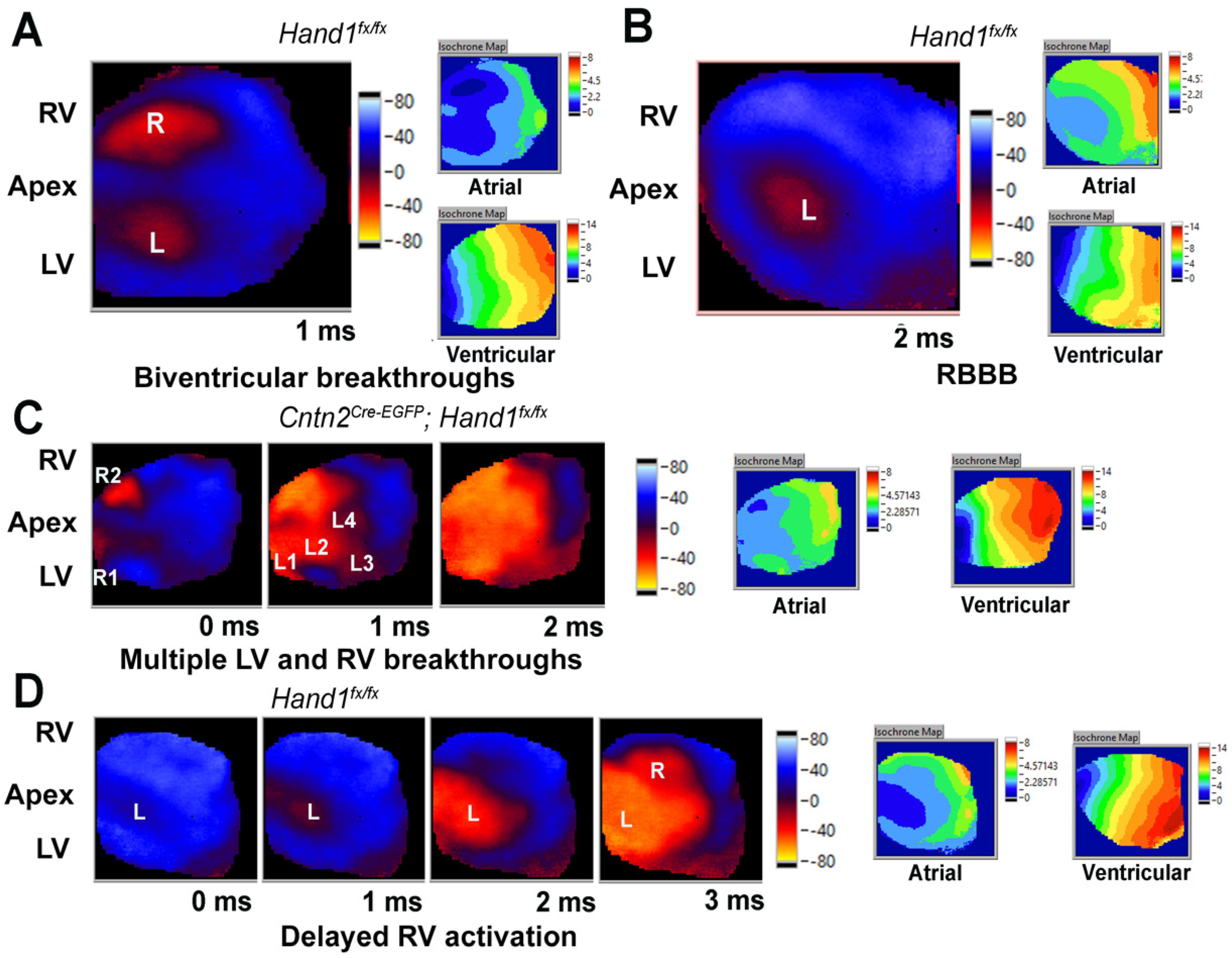
3.1. Neonatal Deletion of Hand1 Does Not Observably alter CCS Function
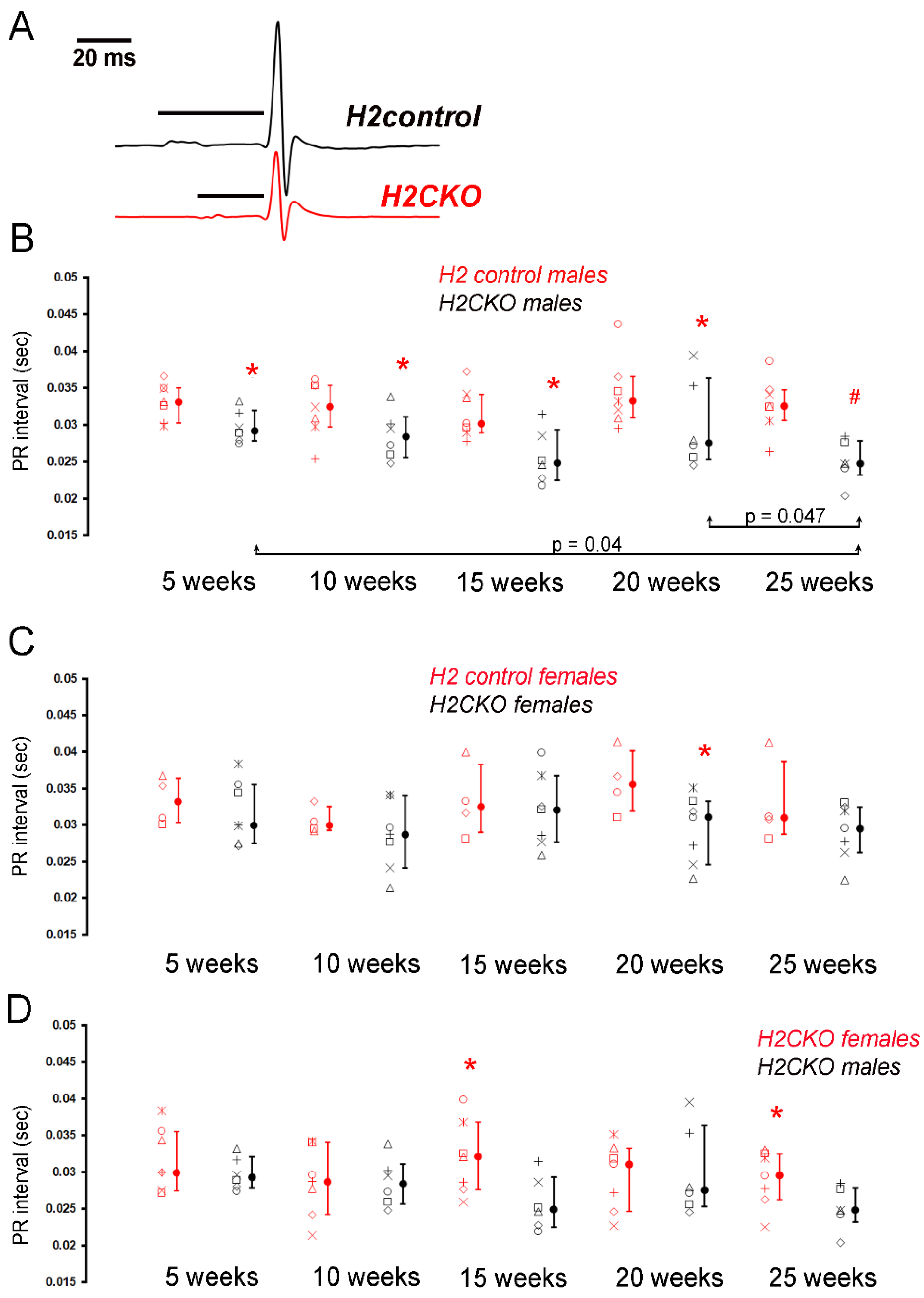
3.2. Neonatal Deletion of Hand2 Results in Sex-Dependent Effects on CCS Function
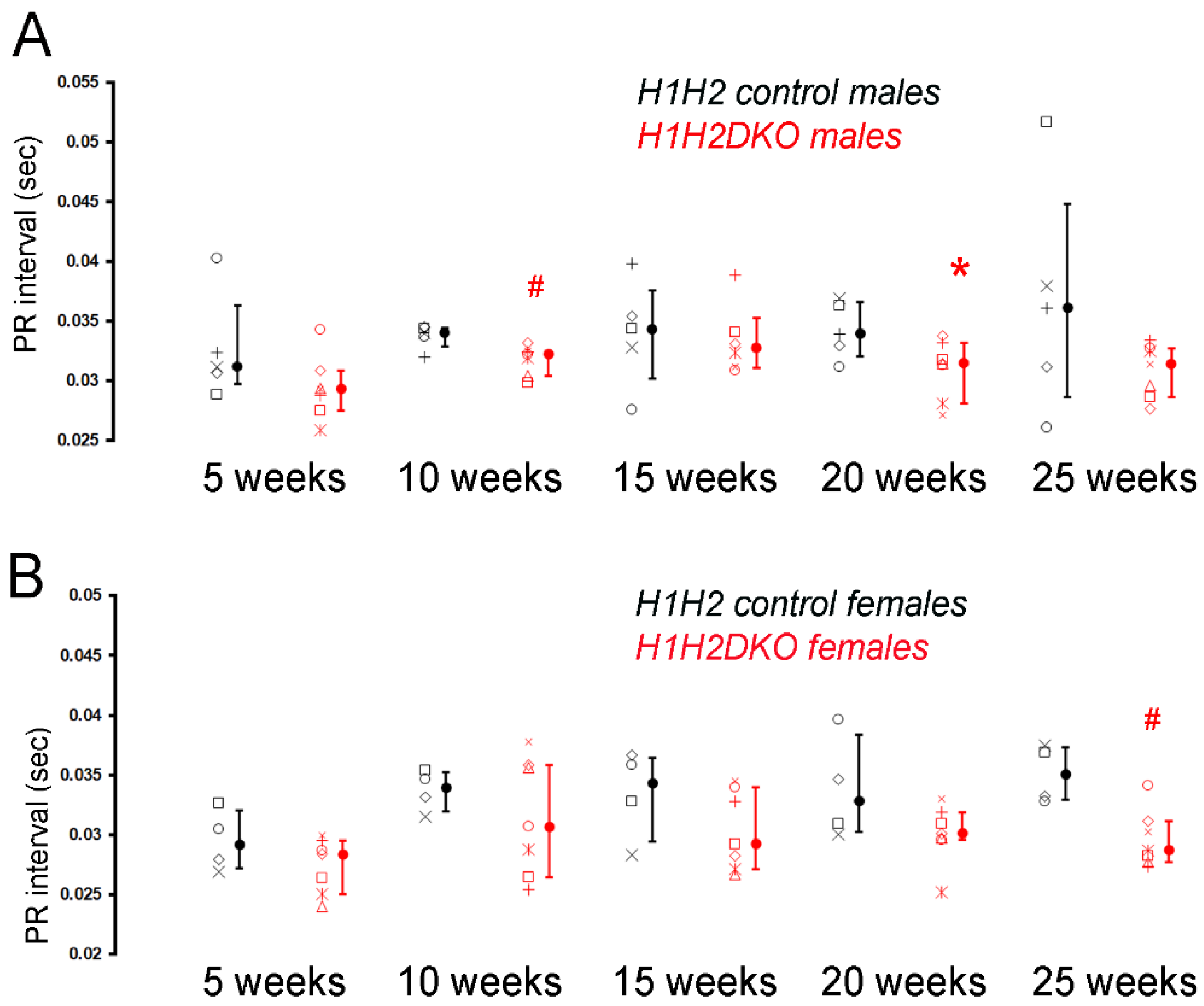
3.3. Neonatal Deletion of Hand1 and Hand2 Results in an Intermediate CCS Phenotype
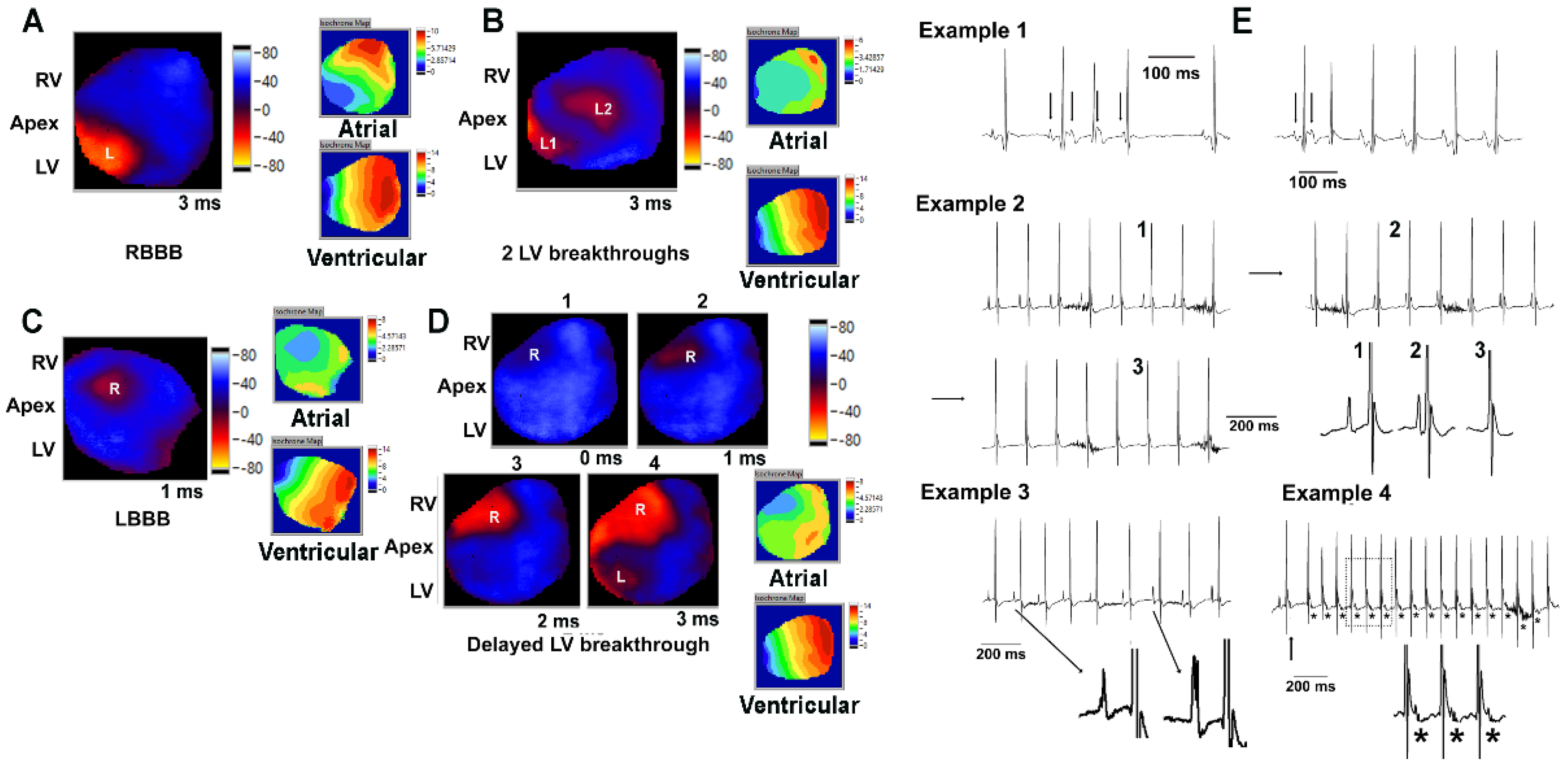
3.4. Embryonic Deletion of Hand2 within the LV Results in Increase in PR Interval and Arrhythmias
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Padala, S.K.; Cabrera, J.A.; Ellenbogen, K.A. Anatomy of the cardiac conduction system. Pacing Clin. Electrophysiol. 2021, 44, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Nygren, A.; Clark, R.B.; Belke, D.D.; Kondo, C.; Giles, W.R.; Witkowski, F.X. Voltage-sensitive dye mapping of activation and conduction in adult mouse hearts. Ann. Biomed. Eng. 2000, 28, 958–967. [Google Scholar] [CrossRef] [PubMed]
- Tamaddon, H.S.; Vaidya, D.; Simon, A.M.; Paul, D.L.; Jalife, J.; Morley, G.E. High-resolution optical mapping of the right bundle branch in connexin40 knockout mice reveals slow conduction in the specialized conduction system. Circ. Res. 2000, 87, 929–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miquerol, L.; Beyer, S.; Kelly, R.G. Establishment of the mouse ventricular conduction system. Cardiovasc. Res. 2011, 91, 232–242. [Google Scholar] [CrossRef] [Green Version]
- Rubart, M.; Zipes, D.P. Mechanisms of sudden cardiac death. J. Clin. Investig. 2005, 115, 2305–2315. [Google Scholar] [CrossRef] [Green Version]
- Kelly, R.G.; Buckingham, M.E.; Moorman, A.F. Heart fields and cardiac morphogenesis. Cold Spring Harb. Perspect. Med. 2014, 4, a015750. [Google Scholar] [CrossRef] [Green Version]
- van Weerd, J.H.; Christoffels, V.M. The formation and function of the cardiac conduction system. Development 2016, 143, 197–210. [Google Scholar] [CrossRef] [Green Version]
- Christoffels, V.M.; Moorman, A.F. Development of the cardiac conduction system: Why are some regions of the heart more arrhythmogenic than others? Circ. Arrhythm. Electrophysiol. 2009, 2, 195–207. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharyya, S.; Munshi, N.V. Development of the cardiac conduction system. Cold Spring Harb. Perspect. Biol. 2020, 12, a037408. [Google Scholar] [CrossRef]
- George, R.M.; Firulli, A.B. Hand factors in cardiac development. Anat. Rec. 2019, 302, 101–107. [Google Scholar] [CrossRef] [Green Version]
- Breckenridge, R.A.; Zuberi, Z.; Gomes, J.; Orford, R.; Dupays, L.; Felkin, L.E.; Clark, J.E.; Magee, A.I.; Ehler, E.; Birks, E.J.; et al. Overexpression of the transcription factor Hand1 causes predisposition towards arrhythmia in mice. J. Mol. Cell Cardiol. 2009, 47, 133–141. [Google Scholar] [CrossRef]
- McFadden, D.G.; Barbosa, A.C.; Richardson, J.A.; Schneider, M.D.; Srivastava, D.; Olson, E.N. The Hand1 and Hand2 transcription factors regulate expansion of the embryonic cardiac ventricles in a gene dosage-dependent manner. Development 2005, 132, 189–201. [Google Scholar] [CrossRef] [Green Version]
- Firulli, B.A.; George, R.M.; Harkin, J.; Toolan, K.P.; Gao, H.; Liu, Y.; Zhang, W.; Field, L.J.; Liu, Y.; Shou, W.; et al. HAND1 loss-of-function within the embryonic myocardium reveals survivable congenital cardiac defects and adult heart failure. Cardiovasc. Res. 2020, 116, 605–618. [Google Scholar] [CrossRef]
- Fernandez-Perez, A.; Sathe, A.A.; Bhakta, M.; Leggett, K.; Xing, C.; Munshi, N.V. Hand2 selectively reorganizes chromatin accessibility to induce pacemaker-like transcriptional reprogramming. Cell Rep. 2019, 27, 2354–2369 e7. [Google Scholar] [CrossRef] [Green Version]
- Vincentz, J.W.; Firulli, B.A.; Toolan, K.P.; Arking, D.E.; Sotoodehnia, N.; Wan, J.; Chen, P.S.; De Gier-De Vries, C.; Christoffels, V.M.; Rubart-Von Der Lohe, M.; et al. Variation in a left ventricle-specific Hand1 enhancer impairs GATA transcription factor binding and disrupts conduction system development and function. Circ. Res. 2019, 125, 575–589. [Google Scholar] [CrossRef]
- Sotoodehnia, N.; Isaacs, A.; De Bakker, P.I.; Dörr, M.; Newton-Cheh, C.; Nolte, I.M.; Van Der Harst, P.; Müller, M.; Eijgelsheim, M.; Alonso, A.; et al. Common variants in 22 loci are associated with QRS duration and cardiac ventricular conduction. Nat. Genet. 2010, 42, 1068–1076. [Google Scholar] [CrossRef] [Green Version]
- Vincentz, J.W.; Toolan, K.P.; Zhang, W.; Firulli, A.B. Hand factor ablation causes defective left ventricular chamber development and compromised adult cardiac function. PLoS Genet. 2017, 13, e1006922. [Google Scholar] [CrossRef]
- Bhattacharyya, S.; Bhakta, M.; Munshi, N.V. Phenotypically silent Cre recombination within the postnatal ventricular conduction system. PLoS ONE 2017, 12, e0174517. [Google Scholar] [CrossRef] [Green Version]
- Morikawa, Y.; Cserjesi, P. Cardiac neural crest expression of Hand2 regulates outflow and second heart field development. Circ. Res. 2008, 103, 1422–1429. [Google Scholar] [CrossRef]
- George, R.M.; Firulli, A.B. Deletion of a Hand1 lncRNA-Containing Septum Transversum Enhancer Alters lncRNA Expression but Is Not Required for Hand1 Expression. J. Cardiovasc. Dev. Dis. 2021, 8, 50. [Google Scholar] [CrossRef]
- Boukens, B.J.; Rivaud, M.R.; Rentschler, S.; Coronel, R. Misinterpretation of the mouse ECG: ‘musing the waves of Mus musculus’. J. Physiol. 2014, 592, 4613–4626. [Google Scholar] [CrossRef]
- Mitchell, G.F.; Jeron, A.; Koren, G. Measurement of heart rate and Q-T interval in the conscious mouse. Am. J. Physiol. 1998, 274, H747–H751. [Google Scholar] [CrossRef]
- Maruyama, M.; Li, B.Y.; Chen, H.; Xu, X.; Song, L.S.; Guatimosim, S.; Zhu, W.; Yong, W.; Zhang, W.; Bu, G.; et al. FKBP12 is a critical regulator of the heart rhythm and the cardiac voltage-gated sodium current in mice. Circ. Res. 2011, 108, 1042–1052. [Google Scholar] [CrossRef]
- Baker, L.C.; London, B.; Choi, B.R.; Koren, G.; Salama, G. Enhanced dispersion of repolarization and refractoriness in transgenic mouse hearts promotes reentrant ventricular tachycardia. Circ. Res. 2000, 86, 396–407. [Google Scholar] [CrossRef] [Green Version]
- VanDusen, N.J.; Casanovas, J.; Vincentz, J.W.; Firulli, B.A.; Osterwalder, M.; Lopez-Rios, J.; Zeller, R.; Zhou, B.; Grego-Bessa, J.; De La Pompa, J.L.; et al. Hand2 is an essential regulator for two Notch-dependent functions within the embryonic endocardium. Cell Rep. 2014, 9, 2071–2083. [Google Scholar] [CrossRef] [Green Version]
- Aanhaanen, W.T.; Boukens, B.J.; Sizarov, A.; Wakker, V.; de Gier-de Vries, C.; van Ginneken, A.C.; Moorman, A.F.; Coronel, R.; Christoffels, V.M. Defective Tbx2-dependent patterning of the atrioventricular canal myocardium causes accessory pathway formation in mice. J. Clin. Investig. 2011, 121, 534–544. [Google Scholar] [CrossRef] [Green Version]
- Munshi, N.V.; McAnally, J.; Bezprozvannaya, S.; Berry, J.M.; Richardson, J.A.; Hill, J.A.; Olson, E.N. Cx30.2 enhancer analysis identifies Gata4 as a novel regulator of atrioventricular delay. Development 2009, 136, 2665–2674. [Google Scholar] [CrossRef] [Green Version]
- McFadden, D.G.; Charité, J.; Richardson, J.A.; Srivastava, D.; Firulli, A.B.; Olson, E.N. A GATA-dependent right ventricular enhancer controls dHAND transcription in the developing heart. Development 2000, 127, 5331–5341. [Google Scholar]
- Cserjesi, P.; Brown, D.; Lyons, G.E.; Olson, E.N. Expression of the novel basic helix-loop-helix gene eHAND in neural crest derivatives and extraembryonic membranes during mouse development. Dev. Biol. 1995, 170, 664–678. [Google Scholar] [CrossRef] [Green Version]
- Howard, M.; Foster, D.N.; Cserjesi, P. Expression of HAND gene products may be sufficient for the differentiation of avian neural crest-derived cells into catecholaminergic neurons in culture. Dev. Biol. 1999, 215, 62–77. [Google Scholar] [CrossRef] [Green Version]
- Hendershot, T.J.; Liu, H.; Clouthier, D.E.; Shepherd, I.T.; Coppola, E.; Studer, M.; Firulli, A.B.; Pittman, D.L.; Howard, M.J. Conditional deletion of Hand2 reveals critical functions in neurogenesis and cell type-specific gene expression for development of neural crest-derived noradrenergic sympathetic ganglion neurons. Dev. Biol. 2008, 319, 179–191. [Google Scholar] [CrossRef] [Green Version]
- Vincentz, J.W.; VanDusen, N.J.; Fleming, A.B.; Rubart, M.; Firulli, B.A.; Howard, M.J.; Firulli, A.B. A Phox2- and Hand2-dependent Hand1 cis-regulatory element reveals a unique gene dosage requirement for Hand2 during sympathetic neurogenesis. J. Neurosci. 2012, 32, 2110–2120. [Google Scholar] [CrossRef]
- Gordan, R.; Gwathmey, J.K.; Xie, L.H. Autonomic and endocrine control of cardiovascular function. World J. Cardiol. 2015, 7, 204–214. [Google Scholar] [CrossRef]
- Harris, J.P.; Bhakta, M.; Bezprozvannaya, S.; Wang, L.; Lubczyk, C.; Olson, E.N.; Munshi, N.V. MyoR modulates cardiac conduction by repressing Gata4. Mol. Cell Biol. 2015, 35, 649–661. [Google Scholar] [CrossRef] [Green Version]
- Dai, Y.S.; Cserjesi, P.; Markham, B.E.; Molkentin, J.D. The transcription factors GATA4 and dHAND physically interact to synergistically activate cardiac gene expression through a p300-dependent mechanism. J. Biol. Chem. 2002, 277, 24390–24398. [Google Scholar] [CrossRef] [Green Version]
- Kreuzberg, M.M.; Schrickel, J.W.; Ghanem, A.; Kim, J.S.; Degen, J.; Janssen-Bienhold, U.; Lewalter, T.; Tiemann, K.; Willecke, K. Connexin30.2 containing gap junction channels decelerate impulse propagation through the atrioventricular node. Proc. Natl. Acad. Sci. USA 2006, 103, 5959–5964. [Google Scholar] [CrossRef] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
George, R.M.; Guo, S.; Firulli, B.A.; Rubart, M.; Firulli, A.B. Neonatal Deletion of Hand1 and Hand2 within Murine Cardiac Conduction System Reveals a Novel Role for HAND2 in Rhythm Homeostasis. J. Cardiovasc. Dev. Dis. 2022, 9, 214. https://doi.org/10.3390/jcdd9070214
George RM, Guo S, Firulli BA, Rubart M, Firulli AB. Neonatal Deletion of Hand1 and Hand2 within Murine Cardiac Conduction System Reveals a Novel Role for HAND2 in Rhythm Homeostasis. Journal of Cardiovascular Development and Disease. 2022; 9(7):214. https://doi.org/10.3390/jcdd9070214
Chicago/Turabian StyleGeorge, Rajani M., Shuai Guo, Beth A. Firulli, Michael Rubart, and Anthony B. Firulli. 2022. "Neonatal Deletion of Hand1 and Hand2 within Murine Cardiac Conduction System Reveals a Novel Role for HAND2 in Rhythm Homeostasis" Journal of Cardiovascular Development and Disease 9, no. 7: 214. https://doi.org/10.3390/jcdd9070214
APA StyleGeorge, R. M., Guo, S., Firulli, B. A., Rubart, M., & Firulli, A. B. (2022). Neonatal Deletion of Hand1 and Hand2 within Murine Cardiac Conduction System Reveals a Novel Role for HAND2 in Rhythm Homeostasis. Journal of Cardiovascular Development and Disease, 9(7), 214. https://doi.org/10.3390/jcdd9070214