
A Contribution to the Phylogeny and Taxonomy of Hydnum (Cantharellales, Basidiomycota) from China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Morphological Studies
2.2. DNA Extraction, PCR Amplification and Sequencing
2.3. Phylogenetic Analyses
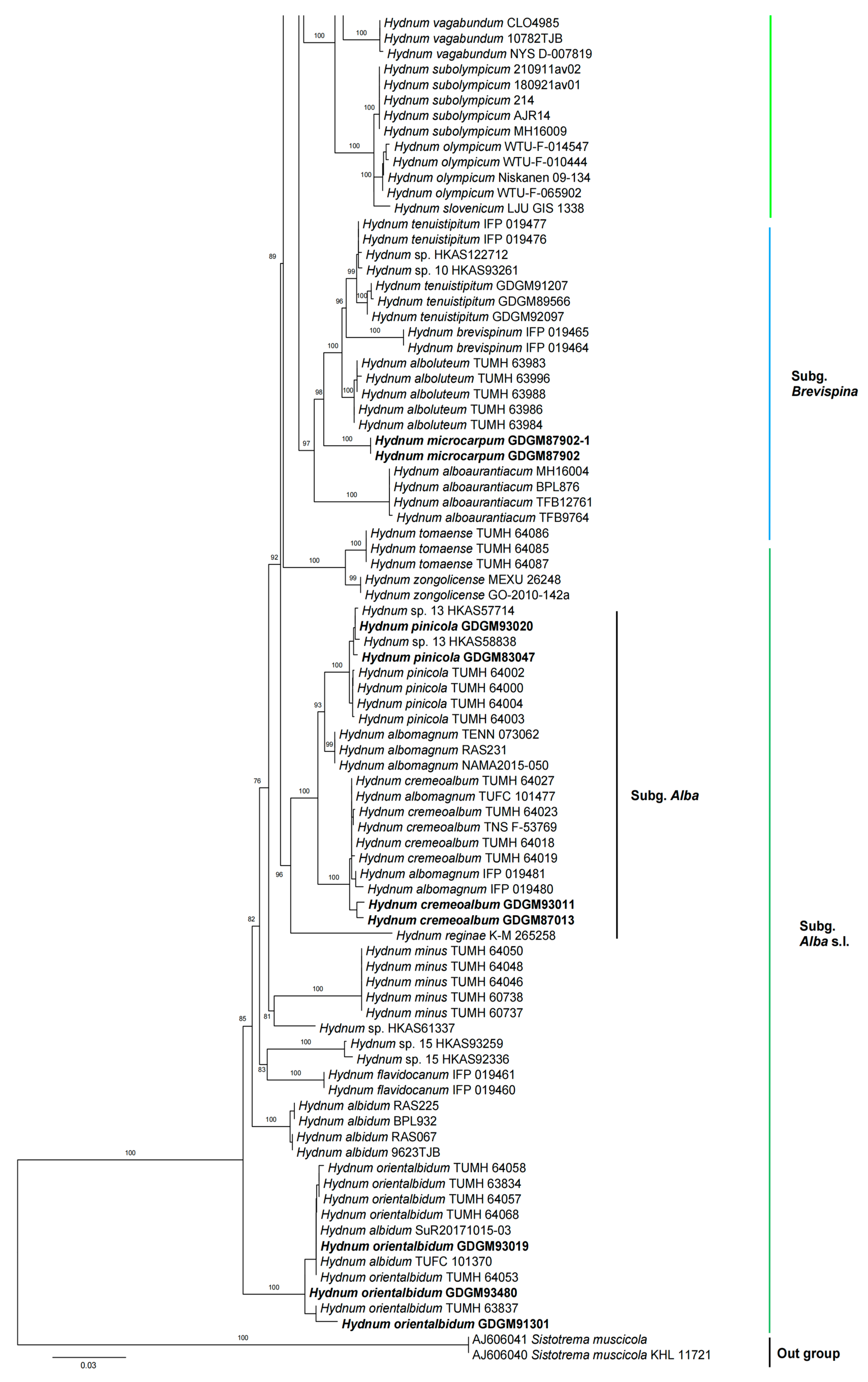
3. Results
3.1. Molecular Phylogeny
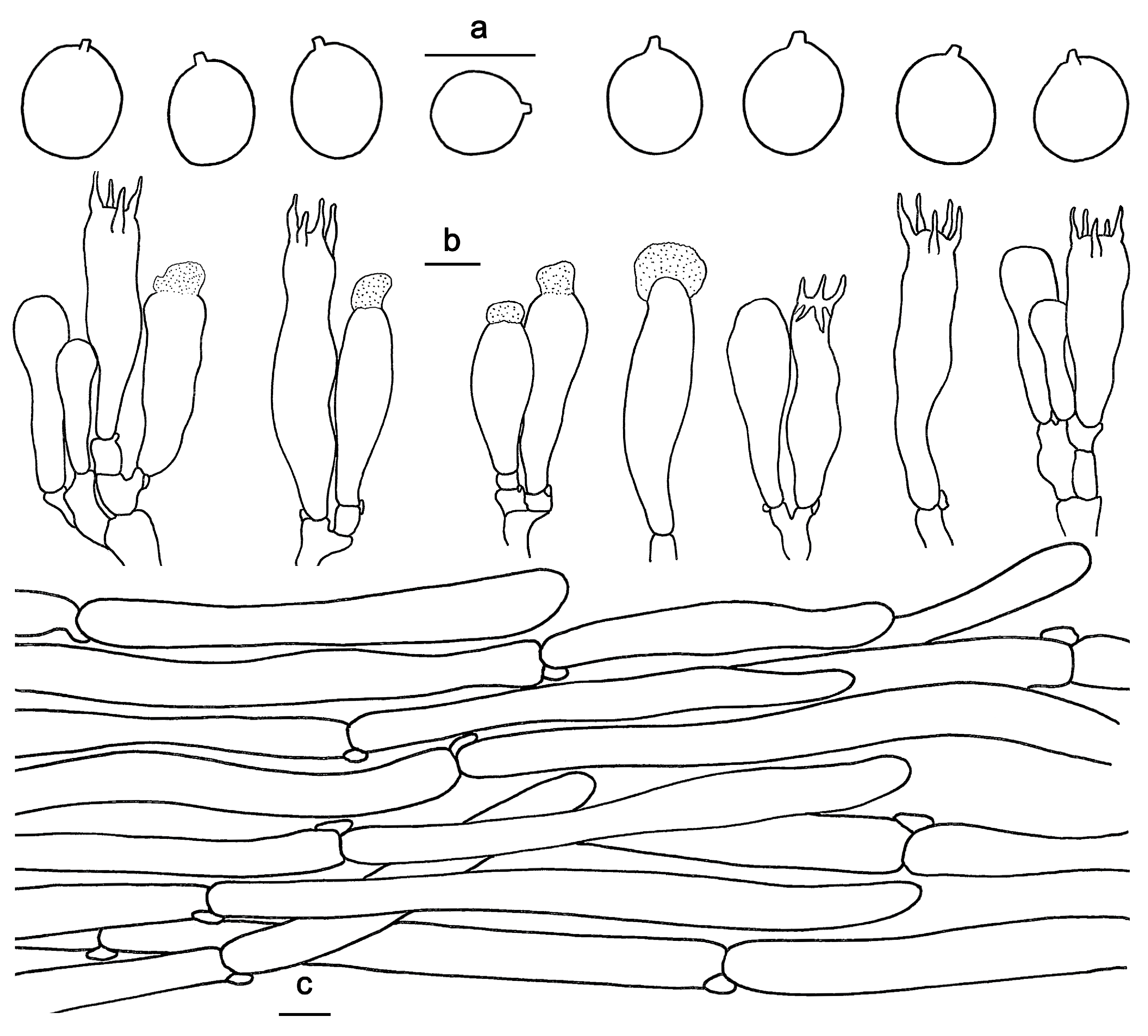
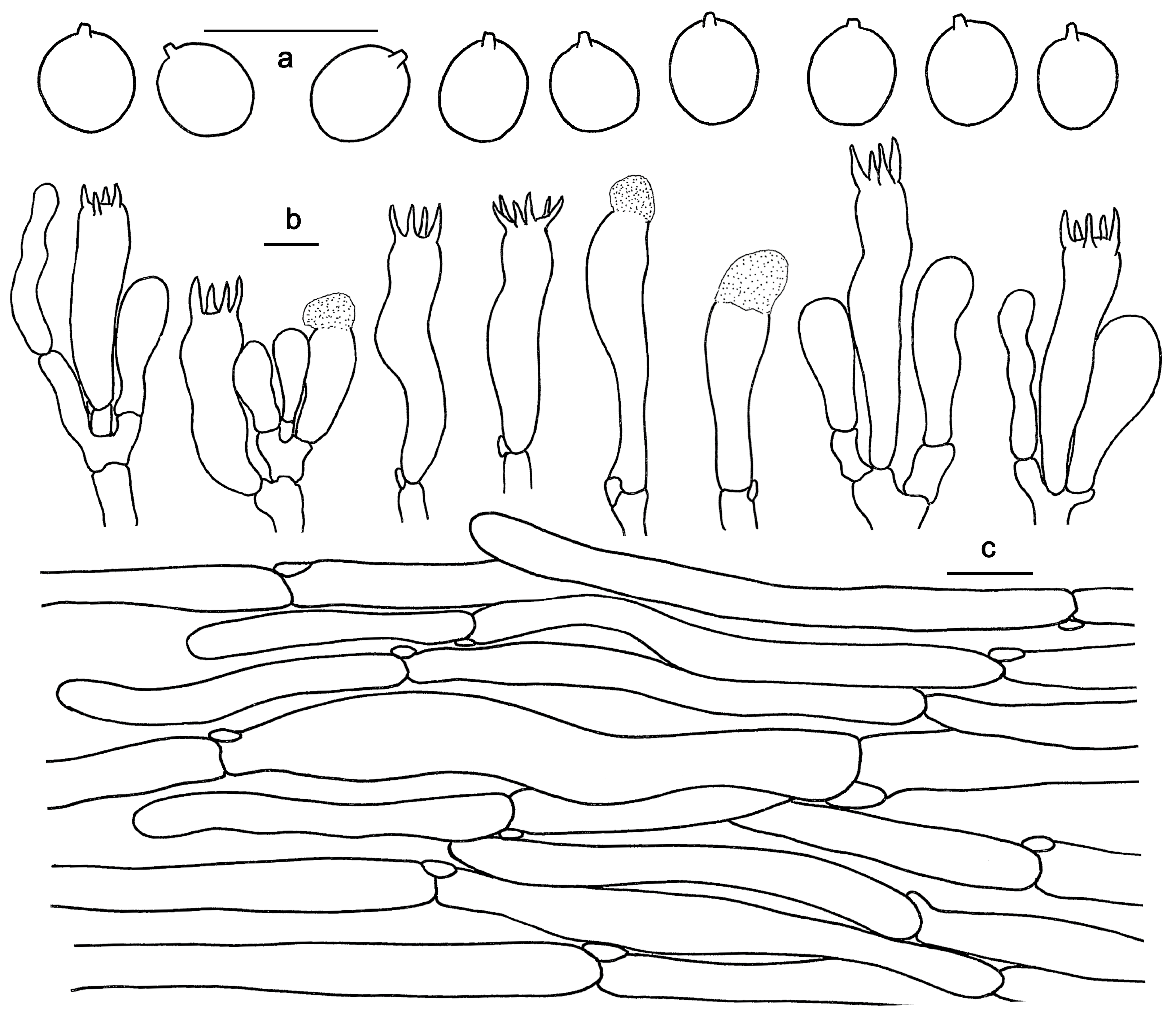
3.2. Taxonomy
4. Discussion
| 1. | Pileus white, cream white, or pale yellow | 2 |
| 1. | Pileus orange, reddish orange, or reddish brown | 11 |
| 2. | Basidiocarps medium to large; pileus > 60 mm broad | 3 |
| 2. | Basidiocarps small; pileus ≤ 60 mm broad | 4 |
| 3. | Distributed in sub-alpine regions | H. sinorepandum |
| 3. | Distributed in subtropical regions, elevation < 1000 m | H. cremeoalbum |
| 4. | Pileus ≤ 25 mm broad | 5 |
| 4. | Pileus usually > 25 mm broad | 7 |
| 5. | Basidiospores > 5 μm long on average | 6 |
| 5. | Basidiospores < 5 μm long on average | H. minum |
| 6. | Spines 0.5–1.5 mm long, white to yellowish white when fresh, grayish yellow to grayish orange when dry | H. microcarpum |
| 6. | Spines 0.2–0.8 mm long, pure white when fresh, yellowish white when dry | H. brevispinum |
| 7. | Basidiospores > 7.5 μm long on average | 8 |
| 7. | Basidiospores < 7.5 μm long on average | 9 |
| 8. | Distributed in northeastern China; basidiospores av. 9.07 × 7.04 μm | H. flabellatum |
| 8. | Distributed in southwestern China; basidiospores av. 7.75 × 6.01 μm | H. flavidocanum |
| 9. | Spines white; basidiospores av. 4.65 × 3.96 μm | H. orientalbidum |
| 9. | Spines orange-white when fresh | 10 |
| 10. | In mixed forests; basidiospores av. 4.72 × 4.02 μm | H. pinicola |
| 10. | In broad-leaved forests; basidiospores av. 7.08 × 6.09 μm | H. tenuistipitum |
| 11. | Basidiocarps small; pileus < 20 mm broad, orange-white to greyish orange; basidiospores av. 9.81 × 9.03 μm | H. longibasidium |
| 11. | Basidiocarps medium to large; pileus ≥ 20 mm broad | 12 |
| 12. | Basidiospores > 9 μm long on average | 13 |
| 12. | Basidiospores < 9 μm long on average | 15 |
| 13. | Holarctic distribution; pileus reddish yellow, light orange to orange | H. melitosarx |
| 13. | Only distributed in Asia | 14 |
| 14. | Distributed in subalpine regions of southwestern China; pileus orange to reddish orange, changing orange-white when bruised; spines adnate, orange-white to pale orange | H. longipes |
| 14. | Distributed in coniferous forests of northwestern China; pileus orange-white to pale orange; spines light yellow when fresh | H. pallidocroceum |
| 15. | Pileus can up to 80 mm broad, pale orange to light orange; spines up to 9 mm long; distributed in temperate regions of Asia | H. berkeleyanum |
| 15. | Pileus less than 80 mm broad | 16 |
| 16. | Spines decurrent | 17 |
| 16. | Spines non-decurrent to subdecurrent | 18 |
| 17. | Pileus very pale to medium orange ocher; basidiospores av. 7.5 × 7 μm; distributed in coniferous forests | H. jussii |
| 17. | Pileus orange-white to pale orange; basidiospores av. 8.75 × 6.99 μm; distributed in broad-leaved forests | H. pallidomarginatum |
| 18. | Distributed in broad-leaved forests in central China | 19 |
| 18. | Distributed in mixed forests | 20 |
| 19. | Spines white, 0.5–3 mm long; pileus orange-white | H. sphaericum |
| 19. | Spines orange-white, 2–6 mm long; pileus light orange to brownish orange | H. tangerinum |
| 20. | Pileus orange to brown; spines orange-white when fresh, up to 5 mm long; basidiospores av. 8.64 × 8.17 μm with Q value = 1.05–1.09 | H. ventricosum |
| 20. | Pileus ocher to light ocher; spines pale ocher, 1–1.7 mm long; basidiospores av. 8.2–8.7 × 6.4–6.8 μm with Q value = 1.27–1.3 | H. vesterholtii |
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hibbett, D.S.; Bauer, R.; Binder, M.; Giachini, A.J.; Hosaka, K.; Justo, A.; Larsson, E.; Larsson, K.H.; Lawrey, J.D.; Miettinen, O.; et al. Agaricomycetes. In The Mycota Systematics and Evolution, 2nd ed.; VII Part A; McLaughlin, D.J., Spatafora, J.W., Eds.; Springer-Verlag: Berlin/Heidelberg, Germany, 2014; pp. 373–429. [Google Scholar] [CrossRef]
- Cao, T.; Hu, Y.P.; Yu, J.R.; Wei, T.Z.; Yuan, H.S. A phylogenetic overview of the Hydnaceae (Cantharellales, Basidiomycota) with new taxa from China. Stud. Mycol. 2021, 99, 100121. [Google Scholar] [CrossRef]
- He, X.L.; Peng, W.H.; Wang, D. An Illustration of Important Wild Edible Fungi in Sichuan; Science Press: Beijing, China, 2021; pp. 1–206. [Google Scholar]
- Yang, Z.L.; Wang, X.H.; Wu, G. Mushrooms of Yunnan; Science Press: Beijing, China, 2022; pp. 1–378. [Google Scholar]
- Feng, B.; Wang, X.H.; Ratkowsky, D.; Gates, G.; Lee, S.S.; Grebenc, T.; Yang, Z.L. Multilocus phylogenetic analyses reveal unexpected abundant diversity and significant disjunct distribution pattern of the hedgehog mushrooms (Hydnum L.). Sci. Rep. 2016, 6, 25586. [Google Scholar] [CrossRef]
- Niskanen, T.; Liimatainen, K.; Nuytinck, J.; Kirk, P.; Olariaga, I.; Roberto Garibay-Orijel, I.; Norvell, L.; Huhtinen, S.; Kytövuori, I.; Ruotsalainen, J.; et al. Identifying and naming the currently known diversity of the genus Hydnum, with an emphasis on European and North American taxa. Mycologia 2018, 110, 890–918. [Google Scholar] [CrossRef]
- Moncalvo, J.M.; Nilsson, R.H.; Koster, B.; Dunham, S.M.; Bernauer, T.; Matheny, P.B.; Porter, T.M.; Margaritescu, S.; Weiß, M.; Garnica, S.; et al. The cantharelloid clade: Dealing with incongruent gene trees and phylogenetic reconstruction methods. Mycologia 2006, 98, 937–948. [Google Scholar] [CrossRef]
- Ying, J.Z.; Zang, M. Economic Macrofungi from Southwestern China; Science Press: Beijing, China, 1994; pp. 1–398. [Google Scholar]
- Zang, M.; Li, B.; Xi, J.; Zhang, D. Fungi of Hengduan Mountains; Science Press: Beijing, China, 1996. [Google Scholar]
- Li, Y.; Li, T.H.; Yang, Z.L.; Bau, T.; Dai, Y.C. Atlas of Chinese Macrofungal Resources; Central China Farmer’s Publishing House: Zhengzhou, China, 2015. [Google Scholar]
- Zhang, M.; Wang, C.Q.; Buyck, B.; Deng, W.Q.; Li, T.H. Multigene Phylogeny and Morphology Reveal Unexpectedly High Number of New Species of Cantharellus Subgenus Parvocantharellus (Hydnaceae, Cantharellales) in China. J. Fungi 2021, 7, 919. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, C.Q.; Gan, M.S.; Li, Y.; Shao, S.C.; Qin, W.Q.; Deng, W.Q.; Li, T.H. Diversity of Cantharellus (Cantharellales, Basidiomycota) in China with Description of Some New Species and New Records. J. Fungi 2022, 8, 483. [Google Scholar] [CrossRef] [PubMed]
- Kornerup, A.; Wanscher, J.H. Methuen Handbook of Colour; Eyre Methuen: London, UK, 1978. [Google Scholar]
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef]
- Buyck, B.; Kauff, F.; Eyssartier, G.; Couloux, A.; Hofstetter, V. A multilocus phylogeny for worldwide Cantharellus (Cantharellales, Agaricomycetidae). Fungal Divers. 2014, 64, 101–121. [Google Scholar] [CrossRef]
- Katoh, K.; Kuma, K.I.; Toh, H.; Miyata, T. MAFFT version 5: Improvement in accuracy of multiple sequence alignment. Nucleic Acids Res. 2005, 33, 511–518. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef]
- Zhang, D.; Gao, F.L.; Li, W.X.; Jakovlić, I.; Zou, H.; Zhang, J.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol. Biol. Evol. 2017, 34, 772–773. [Google Scholar] [CrossRef]
- Kim, J.S.; Lee, W.; Kim, C.; Park, H.; Kim, C.S.; Lim, Y.W. Unveiling the Diversity of Hydnum in the Republic of Korea with One New Species, Hydnum paucispinum. Mycobiology 2023, 51, 300–312. [Google Scholar] [CrossRef]
- Sugawara, R.; Maekawa, N.; Sotome, K.; Nakagiri, A.; Endo, N. Systematic revision of Hydnum species in Japan. Mycologia 2022, 114, 413–452. [Google Scholar] [CrossRef]
- Swenie, R.A.; Baroni, T.J.; Matheny, P.B. Six new species and reports of Hydnum (Cantharellales) from eastern North America. MycoKeys 2018, 72, 35–72. [Google Scholar] [CrossRef]
- Yanaga, K.; Sotome, K.; Ushijima, S.; Maekawa, N. Hydnum species producing whitish basidiomata in Japan. Mycoscience 2015, 56, 434–442. [Google Scholar] [CrossRef]
- Sugawara, R.; Aoki, W.; Yamada, A.; Nakagiri, A.; Endo, N. Ecological speciation of Japanese hedgehog mushroom: Hydnum subalpinum sp. nov. is distinguished from its sister species H. repando-orientale by means of integrative taxonomy. Mycol Progress 2022, 21, 94. [Google Scholar] [CrossRef]
- Wang, X.H.; Das, K.; Horman, J.; Antonin, V.; Baghela, A.; Chakraborty, D.; Hembrom, M.E.; Nakasone, K.; Ortiz-Santana, B.; Vizzini, A.; et al. Fungal Biodiversity Profiles 51–60. Cryptogam. Mycol. 2018, 39, 211–257. [Google Scholar] [CrossRef]
- Olariaga, I.; Grebenc, T.; Salcedo, I.; Martín, M.P. Two new species of Hydnum with ovoid basidiospores: H. ovoideisporum and H. vesterholtii. Mycologia 2012, 104, 1443–1455. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.L. The Macrofungi in China; Henan Science and Technology Press: Zhengzhou, China, 2000; pp. 1–719. [Google Scholar]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Pileus | Spines | Spores Size | Habit and Distribution | References |
|---|---|---|---|---|---|
| H. berkeleyanum | 25–80 mm wide; pale orange to light orange with a brown center, pale orange to light orange toward margin. | Spines adnexed to subdecurrent; initially white (A1), becoming orange-white to pale orange at maturity; 2–9 mm long. | 7.5–8.4–8.5–9.5 × 6.9–7.9–8–8.8 µm; Q = 1.01–1.17 | In temperate mixed forest; Known from India and China | [4,28] |
| H. brevispinum | 10–15 mm wide; pure white to yellowish white, yellowish white to greyish orange when dry. | Spines non-decurrent to subdecurrent; pure white when fresh, yellowish white when dry; 0.2–0.8 mm long. | (4.5)5–5.8(6) × (3.5)3.8–4.8(5) μm, av. 5.28 × 4.16 μm, Q = 1.27–1.31 | In angiosperm forests; distributed in Hunan Province. | [2] |
| H. cremeoalbum | 30–100 mm wide; white to cream white. | Spines adnate, non-decurrent; yellowish white, orange-white to pale orange; 2–5 mm long. | 4–5.5 × 3.5–4.5 μm, av. 4.73 × 3.88 μm, Q = 1.1–1.42. | In broad-leaved or mixed forests; known from southern China and Japan. | [27] Present study |
| H. flabellatum | 30–45 mm wide; yellowish white, pale yellow to greyish yellow, often with brownish orange scales. | Spines non-decurrent or subdecurrent; orange-white when fresh, greyish orange when dry; 0.6–2 mm long. | (7.8)8.5–9.5(10) × (6)6.5–7.8(8) μm, av. 9.07 × 7.04 μm, Q = 1.26–1.29 | In mixed forests; distributed in Liaoning Province. | [2] |
| H. flavidocanum | 20–30 mm wide; yellowish white or yellowish gray at center, whitish towards margin. | Spines non-decurrent or subdecurrent; orange-white when fresh, greyish orange when dry; 0.5–2 mm long, | (7)7.2–8.8(8.9) × (5.2) 5.5–6.5(6.8) μm, av. 7.75 × 6.01 μm, Q = 1.29–1.31 | In mixed forests; distributed in Yunnan Province. | [2] |
| H. jussii | 35–60 mm wide; very pale to medium orange ocher. | Spines somewhat decurrent; whitish to very pale ochraceous, later pale brownish ochraceous. | 7.2–8 × 6.6–7.5 μm, av. 7.5 × 7 μm, Q = 1.03–1.18 | In coniferous forests. Known from Finland and China. | [3,6] |
| H. longibasidium | 10–15 mm wide; orange-white to greyish orange when fresh, greyish orange to brownish yellow when dry. | Spines non-decurrent or subdecurrent; orange-white to pale orange when fresh, greyish orange to brownish yellow when dry; 1–4 mm long. | (8)8.5–11(11.5) × (7.5)7.8–9.8(10) μm, av. 9.81 × 9.03 μm, Q = 1.09–1.13 | In angiosperm forests; distributed in Hunan Province. | [2] |
| H. longipes | 20–30 mm wide; orange, light orange, pastel red to reddish orange, changing to orange-white when bruised. | Spines adnate, non-decurrent; orange-white to pale orange; 1–3 mm long. | (7.5)8–10(10.5) × (6)7–8.5(9) μm, av. 9.32 × 7.63 μm, Q = 1.05–1.43 | In Quercus aquifolioides and Pinus densata dominated forests; distributed in Yunnan Province | Present study |
| H. melitosarx | 20–40 mm wide, reddish yellow, light yellow, light orange to orange. | Spines adnate; white, yellowish white to pale orange; 2–5 mm long. | 8–11× 7–10 μm, av. 9.37 × 8.71 μm, Q = 1–1.28 | In broad-leaved or mixed forests. Known from Asia, Europe, and North America. | [6] Present study |
| H. microcarpum | 10–20 mm wide; white, yellowish white to orange-white, changing from brownish orange to brownish red when bruised. | Spines adnate to subdecurrent; white to yellowish white when fresh, grayish yellow to grayish orange when dry; 0.5–1.5 mm long. | 5–6(6.5) × 5–5.5(6) μm, av. 5.78 × 5.26 μm, Q = 1–1.2 | In mixed forests dominated; distributed in Guangdong Province | Present study |
| H. minum | 10–25 mm wide; whitish, cream yellow to pale buff, changing reddish brown when bruised. | Spines adnate, non-decurrent; whitish, cream yellow to pale buff; 0.5–1.7 mm long. | 4.5–5.5 × 3–4.5 μm, av. 4.8 × 3.8 μm, Q = 1.1–1.5 | In mixed forest; Known from Japan and China. | [2,26] |
| H. orientalbidum | 30–60 mm wide; white. | Spines adnate; white; 2–3 mm long. | 4–5 × 3.5–4.5 μm, av. 4.65 × 3.96 μm, Q = 1.1–1.28 | In broad-leaved forests. Known from China and Japan. | Present study |
| H. pallidocroceum | 25–40 mm wide; orange-white to pale orange. | Spines non-decurrent; light yellow when fresh, orange-white to pale orange when dry; 1–5 mm long, | (7.5) 7.8–9.5(10) × (5.5)6–7.5(8) μm, av. 9.09 × 6.72 μm, Q = 1.32–1.35 | In Pinus sp. and Picea sp. forests; distributed in Xinjiang Province. | [2] |
| H. pallidomarginatum | 20–35 mm wide; orange-white to pale orange, with a light color zone towards center. | Spines decurrent; orange-white to pale orange when fresh, brownish orange when dry; 0.5–2 mm long, | (8) 8.2–9.8(10) × (6)6.5–7.8(8.2) μm, av. 8.75 × 6.99 μm, Q = 1.25–1.28 | In angiosperm forests; distributed in Yunnan Province. | [2] |
| H. pinicola | 25–60 mm wide; yellowish white, pale yellow to pale orange. | Spines adnate; orange-white, pale orange when fresh, pinkish white to pale red when dry; 1.5–3 mm long. | 4–6 × 3.5–5 μm, av. 4.72 × 4.02 μm, Q = 1–1.25 | In mixed forests; known from China and Japan. | [24] Present study |
| H. sinorepandum | 40–120 mm wide; yellowish white, pale yellow, light yellow to light orange. | Spines adnate; white, yellowish white, pale yellow to pale orange; 3–7 mm long. | (7.5)8–9(10) × 6.5–7.5 μm, av. 8.16 × 7.12 μm, Q = 1–1.38 | In broad-leaved and mixed forests; distributed in Yunnan Province. | Present study |
| H. sphaericum | 20–35 mm wide; orange-white when moist, greyish orange to brownish orange when dry. | Spines non-decurrent to subdecurrent; white when fresh, brownish orange when dry; 0.5–3 mm long, | (7.5)8–8.8(9) × (6)6.5–7.5(8) μm, av. 8.36 × 6.94 μm, Q = 1.2–1.23 | In angiosperm forest; distributed in Hunan Province. | [2] |
| H. tangerinum | 10–50 mm wide; pale orange, light orange, orange to brownish orange when moist, greyish orange when dry. | Spines non-decurrent; orange-white when fresh, greyish orange when dry; 2–6 mm long. | (7)7.2–8.8(9) × (5.5) 5.8–7(7.5) μm, av. 8.11 × 6.19 μm, Q = 1.23–1.31 | In angiosperm forest; distributed in Hunan Province | [2] |
| H. tenuistipitum | 10–30 mm wide; yellow white to orange-white when moist, pale orange to greyish orange when dry. | Spines non-decurrent to subdecurrent; orange-white when fresh, light brown when dry; 1–3 mm long. | (6.5)6.8–7.2(7.5) × (5.2)5.5–6.5(6.8) μm, av. 7.08 × 6.09 μm, Q = 1.07–1.16 | In angiosperm forest; distributed in Hunan Province. | [2] |
| H. ventricosum | 28–35 mm wide; orange to brown. | Spines non-decurrent; orange-white when fresh, brownish orange when dry; 1–5 mm long. | (7.5)8.2–9(9.5) × (7)7.5–8.5(9) μm, av. 8.64 × 8.17 μm, Q = 1.05–1.09 | In mixed forests; distributed in Liaoning Province. | [2] |
| H. vesterholtii | 10–30(–50) mm wide, ocher to light ocher. | Spines non-decurrent to slightly decurrent; pale ocher; 1–1.7 mm long. | (7)8–9(9.5) × 6–7.5(8) μm, av. 8.2–8.7 × 6.4–6.8 μm; Q = 1.27–1.3; | In Abies alba and Fagus sylvatica forest. Known from China and France. | [3,29] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.; Wang, C.; Bai, H.; Deng, W. A Contribution to the Phylogeny and Taxonomy of Hydnum (Cantharellales, Basidiomycota) from China. J. Fungi 2024, 10, 98. https://doi.org/10.3390/jof10020098
Zhang M, Wang C, Bai H, Deng W. A Contribution to the Phylogeny and Taxonomy of Hydnum (Cantharellales, Basidiomycota) from China. Journal of Fungi. 2024; 10(2):98. https://doi.org/10.3390/jof10020098
Chicago/Turabian StyleZhang, Ming, Chaoqun Wang, Hongfen Bai, and Wangqiu Deng. 2024. "A Contribution to the Phylogeny and Taxonomy of Hydnum (Cantharellales, Basidiomycota) from China" Journal of Fungi 10, no. 2: 98. https://doi.org/10.3390/jof10020098
APA StyleZhang, M., Wang, C., Bai, H., & Deng, W. (2024). A Contribution to the Phylogeny and Taxonomy of Hydnum (Cantharellales, Basidiomycota) from China. Journal of Fungi, 10(2), 98. https://doi.org/10.3390/jof10020098