
The Host Response to Coccidioidomycosis

 , , and
, , and
Abstract
:1. Introduction
2. Innate Immunity
2.1. Local Factors in the Lung
2.2. Polymorphonuclear Leukocytes
2.3. Monocytes/Macrophages and Dendritic Cells
2.4. Cellular Receptors and Pathways for the Innate Immune Response
2.4.1. C-Type Lectin Receptors
2.4.2. Toll-Like Receptors (TLR)
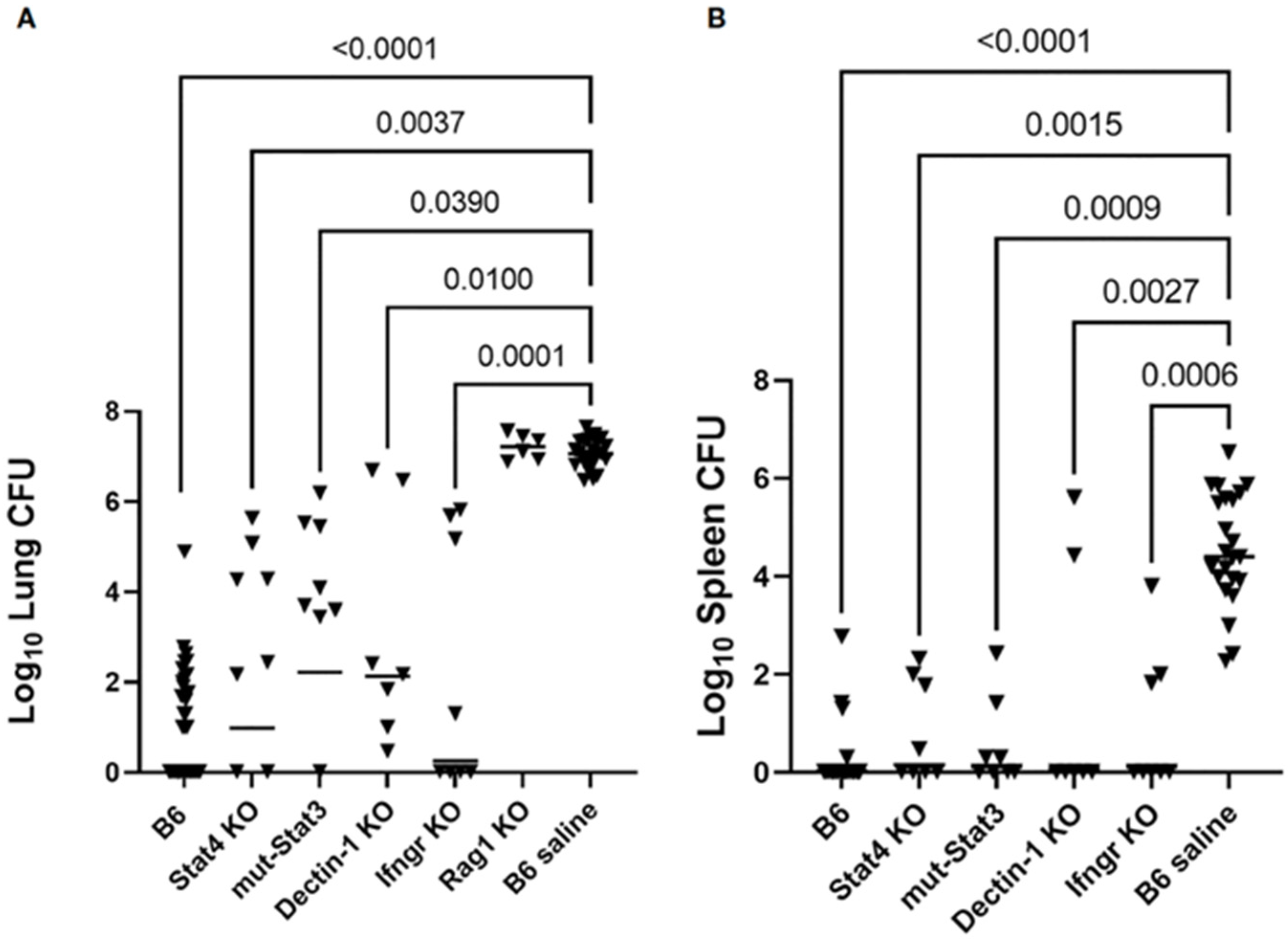
2.4.3. In Vivo Studies of Murine Innate Immunity
2.5. Genetically Determined Resistance to Infection and Innate Immunity in Mice
2.6. Genetic Predisposition to Severe Disease in Human Beings
2.7. Effector Mechanisms
3. Acquired Immunity
3.1. B-lymphocytes
3.2. T-Lymphocytes
3.2.1. Studies in Mice
3.2.2. Immunity in Humans
4. Vaccines
4.1. Whole Spherule and Spherule Extract Vaccines
4.2. Live Attenuated Mutants
4.3. Vaccine Candidate Proteins

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antigen | Form | Adjuvant | Activity | Reference |
|---|---|---|---|---|
| Live attenuated mutants | N/A | N/A | 4 + | [79] [104] |
| Formalin-killed spherules | N/A | N/A | 4 + mice - humans | [96] [101] |
| Spherule extract 1 | N/A | Various | 4 + | [96] |
| Ag2/PRA | Protein, DNA | Various | 2 + | [110] |
| Ag2/PRA | Protein, DNA | Various | - protein 1 + cDNA | [113,114] |
| Expression library 1 | DNA | None | 3 + | [121] |
| Prp2 | Protein | CpG | 1 + | [122] |
| B-glucanosltransferase | Protein | CpG | 2 + | [123] |
| Calnexin | Protein | Glucan and Adjuplex | 2 + | [115] |
| Aspartyl protease | Protein | CpG | 3 + | [116] |
| CSA 2 | Protein | CpG and MPLA 3 | 1 + | [112] |
| Multivalent antigens | ||||
| Ag2/PRA CSA fusion protein | Protein | CpG and MPLA | 3 + | [112] |
| Phospholipase, α-mannosidase and aspartyl protease peptide fusion protein | Protein | CpG glucan particles | 2 + | [117] |
| rCpa1 4 | Protein | Glucan chitin particles | 4 + | [119] |
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Smith, C.E. Epidemiology of Acute Coccidioidomycosis with Erythema Nodosum (“San Joaquin” or “Valley Fever”). Am. J. Public Health Nations Health 1940, 30, 600–611. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, C.; Barker, B.M.; Hoover, S.; Nix, D.E.; Ampel, N.M.; Frelinger, J.A.; Orbach, M.J.; Galgiani, J.N. Recent advances in our understanding of the environmental, epidemiological, immunological, and clinical dimensions of coccidioidomycosis. Clin. Microbiol. Rev. 2013, 26, 505–525. [Google Scholar] [CrossRef] [PubMed]
- Kirkland, T.N.; Fierer, J. Coccidioides immitis and posadasii; A review of their biology, genomics, pathogenesis, and host immunity. Virulence 2018, 9, 1426–1435. [Google Scholar] [CrossRef] [PubMed]
- Kirkland, T.N.; Stevens, D.A.; Hung, C.Y.; Beyhan, S.; Taylor, J.W.; Shubitz, L.F.; Duttke, S.H.; Heidari, A.; Johnson, R.H.; Deresinski, S.C.; et al. Coccidioides Species: A Review of Basic Research: 2022. J. Fungi 2022, 8, 859. [Google Scholar] [CrossRef] [PubMed]
- Kollath, D.R.; Miller, K.J.; Barker, B.M. The mysterious desert dwellers: Coccidioides immitis and Coccidioides posadasii, causative fungal agents of coccidioidomycosis. Virulence 2019, 10, 222–233. [Google Scholar] [CrossRef] [PubMed]
- Hung, C.Y.; Hsu, A.P.; Holland, S.M.; Fierer, J. A review of innate and adaptive immunity to coccidioidomycosis. Med. Mycol. 2019, 57, S85–S92. [Google Scholar] [CrossRef]
- Cox, R.A.; Magee, D.M. Coccidioidomycosis: Host response and vaccine development. Clin. Microbiol. Rev. 2004, 17, 804–839. [Google Scholar] [CrossRef]
- Ward, R.A.; Vyas, J.M. The first line of defense: Effector pathways of anti-fungal innate immunity. Curr. Opin. Microbiol. 2020, 58, 160–165. [Google Scholar] [CrossRef]
- Kirkland, T.N.; Fierer, J. Innate Immune Receptors and Defense Against Primary Pathogenic Fungi. Vaccines 2020, 8, 303. [Google Scholar] [CrossRef]
- Donovan, F.M.; Shubitz, L.; Powell, D.; Orbach, M.; Frelinger, J.; Galgiani, J.N. Early Events in Coccidioidomycosis. Clin. Microbiol. Rev. 2019, 33, e00112-19. [Google Scholar] [CrossRef] [PubMed]
- Medzhitov, R. Approaching the asymptote: 20 years later. Immunity 2009, 30, 766–775. [Google Scholar] [CrossRef] [PubMed]
- Ward, R.A.; Thompson, G.R., 3rd; Villani, A.C.; Li, B.; Mansour, M.K.; Wuethrich, M.; Tam, J.M.; Klein, B.S.; Vyas, J.M. The Known Unknowns of the Immune Response to Coccidioides. J. Fungi 2021, 7, 377. [Google Scholar] [CrossRef] [PubMed]
- Hsu, A.P.; Korzeniowska, A.; Aguilar, C.C.; Gu, J.; Karlins, E.; Oler, A.J.; Chen, G.; Reynoso, G.V.; Davis, J.; Chaput, A.; et al. Immunogenetics associated with severe coccidioidomycosis. JCI Insight 2022, 7, e159491. [Google Scholar] [CrossRef] [PubMed]
- Margolis, D.A.; Viriyakosol, S.; Fierer, J.; Kirkland, T.N. The role of reactive oxygen intermediates in experimental coccidioidomycois in mice. BMC Microbiol. 2011, 11, 71. [Google Scholar] [CrossRef] [PubMed]
- Awasthi, S.; Magee, D.M.; Coalson, J.J. Coccidioides posadasii infection alters the expression of surfactant proteins SP-A and SP-D. Respir. Res. 2004, 5, 28. [Google Scholar] [CrossRef] [PubMed]
- Galgiani, J.N.; Isenberg, R.A.; Stevens, D.A. Chemotaxigenic activity of extracts from the mycelial and spherule phases of Coccidioides immitis for human polymorphonuclear leukocytes. Infect. Immun. 1978, 21, 862–865. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.Y.; Thompson, G.R., 3rd; Hastey, C.J.; Hodge, G.C.; Lunetta, J.M.; Pappagianis, D.; Heinrich, V. Coccidioides Endospores and Spherules Draw Strong Chemotactic, Adhesive, and Phagocytic Responses by Individual Human Neutrophils. PLoS ONE 2015, 10, e0129522. [Google Scholar] [CrossRef]
- Galgiani, J.N.; Payne, C.M.; Jones, J.F. Human polymorphonuclear-leukocyte inhibition of incorporation of chitin precursors into mycelia of Coccidioides immitis. J. Infect. Dis. 1984, 149, 404–412. [Google Scholar] [CrossRef]
- Galgiani, J.N. Inhibition of different phases of Coccidioides immitis by human neutrophils or hydrogen peroxide. J. Infect. Dis. 1986, 153, 217–222. [Google Scholar] [CrossRef]
- Frey, C.L.; Drutz, D.J. Influence of fungal surface components on the interaction of Coccidioides immitis with polymorphonuclear neutrophils. J. Infect. Dis. 1986, 153, 933–943. [Google Scholar] [CrossRef]
- Segal, G.P.; Lehrer, R.I.; Selsted, M.E. In Vitro Effect of Phagocyte Cationic Peptides on Coccidioides immitis. J. Infect. Dis. 1985, 151, 890–894. [Google Scholar] [CrossRef] [PubMed]
- Carlin, A.F.; Viriyakosol, S.; Okamoto, S.; Walls, L.; Fierer, J. Interleukin-8 Receptor 2 (IL-8R2)-Deficient Mice Are More Resistant to Pulmonary Coccidioidomycosis than Control Mice. Infect. Immun. 2020, 89, e00883-19. [Google Scholar] [CrossRef]
- Hung, C.Y.; Jiménez-Alzate, M.; Gonzalez, A.; Wüthrich, M.; Klein, B.S.; Cole, G.T. Interleukin-1 receptor but not Toll-like receptor 2 is essential for MyD88-dependent Th17 immunity to Coccidioides infection. Infect. Immun. 2014, 82, 2106–2114. [Google Scholar] [CrossRef] [PubMed]
- Roy, R.M.; Klein, B.S. Dendritic cells in antifungal immunity and vaccine design. Cell Host Microbe 2012, 11, 436–446. [Google Scholar] [CrossRef] [PubMed]
- Marakalala, M.J.; Martinez, F.O.; Plüddemann, A.; Gordon, S. Macrophage Heterogeneity in the Immunopathogenesis of Tuberculosis. Front. Microbiol. 2018, 9, 1028. [Google Scholar] [CrossRef]
- Dalod, M.; Scheu, S. Dendritic cell functions in vivo: A user’s guide to current and next- generation mutant mouse models. Eur. J. Immunol. 2022, 52, 1712–1749. [Google Scholar] [CrossRef]
- Beaman, L.; Benjamini, E.; Pappagianis, D. Role of lymphocytes in macrophage-induced killing of Coccidioides immitis in vitro. Infect. Immun. 1981, 34, 347–353. [Google Scholar] [CrossRef]
- Beaman, L.; Benjamini, E.; Pappagianis, D. Activation of macrophages by lymphokines: Enhancement of phagosome-lysosome fusion and killing of Coccidioides immitis. Infect. Immun. 1983, 39, 1201–1207. [Google Scholar] [CrossRef]
- Diep, A.L.; Tejeda-Garibay, S.; Miranda, N.; Hoyer, K.K. Macrophage and Dendritic Cell Activation and Polarization in Response to Coccidioidesposadasii Infection. J. Fungi 2021, 7, 630. [Google Scholar] [CrossRef]
- Taylor, P.R.; Brown, G.D.; Reid, D.M.; Willment, J.A.; Martinez-Pomares, L.; Gordon, S.; Wong, S.Y. The beta-glucan receptor, dectin-1, is predominantly expressed on the surface of cells of the monocyte/macrophage and neutrophil lineages. J. Immunol. 2002, 169, 3876–3882. [Google Scholar] [CrossRef]
- Viriyakosol, S.; Jimenez Mdel, P.; Gurney, M.A.; Ashbaugh, M.E.; Fierer, J. Dectin-1 is required for resistance to coccidioidomycosis in mice. mBio 2013, 4, e00597-12. [Google Scholar] [CrossRef]
- Del Pilar Jiménez-A, M.; Viriyakosol, S.; Walls, L.; Datta, S.K.; Kirkland, T.; Heinsbroek, S.E.M.; Brown, G.; Fierer, J. Susceptibility to Coccidioides species in C57BL/6 mice is associated with expression of a truncated splice variant of Dectin-1 (Clec7a). Genes Immun. 2008, 9, 338–348. [Google Scholar] [CrossRef] [PubMed]
- Campuzano, A.; Zhang, H.; Ostroff, G.R.; Dos Santos Dias, L.; Wuthrich, M.; Klein, B.S.; Yu, J.J.; Lara, H.H.; Lopez-Ribot, J.L.; Hung, C.Y. CARD9-Associated Dectin-1 and Dectin-2 Are Required for Protective Immunity of a Multivalent Vaccine against Coccidioides posadasii Infection. J. Immunol. 2020, 204, 3296–3306. [Google Scholar] [CrossRef] [PubMed]
- Kirkland, T.N.; Fierer, J. Inbred Mouse Strains Differ in Resistance to Lethal Coccidioides immitis Infection. Infect. Immun. 1983, 40, 912–916. [Google Scholar] [CrossRef] [PubMed]
- Heinsbroek, S.E.; Taylor, P.R.; Rosas, M.; Willment, J.A.; Williams, D.L.; Gordon, S.; Brown, G.D. Expression of functionally different dectin-1 isoforms by murine macrophages. J. Immunol. 2006, 176, 5513–5518. [Google Scholar] [CrossRef] [PubMed]
- Powell, D.A.; Hsu, A.P.; Butkiewicz, C.D.; Trinh, H.T.; Frelinger, J.A.; Holland, S.M.; Galgiani, J.N.; Shubitz, L.F. Vaccine Protection of Mice With Primary Immunodeficiencies Against Disseminated Coccidioidomycosis. Front. Cell Infect. Microbiol. 2021, 11, 790488. [Google Scholar] [CrossRef] [PubMed]
- Viriyakosol, S.; Jimenez Mdel, P.; Saijo, S.; Fierer, J. Neither dectin-2 nor the mannose receptor is required for resistance to Coccidioides immitis in mice. Infect. Immun. 2014, 82, 1147–1156. [Google Scholar] [CrossRef] [PubMed]
- Dionne, S.O.; Podany, A.B.; Ruiz, Y.W.; Ampel, N.M.; Galgiani, J.N.; Lake, D.F. Spherules Derived from Coccidioides posadasii Promote Human Dendritic Cell Maturation and Activation. Infect. Immun. 2006, 74, 2415–2422. [Google Scholar] [CrossRef] [PubMed]
- Ampel, N.M.; Nelson, D.K.; Li, L.; Dionne, S.O.; Lake, D.F.; Simmons, K.A.; Pappagianis, D. The Mannose Receptor Mediates the Cellular Immune Response in Human Coccidioidomycosis. Infect. Immun. 2005, 73, 2554–2555. [Google Scholar] [CrossRef]
- Wang, H.; LeBert, V.; Hung, C.Y.; Galles, K.; Saijo, S.; Lin, X.; Cole, G.T.; Klein, B.S.; Wuthrich, M. C-type lectin receptors differentially induce th17 cells and vaccine immunity to the endemic mycosis of North America. J. Immunol. 2014, 192, 1107–1119. [Google Scholar] [CrossRef]
- Viriyakosol, S.; Fierer, J.; Brown, G.D.; Kirkland, T.N. Innate immunity to the pathogenic fungus Coccidioides posadasii is dependent on TLR2 and dectin-1. Infect. Immun. 2005, 73, 1553–1560. [Google Scholar] [CrossRef] [PubMed]
- Viriyakosol, S.; Walls, L.; Okamoto, S.; Raz, E.; Williams, D.L.; Fierer, J. Myeloid Differentiation Factor 88 and Interleukin-1R1 Signaling Contribute to Resistance to Coccidioides immitis. Infect. Immun. 2018, 86, e00028-18. [Google Scholar] [CrossRef] [PubMed]
- Lewis, E.R.; David, V.R.; Doyle, A.L.; Rajabi, K.; Kiefer, J.A.; Pirrotte, P.; Barker, B.M. Differences in Host Innate Responses among Coccidioides Isolates in a Murine Model of Pulmonary Coccidioidomycosis. Eukaryot. Cell 2015, 14, 1043–1053. [Google Scholar] [CrossRef] [PubMed]
- Hung, C.Y.; Castro-Lopez, N.; Cole, G.T. Card9- and MyD88-Mediated Gamma Interferon and Nitric Oxide Production Is Essential for Resistance to Subcutaneous Coccidioides posadasii Infection. Infect. Immun. 2016, 84, 1166–1175. [Google Scholar] [CrossRef] [PubMed]
- Higgins Keppler, E.A.; Van Dyke, M.C.C.; Mead, H.L.; Lake, D.F.; Magee, D.M.; Barker, B.M.; Bean, H.D. Volatile Metabolites in Lavage Fluid Are Correlated with Cytokine Production in a Valley Fever Murine Model. J. Fungi 2023, 9, 115. [Google Scholar] [CrossRef] [PubMed]
- Kirkland, T.N.; Fierer, J. Genetic control of resistance to Coccidioides immitis: A single gene that is expressed in spleen cells determines resistance. J. Immunol. 1985, 135, 548–552. [Google Scholar] [CrossRef] [PubMed]
- Cox, R.A.; Kennell, W.; Boncyk, L.; Murphy, J.W. Induction and expression of cell-mediated immune responses in inbred mice infected with Coccidioides immitis. Infect. Immun. 1988, 56, 13–17. [Google Scholar] [CrossRef]
- Magee, D.M.; Cox, R.A. Interleukin-12 regulation of host defenses against Coccidioides immitis. Infect. Immun. 1996, 64, 3609–3613. [Google Scholar] [CrossRef]
- Fierer, J.; Walls, L.; Eckmann, L.; Yamamoto, T.; Kirkland, T.N. Importance of interleukin-10 in genetic susceptibility of mice to Coccidioides immitis. Infect. Immun. 1998, 66, 4397–4402. [Google Scholar] [CrossRef]
- Fierer, J.; Walls, L.; Wright, F.; Kirkland, T.N. Genes influencing resistance to Coccidioides immitis and the interleukin-10 response map to chromosomes 4 and 6 in mice. Infect. Immun. 1999, 67, 2916–2919. [Google Scholar] [CrossRef]
- Willment, J.A.; Gordon, S.; Brown, G.D. Characterization of the human beta-glucan receptor and its alternatively spliced isoforms. J. Biol. Chem. 2001, 276, 43818–43823. [Google Scholar] [CrossRef]
- Woelk, C.H.; Zhang, J.X.; Walls, L.; Viriyakosol, S.; Singhania, A.; Kirkland, T.N.; Fierer, J. Factors regulated by interferon gamma and hypoxia-inducible factor 1A contribute to responses that protect mice from Coccidioides immitis infection. BMC Microbiol. 2012, 12, 218. [Google Scholar] [CrossRef] [PubMed]
- Vinh, D.C.; Masannat, F.; Dzioba, R.B.; Galgiani, J.N.; Holland, S.M. Refractory disseminated coccidioidomycosis and mycobacteriosis in interferon-gamma receptor 1 deficiency. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2009, 49, e62–e65. [Google Scholar] [CrossRef] [PubMed]
- Vinh, D.C.; Schwartz, B.; Hsu, A.P.; Miranda, D.J.; Valdez, P.A.; Fink, D.; Lau, K.P.; Long-Priel, D.; Kuhns, D.B.; Uzel, G.; et al. Interleukin-12 Receptor 1 Deficiency Predisposing to Disseminated Coccidioidomycosis. Clin. Infect. Dis. 2011, 52, e99–e102. [Google Scholar] [CrossRef]
- Sampaio, E.P.; Hsu, A.P.; Pechacek, J.; Bax, H.I.; Dias, D.L.; Paulson, M.L.; Chandrasekaran, P.; Rosen, L.B.; Carvalho, D.S.; Ding, L.; et al. Signal transducer and activator of transcription 1 (STAT1) gain-of-function mutations and disseminated coccidioidomycosis and histoplasmosis. J. Allergy Clin. Immunol. 2013, 131, 1624–1634. [Google Scholar] [CrossRef] [PubMed]
- Shubitz, L.F.; Powell, D.A.; Butkiewicz, C.D.; Lewis, M.L.; Trinh, H.T.; Frelinger, J.A.; Orbach, M.J.; Galgiani, J.N. A Chronic Murine Disease Model of Coccidioidomycosis Using Coccidioides posadasii, strain 1038. J. Infect. Dis. 2020, 223, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.; Fleming, P.; Ciesielski, C.; Hu, D.; Kaplan, J.; Ward, J. Coccidioidomycosis among persons with AIDS in the united states. J. Infect. Dis. 1995, 171, 961–964. [Google Scholar] [CrossRef]
- Blair, J.E.; Ampel, N.M.; Hoover, S.E. Coccidioidomycosis in selected immunosuppressed hosts. Med. Mycol. 2019, 57, S56–S63. [Google Scholar] [CrossRef] [PubMed]
- Kirkland, T.N.; Fierer, J. Coccidioidomycosis: A reemerging infectious disease. Emerg. Infect. Dis. 1996, 2, 192. [Google Scholar] [CrossRef]
- Bays, D.J.; Thompson, G.R.; Reef, S.; Snyder, L.; Freifeld, A.J.; Huppert, M.; Salkin, D.; Wilson, M.D.; Galgiani, J.N. Natural History of Disseminated Coccidioidomycosis: Examination of the Veterans Affairs-Armed Forces Database. Clin. Infect. Dis. 2021, 73, e3814–e3819. [Google Scholar] [CrossRef]
- Ruddy, B.E.; Mayer, A.P.; Ko, M.G.; Labonte, H.R.; Borovansky, J.A.; Boroff, E.S.; Blair, J.E. Coccidioidomycosis in African Americans. Mayo Clin. Proc. 2011, 86, 63–69. [Google Scholar] [CrossRef]
- Rosenstein, N.E.; Emery, K.W.; Werner, S.B.; Kao, A.; Johnson, R.; Rogers, D.; Vugia, D.; Reingold, A.; Talbot, R.; Plikaytis, B.D.; et al. Risk factors for severe pulmonary and disseminated coccidioidomycosis: Kern County, California, 1995–1996. Clin. Infect. Dis. 2001, 32, 708–715. [Google Scholar] [CrossRef]
- Wheeler, C.; Lucas, K.D.; Mohle-Boetani, J.C. Rates and risk factors for Coccidioidomycosis among prison inmates, California, USA, 2011. Emerg. Infect. Dis. 2015, 21, 70–75. [Google Scholar] [CrossRef]
- Louie, L.; Ng, S.; Hajjeh, R.; Johnson, R.; Vugia, D.; Werner, S.B.; Talbot, R.; Klitz, W. Influence of host genetics on the severity of coccidioidomycosis. Emerg. Infect. Dis. 1999, 5, 672–680. [Google Scholar] [CrossRef]
- Morand, S.; Ueyama, T.; Tsujibe, S.; Saito, N.; Korzeniowska, A.; Leto, T.L. Duox maturation factors form cell surface complexes with Duox affecting the specificity of reactive oxygen species generation. FASEB J. 2009, 23, 1205–1218. [Google Scholar] [CrossRef]
- Henriet, S.; Verweij, P.E.; Holland, S.M.; Warris, A. Invasive fungal infections in patients with chronic granulomatous disease. Adv. Exp. Med. Biol. 2013, 764, 27–55. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, A.; Hung, C.Y.; Cole, G.T. Absence of phagocyte NADPH oxidase 2 leads to severe inflammatory response in lungs of mice infected with Coccidioides. Microb. Pathog. 2011, 51, 432–441. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, A.; Hung, C.Y.; Cole, G.T. Nitric oxide synthase activity has limited influence on the control of Coccidioides infection in mice. Microb. Pathog. 2011, 51, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, M.d.P.; Walls, L.; Fierer, J. High Levels of Interleukin-10 Impair Resistance to Pulmonary Coccidioidomycosis in Mice in Part through Control of Nitric Oxide Synthase 2 Expression. Infect. Immun. 2006, 74, 3387–3395. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.E.; Saito, M.T.; Simons, S.A. Pattern of 39,500 serologic tests in coccidioidomycosis. J. Am. Med. Assoc. 1956, 160, 546–552. [Google Scholar] [CrossRef]
- Clemons, K.V.; Leathers, C.R.; Lee, K.W. Systemic Coccidioides immitis infection in nude and beige mice. Infect. Immun. 1985, 47, 814–821. [Google Scholar] [CrossRef]
- Beaman, L.; Pappagianis, D.; Benjamini, E. Mechanisms of resistance to infection with Coccidioides immitis in mice. Infect. Immun. 1979, 23, 681–685. [Google Scholar] [CrossRef]
- Kong, Y.M.; Savage, D.C.; Levine, H.B. Enhancement of immune responses in mice by a booster injection of Coccidioides spherules. J. Immunol. 1965, 95, 1048–1056. [Google Scholar] [CrossRef]
- Magee, D.M.; Friedberg, R.L.; Woitaske, M.D.; Johnston, S.A.; Cox, R.A. Role of B cells in vaccine-induced immunity against coccidioidomycosis. Infect. Immun. 2005, 73, 7011–7013. [Google Scholar] [CrossRef] [PubMed]
- Fierer, J.; Waters, C.; Walls, L. Both CD4+ and CD8+ T cells can medicate vaccine-induced protection against Coccidioides immitis infection in mice. J. Infect. Dis. 2006, 193, 1323–1331. [Google Scholar] [CrossRef]
- Mosmann, T.R.; Cherwinski, H.; Bond, M.W.; Giedlin, M.A.; Coffman, R.L. Two types of murine helper T cell clone. I. Definition according to profiles of lymphokine activities and secreted proteins. J. Immunol. 1986, 136, 2348–2357. [Google Scholar] [CrossRef] [PubMed]
- Kara, E.E.; Comerford, I.; Fenix, K.A.; Bastow, C.R.; Gregor, C.E.; McKenzie, D.R.; McColl, S.R. Tailored immune responses: Novel effector helper T cell subsets in protective immunity. PLoS Pathog. 2014, 10, e1003905. [Google Scholar] [CrossRef]
- Kirkland, T.N.; Raz, E.; Datta, S.K. Molecular and cellular mechanisms of protective immunity to coccidioidomycosis. Vaccine 2006, 24, 495–500. [Google Scholar] [CrossRef]
- Xue, J.; Chen, X.; Selby, D.; Hung, C.Y.; Yu, J.J.; Cole, G.T. A genetically engineered live attenuated vaccine of Coccidioides posadasii protects BALB/c mice against coccidioidomycosis. Infect. Immun. 2009, 77, 3196–3208. [Google Scholar] [CrossRef] [PubMed]
- Wüthrich, M.; Gern, B.; Hung, C.Y.; Ersland, K.; Rocco, N.; Pick-Jacobs, J.; Galles, K.; Filutowicz, H.; Warner, T.; Evans, M.; et al. Vaccine-induced protection against 3 systemic mycoses endemic to North America requires Th17 cells in mice. J. Clin. Investig. 2011, 121, 554–568. [Google Scholar] [CrossRef]
- Hung, C.Y.; Gonzalez, A.; Wüthrich, M.; Klein, B.S.; Cole, G.T. Vaccine immunity to coccidioidomycosis occurs by early activation of three signal pathways of T helper cell response (Th1, Th2, and Th17). Infect. Immun. 2011, 79, 4511–4522. [Google Scholar] [CrossRef]
- Smith, C.E.; Whiting, E.G.; Baker, E.E.; Rosenberger, H.G.; Beard, R.R.; Saito, M.T. The use of coccidioidin. Am. Rev. Tuberc. 1948, 57, 330–360. [Google Scholar]
- Oldfield, E.D.; Bone, W.D.; Martin, C.R.; Gray, G.C.; Olson, P.; Schillaci, R.F. Prediction of relapse after treatment of coccidioidomycosis. Clin. Infect. Dis. 1997, 25, 1205–1210. [Google Scholar] [CrossRef]
- Mafi, N.; Murphy, C.B.; Girardo, M.E.; Blair, J.E. Coccidioides (spherulin) skin testing in patients with pulmonary coccidioidomycosis in an endemic regiondagger. Med. Mycol. 2020, 58, 626–631. [Google Scholar] [CrossRef]
- Zweiman, B.; Pappagianis, D.; Maibach, H.; Hildreth, E.A. Coccidioidin delayed hypersensitivity: Skin test and in vitro lymphocyte reactivities. J. Immunol. 1969, 102, 1284–1289. [Google Scholar] [CrossRef]
- Cox, R.A.; Vivas, J.R.; Gross, A.; Lecara, G.; Miller, E.; Brummer, E. In vivo and in vitro cell-mediated responses in coccidioidomycosis. I. Immumologic responses of persons with primary, asymptomatic infections. Am. Rev. Respir. Dis. 1976, 114, 937–943. [Google Scholar]
- Cox, R.A.; Brummer, E.; Lecara, G. In vitro lymphocyte responses of coccidioidin skin test-positive and -negative persons to coccidioidin, spherulin, and a coccidioides cell wall antigen. Infect. Immun. 1977, 15, 751–755. [Google Scholar] [CrossRef]
- Ampel, N.M.; Bejarano, G.C.; Salas, S.D.; Galgiani, J.N. In vitro assessment of cellular immunity in human coccidioidomycosis: Relationship between dermal hypersensitivity, lymphocyte transformation, and lymphokine production by peripheral blood mononuclear cells from healthy adults. J. Infect. Dis. 1992, 165, 710–715. [Google Scholar] [CrossRef] [PubMed]
- Ampel, N.M.; Nelson, D.K.; Chavez, S.; Naus, K.A.; Herman, A.B.; Li, L.; Simmons, K.A.; Pappagianis, D. Preliminary evaluation of whole-blood gamma interferon release for clinical assessment of cellular immunity in patients with active coccidioidomycosis. Clin. Diagn. Lab. Immunol. 2005, 12, 700–704. [Google Scholar] [CrossRef] [PubMed]
- Corry, D.B.; Ampel, N.M.; Christian, L.; Locksley, R.M.; Galgiani, J.N. Cytokine production by peripheral blood mononuclear cells in human coccidioidomycosis. J. Infect. Dis. 1996, 174, 440–443. [Google Scholar] [CrossRef] [PubMed]
- Ampel, N.M.; Kramer, L.A.; Kerekes, K.M.; Johnson, S.M.; Pappagianis, D. Assessment of the human cellular immune response to T27K, a coccidioidal antigen preparation, by flow cytometry of whole blood. Med. Mycol. 2001, 39, 315–520. [Google Scholar] [CrossRef]
- Ampel, N.M.; Robey, I.; Nguyen, C.T.; Roller, B.; August, J.; Knox, K.S.; Pappagianis, D. Ex Vivo Cytokine Release, Determined by a Multiplex Cytokine Assay, in Response to Coccidioidal Antigen Stimulation of Whole Blood among Subjects with Recently Diagnosed Primary Pulmonary Coccidioidomycosis. mSphere 2018, 3, e00065-18. [Google Scholar] [CrossRef] [PubMed]
- Davini, D.; Naeem, F.; Phong, A.; Al-Kuhlani, M.; Valentine, K.M.; McCarty, J.; Ojcius, D.M.; Gravano, D.M.; Hoyer, K.K. Elevated regulatory T cells at diagnosis of Coccidioides infection associates with chronicity in pediatric patients. J. Allergy Clin. Immunol. 2018, 142, 1971–1974.e7. [Google Scholar] [CrossRef]
- Li, L.; Dial, S.M.; Schmelz, M.; Rennels, M.A.; Ampel, N.M. Cellular immune suppressor activity resides in lymphocyte cell clusters adjacent to granulomata in human coccidioidomycosis. Infect. Immun. 2005, 73, 3923–3928. [Google Scholar] [CrossRef]
- Tsai, M.; Thauland, T.J.; Huang, A.Y.; Bun, C.; Fitzwater, S.; Krogstad, P.; Douine, E.D.; Nelson, S.F.; Lee, H.; Garcia-Lloret, M.I.; et al. Disseminated Coccidioidomycosis Treated with Interferon-gamma and Dupilumab. N. Engl. J. Med. 2020, 382, 2337–2343. [Google Scholar] [CrossRef] [PubMed]
- Pappagianis, D.; Hector, R.; Levine, H.B.; Collins, M.S. Immunization of mice against coccidioidomycosis with a subcellular vaccine. Infect. Immun. 1979, 25, 440–445. [Google Scholar] [CrossRef]
- Zimmermann, C.R.; Johnson, S.M.; Martens, G.W.; White, A.G.; Zimmer, B.L.; Pappagianis, D. Protection against lethal murine coccidioidomycosis by a soluble vaccine from spherules. Infect. Immun. 1998, 66, 2342–2345. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.M.; Kerekes, K.M.; Lunetta, J.M.; Pappagianis, D. Characteristics of the protective subcellular coccidioidal T27K vaccine. Ann. N. Y. Acad. Sci. 2007, 1111, 275–289. [Google Scholar] [CrossRef]
- Levine, H.B.; Cobb, J.M.; Smith, C.E. Immunogenicity of spherule-endospore vaccines of Coccidioides immitis for mice. J. Immunol. 1961, 87, 218–227. [Google Scholar] [CrossRef]
- Williams, P.L.; Sable, D.L.; Sorgen, S.P.; Pappagianis, D.; Levine, H.B.; Brodine, S.K.; Brown, B.W.; Grumet, F.C.; Stevens, D.A. Immunologic responsiveness and safety associated with the Coccidioides immitis spherule vaccine in volunteers of white, black, and Filipino ancestry. Am. J. Epidemiol. 1984, 119, 591–602. [Google Scholar] [CrossRef]
- Pappagianis, D. Evaluation of the protective efficacy of the killed Coccidioides immitis spherule vaccine in humans. The Valley Fever Vaccine Study Group. Am. Rev. Respir. Dis. 1993, 148, 656–660. [Google Scholar] [CrossRef]
- Walch, H.A.; Kalvoda, A. Immunization of mice with induced mutants of Coccidioides immitis. I. Characterization of mutants and preliminary studies of their use as viable vaccines. Sabouraudia 1971, 9, 173–184. [Google Scholar] [CrossRef]
- Hurtgen, B.J.; Castro-Lopez, N.; Jiménez-Alzate, M.D.P.; Cole, G.T.; Hung, C.Y. Preclinical identification of vaccine induced protective correlates in human leukocyte antigen expressing transgenic mice infected with Coccidioides posadasii. Vaccine 2016, 34, 5336–5343. [Google Scholar] [CrossRef]
- Narra, H.P.; Shubitz, L.F.; Mandel, M.A.; Trinh, H.T.; Griffin, K.; Buntzman, A.S.; Frelinger, J.A.; Galgiani, J.N.; Orbach, M.J. A Coccidioides posadasii CPS1 Deletion Mutant Is Avirulent and Protects Mice from Lethal Infection. Infect. Immun. 2016, 84, 3007–3016. [Google Scholar] [CrossRef]
- Shubitz, L.F.; Powell, D.A.; Trinh, H.T.; Lewis, M.L.; Orbach, M.J.; Frelinger, J.A.; Galgiani, J.N. Viable spores of Coccidioides posadasii Deltacps1 are required for vaccination and provide long lasting immunity. Vaccine 2018, 36, 3375–3380. [Google Scholar] [CrossRef] [PubMed]
- Shubitz, L.F.; Robb, E.J.; Powell, D.A.; Bowen, R.A.; Bosco-Lauth, A.; Hartwig, A.; Porter, S.M.; Trinh, H.; Moale, H.; Bielefeldt-Ohmann, H.; et al. Δcps1 vaccine protects dogs against experimentally induced coccidioidomycosis. Vaccine 2021, 39, 6894–6901. [Google Scholar] [CrossRef] [PubMed]
- Mandel, M.A.; Beyhan, S.; Voorhies, M.; Shubitz, L.F.; Galgiani, J.N.; Orbach, M.J.; Sil, A. The WOPR family protein Ryp1 is a key regulator of gene expression, development, and virulence in the thermally dimorphic fungal pathogen Coccidioides posadasii. PLoS Pathog. 2022, 18, e1009832. [Google Scholar] [CrossRef]
- Beyhan, S.; Gutierrez, M.; Voorhies, M.; Sil, A. A Temperature-Responsive Network Links Cell Shape and Virulence Traits in a Primary Fungal Pathogen. PLoS Biol. 2013, 11, e1001614. [Google Scholar] [CrossRef]
- Hung, C.; Ampel, N.M.; Christian, L.; Seshan, K.R.; Cole, G.T. A major cell surface antigen of Coccidioides immitis which elicits both humoral and cellular immune responses. Infect. Immun. 2000, 68, 584–593. [Google Scholar] [CrossRef]
- Abuodeh, R.O.; Shubitz, L.F.; Siegel, E.; Snyder, S.; Peng, T.; Orsborn, K.I.; Brummer, E.; Stevens, D.A.; Galgiani, J.N. Resistance to Coccidioides immitis in mice after immunization with recombinant protein or a DNA vaccine of a proline-rich antigen. Infect. Immun. 1999, 67, 2935–2940. [Google Scholar] [CrossRef] [PubMed]
- Kirkland, T.N.; Finley, F.; Orsborn, K.I.; Galgiani, J.N. Evaluation of the proline-rich antigen of Coccidioides immitis as a vaccine candidate in mice. Infect. Immun. 1998, 66, 3519–3522. [Google Scholar] [CrossRef]
- Shubitz, L.F.; Yu, J.J.; Hung, C.Y.; Kirkland, T.N.; Peng, T.; Perrill, R.; Simons, J.; Xue, J.; Herr, R.A.; Cole, G.T.; et al. Improved protection of mice against lethal respiratory infection with Coccidioides posadasii using two recombinant antigens expressed as a single protein. Vaccine 2006, 24, 5904–5911. [Google Scholar] [CrossRef]
- Jiang, C.; Magee, D.M.; Quitugua, T.N.; Cox, R.A. Genetic vaccination against Coccidioides immitis: Comparison of vaccine efficacy of recombinant antigen 2 and antigen 2 cDNA. Infect. Immun. 1999, 67, 630–635. [Google Scholar] [CrossRef]
- Jiang, C.; Magee, D.M.; Cox, R.A. Coadministration of interleukin 12 expression vector with antigen 2 cDNA enhances induction of protective immunity against Coccidioides immitis. Infect. Immun. 1999, 67, 5848–5853. [Google Scholar] [CrossRef]
- Wüthrich, M.; Brandhorst, T.T.; Sullivan, T.D.; Filutowicz, H.; Sterkel, A.; Stewart, D.; Li, M.; Lerksuthirat, T.; LeBert, V.; Shen, Z.T.; et al. Calnexin induces expansion of antigen-specific CD4(+) T cells that confer immunity to fungal ascomycetes via conserved epitopes. Cell Host Microbe 2015, 17, 452–465. [Google Scholar] [CrossRef]
- Tarcha, E.J.; Basrur, V.; Hung, C.Y.; Gardner, M.J.; Cole, G.T. A recombinant aspartyl protease of Coccidioides posadasii induces protection against pulmonary coccidioidomycosis in mice. Infect. Immun. 2006, 74, 516–527. [Google Scholar] [CrossRef]
- Hurtgen, B.J.; Hung, C.Y.; Ostroff, G.R.; Levitz, S.M.; Cole, G.T. Construction and evaluation of a novel recombinant T cell epitope-based vaccine against Coccidioidomycosis. Infect. Immun. 2012, 80, 3960–3974. [Google Scholar] [CrossRef]
- Chu, R.S.; Targoni, O.S.; Krieg, A.M.; Lehmann, P.V.; Harding, C.V. CpG oligodeoxynucleotides act as adjuvants that switch on T helper 1 (Th1) immunity. J. Exp. Med. 1997, 186, 1623–1631. [Google Scholar] [CrossRef] [PubMed]
- Hung, C.Y.; Zhang, H.; Castro-Lopez, N.; Ostroff, G.R.; Khoshlenar, P.; Abraham, A.; Cole, G.T.; Negron, A.; Forsthuber, T.; Peng, T.; et al. Glucan-Chitin Particles Enhance Th17 Response and Improve Protective Efficacy of a Multivalent Antigen (rCpa1) against Pulmonary Coccidioides posadasii Infection. Infect. Immun. 2018, 86, e00070-18. [Google Scholar] [CrossRef] [PubMed]
- Kirkland, T.N. The Quest for a Vaccine Against Coccidioidomycosis: A Neglected Disease of the Americas. J. Fungi 2016, 2, 34. [Google Scholar] [CrossRef] [PubMed]
- Ivey, F.D.; Magee, D.M.; Woitaske, M.D.; Johnston, S.A.; Cox, R.A. Identification of a protective antigen of Coccidioides immitis by expression library immunization. Vaccine 2003, 21, 4359–4367. [Google Scholar] [CrossRef]
- Herr, R.A.; Hung, C.Y.; Cole, G.T. Evaluation of two homologous proline-rich proteins of Coccidioides posadasii as candidate vaccines against coccidioidomycosis. Infect. Immun. 2007, 75, 5777–5787. [Google Scholar] [CrossRef] [PubMed]
- Delgado, N.; Xue, J.; Yu, J.-J.; Hung, C.-Y.; Cole, G.T. A recombinant beta-1,3-glucanosyltransferase homolog of Coccidioides posadasii protects mice against coccidioidomycosis. Infect. Immun. 2003, 71, 3010–3019. [Google Scholar] [CrossRef] [PubMed]
- Dowling, J.K.; Mansell, A. Toll-like receptors: The swiss army knife of immunity and vaccine development. Clin. Transl. Immunol. 2016, 5, e85. [Google Scholar] [CrossRef] [PubMed]
- Reed, S.G.; Orr, M.T.; Fox, C.B. Key roles of adjuvants in modern vaccines. Nat. Med. 2013, 19, 1597–1608. [Google Scholar] [CrossRef]
- Krogstad, P.; Thompson, G.R., 3rd; Heidari, A.; Kuran, R.; Stephens, A.V.; Butte, M.J.; Johnson, R. A Clinicopathological Categorization System for Clinical Research in Coccidioidomycosis. Open Forum Infect. Dis. 2023, 10, ofad597. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kirkland, T.N.; Hung, C.-Y.; Shubitz, L.F.; Beyhan, S.; Fierer, J. The Host Response to Coccidioidomycosis. J. Fungi 2024, 10, 173. https://doi.org/10.3390/jof10030173
Kirkland TN, Hung C-Y, Shubitz LF, Beyhan S, Fierer J. The Host Response to Coccidioidomycosis. Journal of Fungi. 2024; 10(3):173. https://doi.org/10.3390/jof10030173
Chicago/Turabian StyleKirkland, Theo N., Chiung-Yu Hung, Lisa F. Shubitz, Sinem Beyhan, and Joshua Fierer. 2024. "The Host Response to Coccidioidomycosis" Journal of Fungi 10, no. 3: 173. https://doi.org/10.3390/jof10030173
APA StyleKirkland, T. N., Hung, C. -Y., Shubitz, L. F., Beyhan, S., & Fierer, J. (2024). The Host Response to Coccidioidomycosis. Journal of Fungi, 10(3), 173. https://doi.org/10.3390/jof10030173