
Molecular Characterization of the First Partitivirus from a Causal Agent of Salvia miltiorrhiza Dry Rot

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
2.2. Extraction and Purification of DsRNA
2.3. Synthesis, Cloning and Sequencing of cDNA
2.4. Sequence and Phylogenetic Analysis
2.5. Acquisition of Virus-Free Mycoviruses
2.6. RNA Extraction and RT-PCR Detection
2.7. Assay of Biological Properties of Mycovirus-Infected and Mycovirus-Free Strains
3. Results
3.1. DsRNA Segments in F. oxysporum Strains
3.2. Molecular Characterization of FoPV1 in Strain FCR51
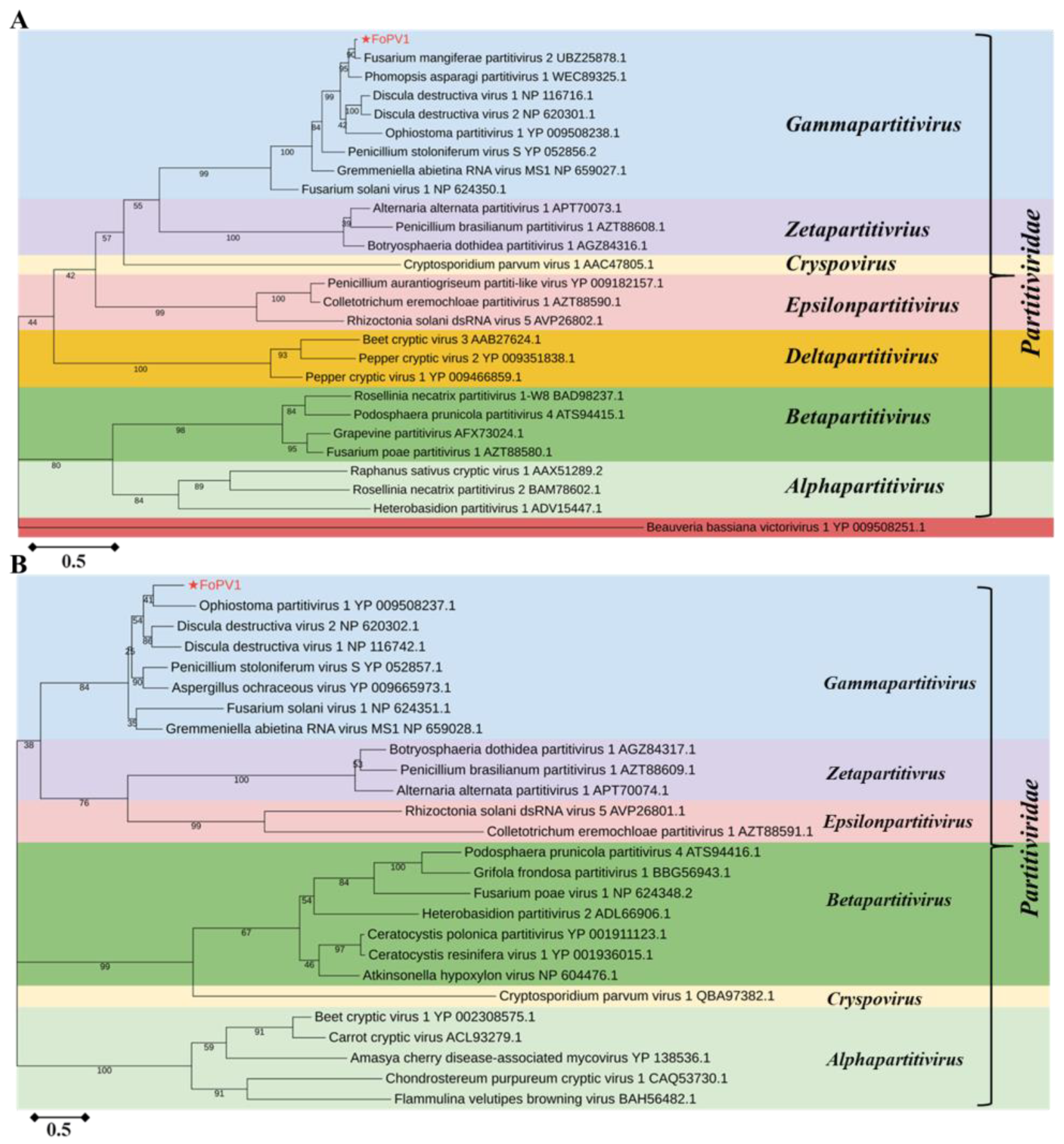
3.3. Phylogenetic Analysis of FoPV1
3.4. Effects of FoPV1 on F. oxysporum Strain FCR51
3.4.1. Biological Effects of FoPV1 on F. oxysporum Strain FCR51
3.4.2. Acquisition of FoPV1 Virus-Free Strain
3.4.3. Effects of FoPV1 on F. oxysporum Strain FCR51
3.5. Influence of FoPV1 to the Virulence of F. oxysporum
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xie, J.; Jiang, D. New insights into mycoviruses and exploration for the biological control of crop fungal diseases. Annu. Rev. Phytopathol. 2014, 52, 45–68. [Google Scholar] [CrossRef] [PubMed]
- Ghabrial, S.A.; Castón, J.R.; Jiang, D.; Nibert, M.L.; Suzuki, N. 50-plus years of fungal viruses. Virology 2015, 479, 356–368. [Google Scholar] [CrossRef] [PubMed]
- Kondo, H.; Botella, L.; Suzuki, N. Mycovirus diversity and evolution revealed/inferred from recent studies. Annu. Rev. Phytopathol. 2022, 60, 307–336. [Google Scholar] [CrossRef] [PubMed]
- Ghabrial, S.A.; Suzuki, N. Viruses of plant pathogenic fungi. Annu. Rev. Phytopathol. 2009, 47, 353–384. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Yang, G.; Shi, N.; Zhao, C.; Hu, F.; Coutts, R.H.; Kotta-Loizou, I.; Huang, B. A novel partitivirus orchestrates conidiation, stress response, pathogenicity, and secondary metabolism of the entomopathogenic fungus Metarhizium majus. PLoS Pathog. 2023, 19, e1011397. [Google Scholar] [CrossRef] [PubMed]
- Nibert, M.L.; Ghabrial, S.A.; Maiss, E.; Lesker, T.; Vainio, E.J.; Jiang, D.; Suzuki, N. Taxonomic reorganization of family Partitiviridae and other recent progress in partitivirus research. Virus Res. 2014, 188, 128–141. [Google Scholar] [CrossRef] [PubMed]
- Vainio, E.J.; Chiba, S.; Ghabrial, S.A.; Maiss, E.; Roossinck, M.; Sabanadzovic, S.; Suzuki, N.; Xie, J.; Nibert, M. ICTV virus taxonomy profile: Partitiviridae. J. Gen. Virol. 2018, 99, 17–18. [Google Scholar] [CrossRef] [PubMed]
- Bhatti, M.F.; Bignell, E.M.; Coutts, R.H. Complete nucleotide sequences of two dsRNAs associated with a new partitivirus infecting Aspergillus fumigatus. Arch. Virol. 2011, 156, 1677–1680. [Google Scholar] [CrossRef]
- Jiang, Y.; Luo, C.; Jiang, D.; Li, G.; Huang, J. The complete genomic sequence of a second novel partitivirus infecting Ustilaginoidea virens. Arch. Virol. 2014, 159, 1865–1868. [Google Scholar] [CrossRef]
- Shiba, K.; Hatta, C.; Sasai, S.; Tojo, M.; T Ohki, S.; Mochizuki, T. Genome sequence of a novel partitivirus identified from the oomycete Pythium nunn. Arch. Virol. 2018, 163, 2561–2563. [Google Scholar] [CrossRef]
- Peyambari, M.; Habibi, M.K.; Fotouhifar, K.B.; Dizadji, A.; Roossinck, M.J. Molecular characterization of a novel putative partitivirus infecting Cytospora sacchari, a plant pathogenic fungus. Plant Pathology J. 2014, 30, 151. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Zeng, M.; Zhang, M.; Shu, C.; Zhou, E. Complete nucleotide sequence of a partitivirus from Rhizoctonia solani AG-1 IA strain C24. Viruses 2018, 10, 703. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Yu, Z.; Sun, Y.; Yang, M.; Jiang, N. Molecular characterization of a novel partitivirus isolated from Rhizoctonia solani. Front. Microbiol. 2022, 13, 978075. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J.; Yang, Z.Z.; Yang, X.; Guo, Z.J.; Xie, W.; Zhang, Y.J. Molecular characterization of a novel partitivirus and a fusarivirus coinfecting the fungus Nigrospora sphaerica. Arch. Virol. 2021, 166, 2325–2331. [Google Scholar] [CrossRef] [PubMed]
- Hassan, S.; Syun-Ichi, U.; Shabeer, S.; Wu, C.F.; Moriyama, H.; Coutts, R.H.; Kotta-Loizou, I.; Jamal, A. Molecular and biological characterization of a partitivirus from Paecilomyces variotii. J. Gen. Virol. 2023, 104, 001925. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.Z.; Guo, J.; Hu, Z.; Zhang, X.T.; Li, X.G.; Zhong, J. A novel partitivirus that confer hypovirulence to the plant pathogenic fungus Colletotrichum liriopes. Front. Microbiol. 2021, 12, 653809. [Google Scholar] [CrossRef] [PubMed]
- Kamaruzzaman, M.; He, G.; Wu, M.; Zhang, J.; Yang, L.; Chen, W.; Li, G. A novel partitivirus in the hypovirulent isolate QT5-19 of the plant pathogenic fungus Botrytis cinerea. Viruses 2019, 11, 24. [Google Scholar] [CrossRef]
- Zhu, J.Z.; Qiu, Z.L.; Gao, B.D.; Li, X.G.; Zhong, J. A novel partitivirus conferring hypovirulence by affecting vesicle transport in the fungus Colletotrichum. Mbio 2024, 15, e02530-23. [Google Scholar] [CrossRef]
- Chun, J.; Yang, H.E.; Kim, D.H. Identification of a novel partitivirus of Trichoderma harzianum NFCF319 and evidence for the related antifungal activity. Front. Plant Sci. 2018, 9, 1699. [Google Scholar] [CrossRef]
- Sasaki, A.; Nakamura, H.; Suzuki, N.; Kanematsu, S. Characterization of a new megabirnavirus that confers hypovirulence with the aid of a co-infecting partitivirus to the host fungus, Rosellinia necatrix. Virus Res. 2016, 219, 73–82. [Google Scholar] [CrossRef]
- Lau, S.K.; Lo, G.C.; Chow, F.W.; Fan, R.Y.; Cai, J.J.; Yuen, K.Y.; Woo, P.C. Novel partitivirus enhances virulence of and causes aberrant gene expression in Talaromyces marneffei. MBio 2018, 9, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Lefkowitz, E.J.; Adams, M.J.; Davison, A.J.; Siddell, S.G.; Simmonds, P. Virus Taxonomy: Classification and Nomenclature of Viruses; The Online 10th Report of the ICTV; Springer Science & Business Media: New York, NY, USA, 2017. [Google Scholar]
- Nuss, D.L. Hypovirulence: Mycoviruses at the fungal–plant interface. Nat. Rev. Microbiol. 2005, 3, 632–642. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.D.; Zhang, L.; Li, G.Q.; Jiang, D.H.; Hou, M.S.; Huang, H.C. Hypovirulence and double-stranded RNA in Botrytis cinerea. Phytopathology 2007, 97, 1590–1599. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Zhang, M.; Chen, Q.; Zhu, M.; Zhou, E. A novel mycovirus closely related to viruses in the genus Alphapartitivirus confers hypovirulence in the phytopathogenic fungus Rhizoctonia solani. Virology 2014, 456, 220–226. [Google Scholar] [CrossRef] [PubMed]
- Kotta-Loizou, I. Mycoviruses and their role in fungal pathogenesis. Curr. Opin. Microbiol. 2021, 63, 10–18. [Google Scholar] [CrossRef]
- Xiao, X.; Cheng, J.; Tang, J.; Fu, Y.; Jiang, D.; Baker, T.S.; Ghabrial, S.A.; Xie, J. A novel partitivirus that confers hypovirulence on plant pathogenic fungi. J. Virol. 2014, 88, 10120–10133. [Google Scholar] [CrossRef]
- Arjona-López, J.M.; Telengech, P.; Suzuki, N.; López-Herrera, C.J. Coinfection of Rosellinia necatrix by a partitivirus and a virga-like virus is associated with hypovirulence. Eur. J. Plant Pathol. 2020, 158, 111–119. [Google Scholar] [CrossRef]
- Gordon, T.R. Fusarium oxysporum and the Fusarium wilt syndrome. Annu. Rev. Phytopathol. 2017, 55, 23–39. [Google Scholar] [CrossRef]
- Hanson, L.; De Lucchi, C.; Stevanato, P.; McGrath, M.; Panella, L.; Sella, L.; Biaggi, M.; Concheri, G. Root rot symptoms in sugar beet lines caused by Fusarium oxysporum f. sp. betae. Eur. J. Plant Pathol. 2018, 150, 589–593. [Google Scholar] [CrossRef]
- Yang, J.; Wang, F.; Wen, Y.; Gao, S.; Lu, C.; Liu, Y.; Liu, H. First report of Fusarium proliferatum causing root rot disease in Salvia miltiorrhizae in China. Plant Dis. 2020, 105, 1210. [Google Scholar]
- Anagnostakis, S.L. Biological control of chestnut blight. Science 1982, 215, 466–471. [Google Scholar] [CrossRef]
- Yu, X.; Li, B.; Fu, Y.; Xie, J.; Cheng, J.; Ghabrial, S.A.; Jiang, D. Extracellular transmission of a DNA mycovirus and its use as a natural fungicide. Proc. Natl. Acad. Sci. USA 2013, 110, 1452–1457. [Google Scholar] [CrossRef]
- Zhang, H.; Xie, J.; Fu, Y.; Cheng, J.; Qu, Z.; Zhao, Z.; Jiang, D. A 2-kb mycovirus converts a pathogenic fungus into a beneficial endophyte for brassica protection and yield enhancement. Mol. Plant 2020, 13, 1420–1433. [Google Scholar] [CrossRef]
- Ying, Z.; Yuanyuan, Z.; Xinru, W.; Yuanyuan, S.; Min, L.; Huijun, X.; Caiyi, W. A Novel Ourmia-Like Mycovirus Confers Hypovirulence-Associated Traits on Fusarium oxysporum. Front. Microbiol. 2020, 11, 3084. [Google Scholar]
- Lemus-Minor, C.G.; Cañizares, M.C.; García-Pedrajas, M.D.; Pérez-Artés, E. Fusarium oxysporum f. sp. dianthi virus 1 accumulation is correlated with changes in virulence and other phenotypic traits of its fungal host. Phytopathology 2018, 108, 957–963. [Google Scholar] [CrossRef] [PubMed]
- Wen, C.; Wan, X.; Zhang, Y.; Du, H.; Wei, C.; Zhong, R.; Zhang, H.; Shi, Y.; Xie, J.; Fu, Y.; et al. Molecular characterization of the first alternavirus identified in Fusarium oxysporum. Viruses 2021, 13, 2026. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.Q. Salvia miltiorrhiza: Chemical and pharmacological review of a medicinal plant. J. Med. Plants Res. 2010, 4, 2813–2820. [Google Scholar]
- Bodah, E.T. Root rot diseases in plants: A review of common causal agents and management strategies. Agric. Res. Technol. 2017, 5, 555661. [Google Scholar]
- Sa, R.; He, S.; Han, D.; Liu, M.; Yu, Y.; Shang, R.; Song, M. Isolation and identification of a new biocontrol bacteria against Salvia miltiorrhiza root rot and optimization of culture conditions for antifungal substance production using response surface methodology. BMC Microbiol. 2022, 22, 231. [Google Scholar] [CrossRef]
- Xie, J.; Wei, D.; Jiang, D.; Fu, Y.; Li, G.; Ghabrial, S.; Peng, Y. Characterization of debilitation-associated mycovirus infecting the plant-pathogenic fungus Sclerotinia sclerotiorum. J. Gen. Virol. 2006, 87, 241–249. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analsis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, K.B.; Holcomb, E.E.; Allscheid, R.L.; Carrington, J.C. Hiding in plain sight: New virus genomes discovered via a systematic analysis of fungal public transcriptomes. PLoS ONE 2019, 14, e0219207. [Google Scholar] [CrossRef] [PubMed]
- Forgia, M.; Isgandarli, E.; Aghayeva, D.N.; Huseynova, I.; Turina, M. Virome characterization of Cryphonectria parasitica isolates from Azerbaijan unveiled a new mymonavirus and a putative new RNA virus unrelated to described viral sequences. Virology 2021, 553, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Pearson, M.N.; Beever, R.E.; Boine, B.; Arthur, K. Mycoviruses of filamentous fungi and their relevance to plant pathology. Mol. Plant. Pathol. 2009, 10, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Chiba, S.; Castón, J.R.; Ghabrial, S.A.; Suzuki, N. ICTV Report Consortium. ICTV virus taxonomy profile: Quadriviridae. J. Gen. Virol. 2018, 99, 1480–1481. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.; Wen, C.; Zhang, Y.; Du, H.; Zhong, R.; Guan, Z.; Wang, M.; Qin, Y.; Wang, F.; Song, L.; et al. The FomYjeF Protein Influences the Sporulation and Virulence of Fusarium oxysporum f. sp. momordicae. Int. J. Mol. Sci. 2023, 24, 7260. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus Name | Accession | Query Cover | Per. Ident | E Value |
|---|---|---|---|---|
| Fusarium mangiferae partitivirus 2 | UBZ25878.1 | 99% | 94.06 | 0 |
| Phomopsis asparagi partitivirus 1 | WEC89325.1 | 99% | 87.94 | 0 |
| Metarhizium brunneum partitivirus 2 | QTC11257.1 | 99% | 87.01 | 0 |
| Colletotrichum truncatum partitivirus 1 | ALF46547.1 | 99% | 87.01 | 0 |
| Plasmopara viticola lesion associated Partitivirus 1 | QHD64804.1 | 99% | 86.64 | 0 |
| Botryotinia fuckeliana partitivirus 1 | YP_001686789.1 | 99% | 84.63 | 0 |
| Ustilaginoidea virens partitivirus 12 | UVX28928.1 | 99% | 84.79 | 0 |
| Virus Name | Accession | Query Cover | Per. Ident | E Value |
|---|---|---|---|---|
| Fusarium mangiferae partitivirus 2 | UBZ25879.1 | 91% | 82.07 | 0 |
| Metarhizium brunneum partitivirus 2 | QTC11258.1 | 99% | 70.3 | 0 |
| Colletotrichum partitivirus 1 | ALD89089.1 | 99% | 71.86 | 0 |
| Sodiomyces alkalinus partitivirus 2 | ATP85066.1 | 99% | 68.14 | 0 |
| Beauveria bassiana partitivirus 2 | CUS18594.1 | 99% | 69.28 | 0 |
| Phomopsis asparagi partitivirus 1 | WEC89326.1 | 99% | 68.14 | 0 |
| Magnaporthe grisea partitivirus 1 | AZT88597.1 | 99% | 65.58 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, L.; Zhong, R.; Guan, Z.; Huang, L.; Wang, G.; Yang, Z.; Shao, K.; Qin, Y.; Wen, C.; Zhao, Y.; et al. Molecular Characterization of the First Partitivirus from a Causal Agent of Salvia miltiorrhiza Dry Rot. J. Fungi 2024, 10, 179. https://doi.org/10.3390/jof10030179
Song L, Zhong R, Guan Z, Huang L, Wang G, Yang Z, Shao K, Qin Y, Wen C, Zhao Y, et al. Molecular Characterization of the First Partitivirus from a Causal Agent of Salvia miltiorrhiza Dry Rot. Journal of Fungi. 2024; 10(3):179. https://doi.org/10.3390/jof10030179
Chicago/Turabian StyleSong, Luyang, Rongrong Zhong, Zhengzhe Guan, Lina Huang, Ganlin Wang, Zhimin Yang, Ke Shao, Yanhong Qin, Caiyi Wen, Ying Zhao, and et al. 2024. "Molecular Characterization of the First Partitivirus from a Causal Agent of Salvia miltiorrhiza Dry Rot" Journal of Fungi 10, no. 3: 179. https://doi.org/10.3390/jof10030179
APA StyleSong, L., Zhong, R., Guan, Z., Huang, L., Wang, G., Yang, Z., Shao, K., Qin, Y., Wen, C., Zhao, Y., & Wang, F. (2024). Molecular Characterization of the First Partitivirus from a Causal Agent of Salvia miltiorrhiza Dry Rot. Journal of Fungi, 10(3), 179. https://doi.org/10.3390/jof10030179