
The Function of Different Subunits of the Molecular Chaperone CCT in the Microsporidium Nosema bombycis: NbCCTζ Interacts with NbCCTα

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Parasite and Host
2.2. Cloning and Bioinformatic Analysis of NbCCTζ
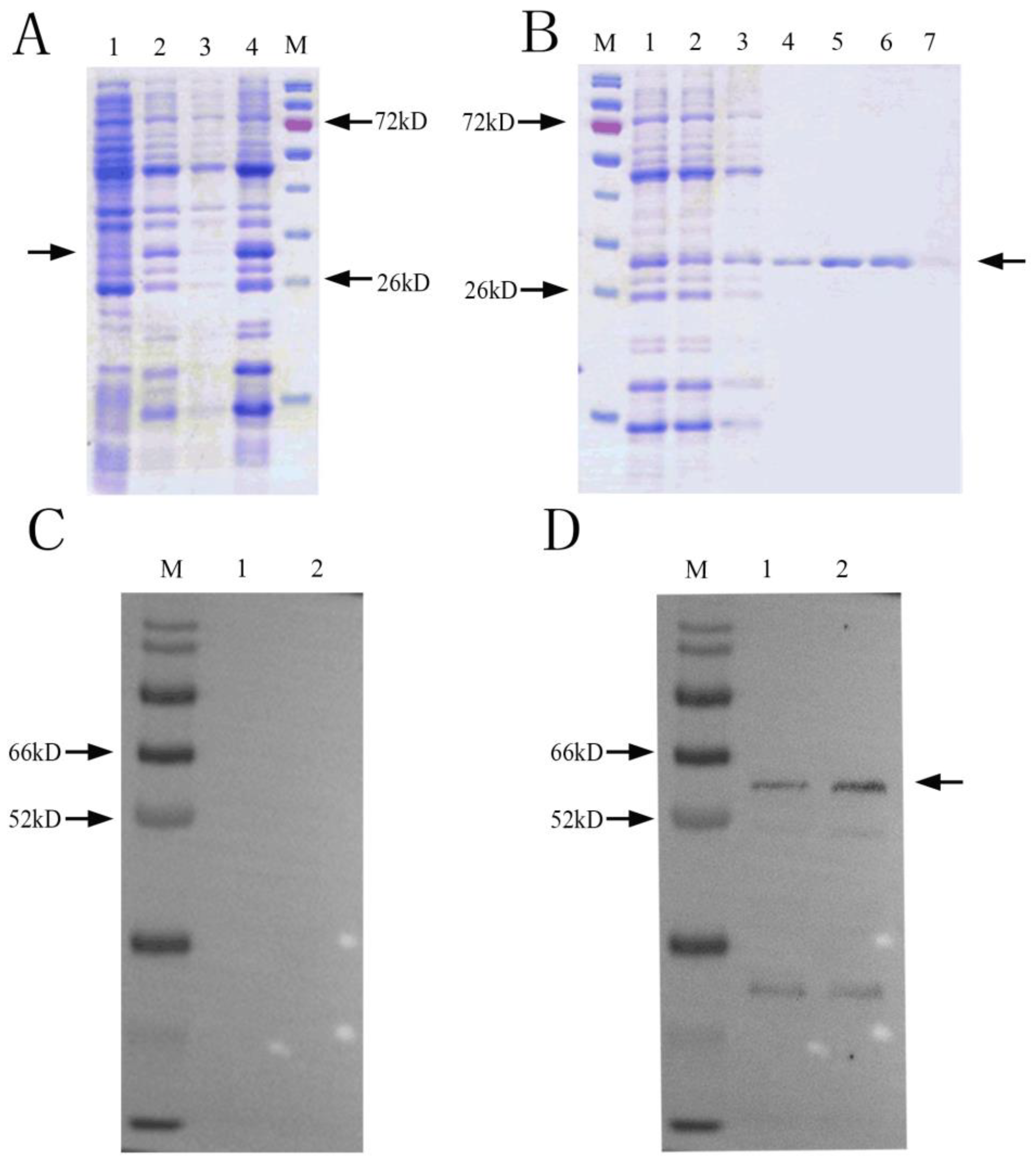
2.3. Expression and Purification of NbCCTζ Recombinant Protein
2.4. Preparation of NbCCTζ Polyclonal Antibody and Western Blot
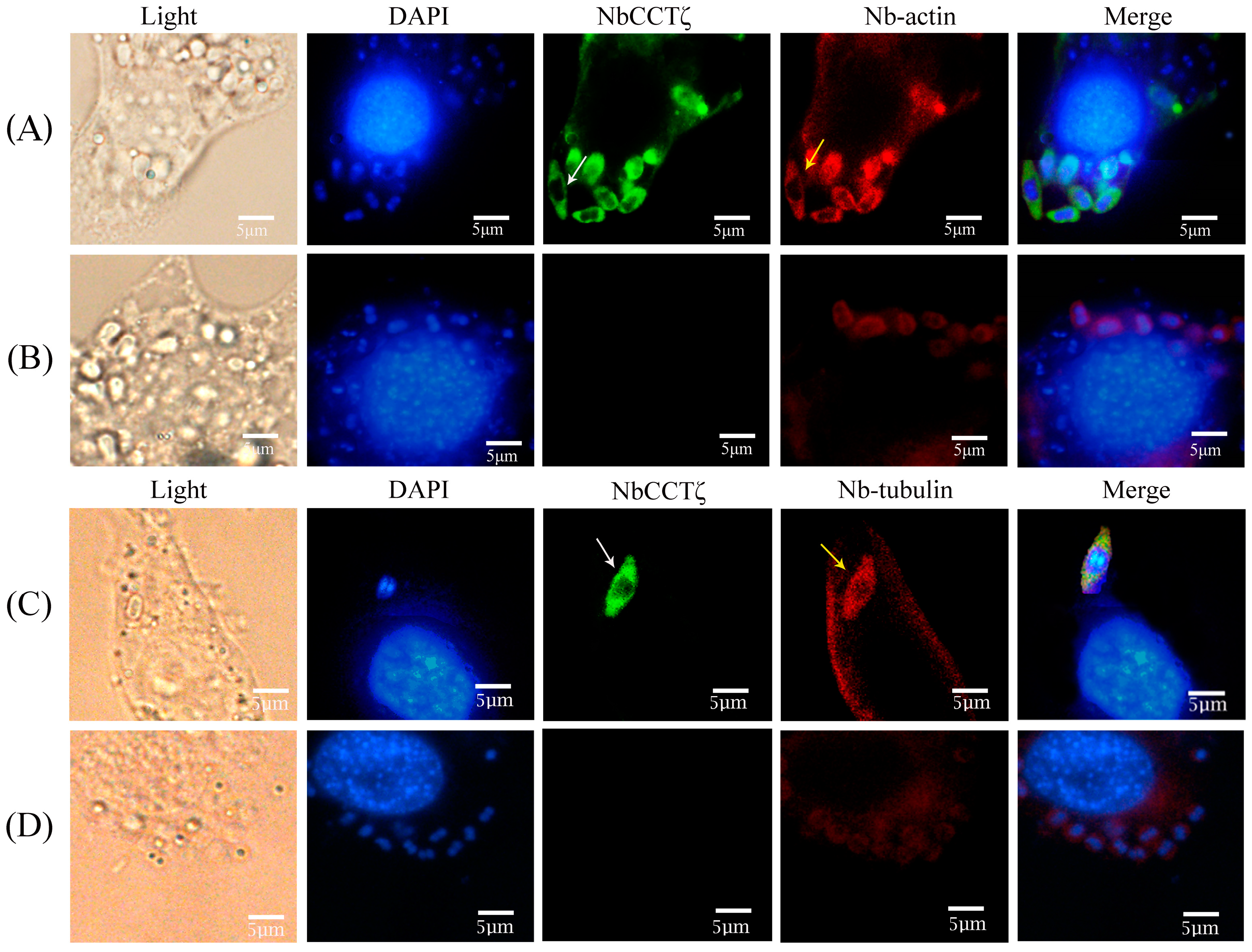
2.5. Immunolocalization of NbCCTζ in N. bombycis
2.6. RNAi
2.7. Yeast Two-Hybrid Assay
3. Results
3.1. Cloning and Expression of NbCCTζ and Western Blot
3.2. Subcellular Localization of NbCCTζ in N. bombycis
3.3. NbCCTζ Interacts with NbCCTα in Yeast Two-Hybrid Assay
3.4. Colocalization of NbCCTζ and NbCCTα
3.5. Knockdown of NbCCTα Downregulated the Expression of NbCCTζ, and the Knockdown of NbCCTζ Downregulated the Expression of NbCCTα
3.6. Knocking down NbCCTζ Suppressed the Proliferation of N. bombycis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guo, R.; Cao, G.; Lu, Y.; Xue, R.; Kumar, D.; Hu, X.; Gong, C. Exogenous gene can be integrated into Nosema bombycis genome by mediating with a non-transposon vector. Parasitol. Res. 2016, 115, 3093–3098. [Google Scholar] [CrossRef] [PubMed]
- Pan, G.; Xu, J.; Li, T.; Xia, Q.; Liu, S.-L.; Zhang, G.; Li, S.; Li, C.; Liu, H.; Yang, L.; et al. Comparative genomics of parasitic silkworm microsporidia reveal an association between genome expansion and host adaptation. BMC Genom. 2013, 14, 186. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Wang, Y.; Wang, L.; Zhou, Z. Molecular and biochemical responses in the midgut of the silkworm, Bombyx mori, infected with Nosema bombycis. Parasites Vectors 2018, 11, 147. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Thomas, J.O.; Chow, R.L.; Lee, G.H.; Cowan, N.J. A cytoplasmic chaperonin that catalyzes beta-actin folding. Cell 1992, 69, 1043–1050. [Google Scholar] [CrossRef]
- Leitner, A.; Joachimiak, L.A.; Bracher, A.; Mönkemeyer, L.; Walzthoeni, T.; Chen, B.; Pechmann, S.; Holmes, S.; Cong, Y.; Ma, B.; et al. The molecular architecture of the eukaryotic chaperonin TRiC/CCT. Structure 2012, 20, 814–825. [Google Scholar] [CrossRef]
- Llorca, O.; Martín-Benito, J.; Gómez-Puertas, P.; Ritco-Vonsovici, M.; Willison, K.R.; Carrascosa, J.L.; Valpuesta, J.M. Analysis of the Interaction between the Eukaryotic Chaperonin CCT and Its Substrates Actin and Tubulin. J. Struct. Biol. 2001, 135, 205–218. [Google Scholar] [CrossRef]
- Hein, M.Y.; Hubner, N.C.; Poser, I.; Cox, J.; Nagaraj, N.; Toyoda, Y.; Gak, I.A.; Weisswange, I.; Mansfeld, J.; Buchholz, F.; et al. A human interactome in three quantitative dimensions organized by stoichiometries and abundances. Cell 2015, 163, 712–723. [Google Scholar] [CrossRef]
- Trinidad, A.G.; Muller, P.A.; Cuellar, J.; Klejnot, M.; Nobis, M.; Valpuesta, J.M.; Vousden, K.H. Interaction of p53 with the CCT complex promotes protein folding and Wild-Type p53 activity. Mol. Cell 2013, 50, 805–817. [Google Scholar] [CrossRef]
- Willison, K.R. The substrate specificity of eukaryotic cytosolic chaperonin CCT. Philos. Trans. R. Soc. B Biol. Sci. 2018, 373, 20170192. [Google Scholar] [CrossRef] [PubMed]
- Grantham, J.; Brackley, K.I.; Willison, K.R. Substantial CCT activity is required for cell cycle progression and cytoskeletal organization in mammalian cells. Exp. Cell Res. 2006, 312, 2309–2324. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.; Zhu, G.; Cui, X.; Kang, L.; Cao, D.; Jiang, Y. Expression of CCT6A mRNA in chicken granulosa cells is regulated by progesterone. Gen. Comp. Endocrinol. 2013, 189, 15–23. [Google Scholar] [CrossRef]
- Génier, S.; Degrandmaison, J.; Moreau, P.; Labrecque, P.; Hébert, T.E.; Parent, J.-L. Regulation of GPCR expression through an interaction with CCT7, a subunit of the CCT/TRiC complex. Mol. Biol. Cell 2016, 27, 3800–3812. [Google Scholar] [CrossRef] [PubMed]
- Kubota, H. Function and regulation of cytosolic molecular chaperone CCT. Vitam. Horm. 2002, 65, 313–331. [Google Scholar] [PubMed]
- Yébenes, H.; Mesa, P.; Muñoz, I.G.; Montoya, G.; Valpuesta, J.M. Chaperonins: Two rings for folding. Trends Biochem. Sci. 2011, 36, 424–432. [Google Scholar] [CrossRef] [PubMed]
- Showalter, A.E.; Martini, A.C.; Nierenberg, D.; Hosang, K.; Fahmi, N.A.; Gopalan, P.; Khaled, A.S.; Zhang, W.; Khaled, A.R. Investigating Chaperonin-Containing TCP-1 subunit 2 as an essential component of the chaperonin complex for tumorigenesis. Sci. Rep. 2020, 10, 798. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Ren, H.; Shao, Y.; Sun, Y.; Zhang, L.; Li, H.; Zhang, X.; Yang, X.; Yu, W.; Fu, J. Chaperonin-containing T-complex protein 1 subunit 8 promotes cell migration and invasion in human esophageal squamous cell carcinoma by regulating α-actin and β-tubulin expression. Int. J. Oncol. 2018, 52, 2021–2030. [Google Scholar] [CrossRef]
- Yin, H.; Miao, X.; Wu, Y.; Wei, Y.; Zong, G.; Yang, S.; Chen, X.; Zheng, G.; Zhu, X.; Guo, Y.; et al. The role of the Chaperonin containing t-complex polypeptide 1, subunit 8 (CCT8) in B-cell non-Hodgkin’s lymphoma. Leuk. Res. 2016, 45, 59–67. [Google Scholar] [CrossRef]
- Dou, L.; Zhang, X. Upregulation of CCT3 promotes cervical cancer progression through FN1. Mol. Med. Rep. 2021, 24, 856. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, P.; Zhang, Z.; Wang, J.; Cheng, Z.; Fan, C. Identification of CCT3 as a prognostic factor and correlates with cell survival and invasion of head and neck squamous cell carcinoma. Biosci. Rep. 2021, 41, BSR20211137. [Google Scholar] [CrossRef]
- Yam, A.Y.; Xia, Y.; Lin, H.-T.J.; Burlingame, A.; Gerstein, M.; Frydman, J. Defining the TRiC/CCT interactome links chaperonin function to stabilization of newly made proteins with complex topologies. Nat. Struct. Mol. Biol. 2008, 15, 1255–1262. [Google Scholar] [CrossRef]
- Van Hove, I.; Verslegers, M.; Hu, T.; Carden, M.; Arckens, L.; Moons, L. A proteomic approach to understand MMP-3-driven developmental processes in the postnatal cerebellum: Chaperonin CCT6A and MAP kinase as contributing factors. Dev. Neurobiol. 2015, 75, 1033–1048. [Google Scholar] [CrossRef]
- Wang, R.; Chen, Y.; Xu, S.; Wei, E.; He, P.; Wang, Q.; Zhang, Y.; Tang, X.; Shen, Z. Ssn6 Interacts with Polar Tube Protein 2 and Transcriptional Repressor for RNA Polymerase II: Insight into Its Involvement in the Biological Process of Microsporidium Nosema bombycis. J. Fungi 2023, 9, 990. [Google Scholar] [CrossRef]
- Sun, F.; Wang, R.; He, P.; Wei, E.; Wang, Q.; Tang, X.; Zhang, Y.; Zhu, F.; Shen, Z. Sar1 Interacts with Sec23/Sec24 and Sec13/Sec31 Complexes: Insight into Its Involvement in the Assembly of Coat Protein Complex II in the Microsporidian Nosema bombycis. Microbiol. Spectr. 2022, 10, e0071922. [Google Scholar] [CrossRef] [PubMed]
- Yao, M.; Wang, R.; Chen, Y.; He, P.; Wei, E.; Zhu, F.; Wang, Q.; Zhang, Y.; Tang, X.; Shen, Z. Identification and subcellular localization analysis of CCTα in microsporidian Nosema bombycis. Infect. Genet. Evol. 2022, 102, 105309. [Google Scholar] [CrossRef]
- Silver, L.M.; Artzt, K.; Bennett, D. A major testicular cell protein specified by a mouse T/t complex gene. Cell 1979, 17, 275–284. [Google Scholar] [CrossRef]
- Hartl, F.U.; Bracher, A.; Hayer-Hartl, M. Molecular chaperones in protein folding and proteostasis. Nature 2011, 475, 324–332. [Google Scholar] [CrossRef] [PubMed]
- Heinz, E.; Williams, T.A.; Nakjang, S.; Noël, C.J.; Swan, D.C.; Goldberg, A.V.; Harris, S.R.; Weinmaier, T.; Markert, S.; Becher, D.; et al. The genome of the obligate intracellular parasite Trachipleistophora hominis: New insights into microsporidian genome dynamics and reductive evolution. PLoS Pathog. 2012, 8, e1002979. [Google Scholar] [CrossRef]
- Barandun, J.; Hunziker, M.; Vossbrinck, C.R.; Klinge, S. Evolutionary compaction and adaptation visualized by the structure of the dormant microsporidian ribosome. Nat. Microbiol. 2019, 4, 1798–1804. [Google Scholar] [CrossRef] [PubMed]
- Jespersen, N.; Ehrenbolger, K.; Winiger, R.R.; Svedberg, D.; Vossbrinck, C.R.; Barandun, J. Structure of the reduced microsporidian proteasome bound by PI31-like peptides in dormant spores. Nat. Commun. 2022, 13, 6962. [Google Scholar] [CrossRef]
- Roobol, A.; Sahyoun, Z.P.; Carden, M.J. Selected subunits of the cytosolic chaperonin associate with microtubules assembled in vitro. J. Biol. Chem. 1999, 274, 2408–2415. [Google Scholar] [CrossRef]
- Leroux, M.R.; Hartl, F. Protein folding: Versatility of the cytosolic chaperonin TRiC/CCT. Curr. Biol. 2000, 10, R260–R264. [Google Scholar] [CrossRef] [PubMed]
- Nakjang, S.; Williams, T.A.; Heinz, E.; Watson, A.K.; Foster, P.G.; Sendra, K.M.; Heaps, S.E.; Hirt, R.P.; Embley, T.M. Reduction and expansion in microsporidian genome evolution: New insights from comparative genomics. Genome Biol. Evol. 2013, 5, 2285–2303. [Google Scholar] [CrossRef] [PubMed]
- Cali, A.; Takvorian, P.M. Developmental morphology and life cycles of the microsporidia. In Microsporidia: Pathogens of Opportunity; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2014; pp. 71–133. [Google Scholar]
- Cong, Y.; Baker, M.L.; Jakana, J.; Woolford, D.; Miller, E.J.; Reissmann, S.; Kumar, R.N.; Redding-Johanson, A.M.; Batth, T.S.; Mukhopadhyay, A.; et al. 4.0-A resolution cryo-EM structure of the mammalian chaperonin TRiC/CCT reveals its unique subunit arrangement. Proc. Natl. Acad. Sci. USA 2010, 107, 4967–4972. [Google Scholar] [CrossRef] [PubMed]
- Dekker, C.; Roe, S.M.; McCormack, E.A.; Beuron, F.; Pearl, L.H.; Willison, K.R. The crystal structure of yeast CCT reveals intrinsic asymmetry of eukaryotic cytosolic chaperonins. EMBO J. 2011, 30, 3078–3090. [Google Scholar] [CrossRef]
- Wagner, C.T.; Lu, I.Y.; Hoffman, M.H.; Sun, W.Q.; Trent, J.D.; Connor, J. T-complex Polypeptide-1 interacts with the erythrocyte cytoskeleton in response to elevated temperatures. J. Biol. Chem. 2004, 279, 16223–16228. [Google Scholar] [CrossRef]
- Qi, J.; Zhu, F.; Shao, L.; Chen, Y.; Li, J.; He, P.; Shang, R.; Sun, F.; Wang, Q.; Zhang, Y.; et al. CCTδ colocalizes with actin and β-tubulin: Insight into its involvement in the cytoskeleton formation of the intracellular parasite Nosema bombycis. J. Invertebr. Pathol. 2021, 184, 107646. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequences | |
|---|---|---|
| NbCCTζ (RNAi) | Sense Antisense | GCUAUUAGAGCCUCUCUUATT (5′-3′) UAAGAGAGGCUCUAAUAGCTT (5′-3′) |
| Negative control (RNAi) | Sense Antisense | UUCUCCGAACGUGUCACGUTT (5′-3′) ACGUGACACGUUCGGAGAATT (5′-3′) |
| NbCCTζ (RT-qPCR) | Forward Reverse | AGCCAGAGCTATTGCTGCTTTTGG (5′-3′) GAATACCAGCCTTGGCAAGGATTTC (5′-3′) |
| NbCCTα (RT-qPCR) | Forward Reverse | GGTGGGGCTATTGAAGTCGC (5′-3′) CCAAACCCGCATTTATTGCCA (5′-3′) |
| BmGAPDH (RT-qPCR) | Forward Reverse | TTCATGCCACAACTGCTACA (5′-3′) AGTCAGCTTGCCATTAAGAG (5′-3′) |
| SSrRNA (RT-qPCR) | Forward Reverse | TCGAGTGCCAGCAGCCGCGG (5′-3′) CGATCCTCTAGCTTACGTCC (5′-3′) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, S.; Chen, Y.; Qi, J.; Wang, R.; Wei, E.; Wang, Q.; Zhang, Y.; Tang, X.; Shen, Z. The Function of Different Subunits of the Molecular Chaperone CCT in the Microsporidium Nosema bombycis: NbCCTζ Interacts with NbCCTα. J. Fungi 2024, 10, 229. https://doi.org/10.3390/jof10030229
Xu S, Chen Y, Qi J, Wang R, Wei E, Wang Q, Zhang Y, Tang X, Shen Z. The Function of Different Subunits of the Molecular Chaperone CCT in the Microsporidium Nosema bombycis: NbCCTζ Interacts with NbCCTα. Journal of Fungi. 2024; 10(3):229. https://doi.org/10.3390/jof10030229
Chicago/Turabian StyleXu, Sheng, Ying Chen, Jingru Qi, Runpeng Wang, Erjun Wei, Qiang Wang, Yiling Zhang, Xudong Tang, and Zhongyuan Shen. 2024. "The Function of Different Subunits of the Molecular Chaperone CCT in the Microsporidium Nosema bombycis: NbCCTζ Interacts with NbCCTα" Journal of Fungi 10, no. 3: 229. https://doi.org/10.3390/jof10030229
APA StyleXu, S., Chen, Y., Qi, J., Wang, R., Wei, E., Wang, Q., Zhang, Y., Tang, X., & Shen, Z. (2024). The Function of Different Subunits of the Molecular Chaperone CCT in the Microsporidium Nosema bombycis: NbCCTζ Interacts with NbCCTα. Journal of Fungi, 10(3), 229. https://doi.org/10.3390/jof10030229