Endophytic Fungi: A Source of Potential Antifungal Compounds




Abstract
:1. Introduction
2. Medicinal Plants
2.1. Compounds Produced by Coelomycetes
2.2. Compounds Produced by Ascomycetes
2.3. Compounds Produced by Hyphomycetes
2.4. Compounds Produced by Basidiomycetes
3. Antifungal Potential of Volatile Organic Compounds (VOCs) from Endophytic Fungi
4. Methods Used for Activation of Silent Biosynthetic Genes
4.1. Epigenetic Modification
4.2. The Co-Culture Strategy
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hawksworth, D.L. The fungal dimension of biodiversity: Magnitude, significance, and conservation. Mycol. Res. 1991, 95, 641–655. [Google Scholar] [CrossRef]
- Schulz, B.; Boyle, C.; Draeger, S.; Römmert, A.K.; Krohn, K. Endophytic fungi: A source of novel biologically active secondary metabolites. Mycol. Res. 2002, 106, 996–1004. [Google Scholar] [CrossRef]
- Nicoletti, R.; Fiorentino, A. Plant bioactive metabolites and drugs produced by endophytic fungi of Spermatophyta. Agriculture 2015, 5, 918–970. [Google Scholar] [CrossRef]
- Strobel, G.; Daisy, B. Bioprospecting for microbial endophytes and their natural products. Microbiol. Mol. Biol. Rev. 2003, 67, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Gunatilaka, A.A.L. Natural products from plant-associated microorganisms: Distribution, structural diversity, bioactivity, and implications of their occurrence. J. Nat. Prod. 2006, 69, 509–526. [Google Scholar] [CrossRef] [PubMed]
- Kharwar, R.N.; Mishra, A.; Gond, S.K.; Stierle, A.; Stierle, D. Anticancer compounds derived from fungal endophytes: Their importance and future challenges. Nat. Prod. Rep. 2011, 28, 1208–1228. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, S.K.; Verekar, S.A. Fungal endophytes: A potential source of antifungal compounds. Front. Biosci. 2012, E4, 2045–2070. [Google Scholar] [CrossRef]
- Deshmukh, S.K.; Verekar, S.A.; Bhave, S.V. Endophytic fungi: A reservoir of antibacterials. Front. Microbiol. 2015, 5, 715. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Li, G.; Lou, H.X. Structural Diversity and Biological Activities of Novel Secondary metabolites from Endophytes. Molecules 2018, 23, 646. [Google Scholar] [CrossRef] [PubMed]
- Carroll, G. Fungal endophytes in stems and leaves: From latent pathogens to mutualistic symbionts. Ecology 1988, 69, 2–9. [Google Scholar] [CrossRef]
- Hallmann, J.; Sikora, R. Toxicity of fungal endophytic secondary metabolites to plant parasitic nematodes and soil borne plant pathogenic fungi. Eur. J. Plant Pathol. 1996, 102, 155–162. [Google Scholar] [CrossRef]
- Sturz, A.V.; Nowak, J. An endophytic community of rhizobacteria and the strategies requires to create yield enhancing associations with crops. Appl. Soil Ecol. 2000, 15, 183–190. [Google Scholar] [CrossRef]
- Azevedo, J.L.; Araujo, W.L. Diversity and applications of endophytic fungi isolated from tropical plants. In Fungi Multifaceted Microbes; Ganguli, B.N., Deshmukh, S.K., Eds.; Anamaya: New Delhi, India, 2007; pp. 189–207. [Google Scholar]
- Deshmukh, S.K.; Prakash, V.; Ranjan, N. Recent advances in the discovery of bioactive metabolites from Pestalotiopsis. Phytochem. Rev. 2017, 16, 883–920. [Google Scholar] [CrossRef]
- Xu, D.; Zhang, B.Y.; Yang, X.L. Antifungal monoterpene derivatives from the plant endophytic fungus Pestalotiopsis foedan. Chem. Biodivers. 2016, 13, 1422–1425. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.S.; Jia, M.; Chen, L.; Zhu, B.; Dong, H.X.; Si, J.P.; Peng, W.; Han, T. Cytotoxic and antifungal constituents isolated from the metabolites of endophytic fungus DO14 from Dendrobium officinale. Molecules 2016, 21, 14. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Liu, X.; Guo, L.; Che, Y.; Liu, L. 2H-Pyran-2-one and2H-Furan-2-one derivatives from the plant endophytic fungus Pestalotiopsis fici. Chem. Biodivers. 2013, 10, 2007–2013. [Google Scholar] [CrossRef] [PubMed]
- Subban, K.; Subramani, R.; Muthumary, J. A novel antibacterial and antifungal phenolic compound from the endophytic fungus Pestalotiopsis mangiferae. Nat. Prod. Res. 2013, 27, 1445–1449. [Google Scholar] [CrossRef] [PubMed]
- Chapla, V.M.; Zeraik, M.L.; Ximenes, V.F.; Zanardi, L.M.; Lopes, M.N.; Cavalheiro, A.J.; Silva, D.H.S.; Young, M.C.M.; da Fonseca, L.M.; Bolzani, V.S.; et al. Bioactive secondary metabolites from Phomopsis sp., an endophytic fungus from Senna spectabilis. Molecules 2014, 19, 6597–6608. [Google Scholar]
- Wu, S.H.; Huang, R.; Miao, C.P.; Chen, Y.W. Two new steroids from an endophytic fungus Phomopsis sp. Chem. Biodivers. 2013, 10, 1276–1283. [Google Scholar] [CrossRef] [PubMed]
- Tanney, J.B.; McMullin, D.R.; Green, B.D.; Miller, J.D.; Seifert, K.A. Production of antifungal and anti-insectan metabolites by the Picea endophyte Diaporthe maritima sp. nov. Fungal Biol. 2016, 120, 1448–1457. [Google Scholar] [CrossRef] [PubMed]
- Rai, M.; Deshmukh, P.; Gade, A.; Ingle, A.; Kövics, G.J.; Irinyi, L. Phoma Saccardo: Distribution, secondary metabolite production and biotechnological applications. Crit. Rev. Microbiol. 2009, 35, 182–196. [Google Scholar] [CrossRef] [PubMed]
- Hussain, H.; Kock, I.; Al-Harras, A.; Al-Rawahi, A.; Abbas, G.; Green, I.R.; Shah, A.; Badshah, A.; Saleem, M.; Draeger, S.; et al. Antimicrobial chemical constituents from endophytic fungus Phoma sp. Asian. Pac. J. Trop. Med. 2014, 7, 699–702. [Google Scholar] [CrossRef]
- Mousa, W.K.; Schwan, A.; Davidson, J.; Auzanneau, F.I.; Strange, P.; Liu, H.; Zhou, T.; Raizada, M.N. An endophytic fungus isolated from finger millet (Eleusine coracana) produces anti-fungal natural products. Front. Microbiol. 2015, 6, 1157. [Google Scholar] [CrossRef] [PubMed]
- Lai, D.; Wang, A.; Cao, Y.; Zhou, K.; Mao, Z.; Dong, X.; Tian, J.; Xu, D.; Dai, J.; Peng, Y.; et al. Bioactivedibenzo-α-pyrone derivatives from the endophytic fungus Rhizopycnis vagum Nitaf22. J. Nat. Prod. 2016, 79, 2022–2031. [Google Scholar] [CrossRef] [PubMed]
- Hussain, H.; Root, N.; Jabeen, F.; Al-Harrasi, A.; Ahmad, M.; Mabood, F.; Hassan, Z.; Shah, A.; Green, I.R.; Schulz, B.; et al. Microsphaerol and seimatorone: Two new compounds isolated from the endophytic fungi, Microsphaeropsis sp. and Seimatosporium sp. Chem. Biodivers. 2015, 12, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Chapla, V.M.; Zeraik, M.L.; Leptokarydis, I.H.; Silva, G.H.; Bolzani, V.S.; Young, M.C.M.; Pfenning, L.H.; Araujo, A.R. Antifungal compounds produced by Colletotrichum gloeosporioides, an endophytic fungus from Michelia champaca. Molecules 2014, 19, 19243–19252. [Google Scholar] [CrossRef] [PubMed]
- Hussain, H.; Root, N.; Jabeen, F.; Al-Harras, A.; Al-Rawahi, A.; Ahmad, M.; Hassan, Z.; Abba, G.; Mabood, F.; Shah, A.; et al. Seimatoric acid and colletonoic acid: Two new compounds from the endophytic fungi, Seimatosporium sp. and Colletotrichum sp. Chin. Chem. Lett. 2014, 25, 1577–1579. [Google Scholar] [CrossRef]
- Sun, P.; Huo, J.; Kurtan, T.; Mandi, A.; Antus, S.; Tang, H.; Draeger, S.; Schulz, B.; Hussain, H.; Krohn, K.; et al. Structural and stereochemical studies of hydroxyanthraquinone derivatives from the endophytic fungus Coniothyrium sp. Chirality 2013, 25, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Macias-Rubalcava, M.L.; Sanchez-Fernandez, R.E. Secondary metabolites of endophytic Xylaria species with potential applications in medicine and agriculture. World J. Microbiol. Biotechnol. 2017, 33, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Song, F.; Wu, S.H.; Zhai, Y.Z.; Xuan, Q.C.; Wang, T. Secondary metabolites from the genus Xylaria and their bioactivities. Chem. Biodivers. 2014, 11, 673–694. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.H.; He, J.; Li, X.N.; Huang, R.; Song, F.; Chen, Y.W.; Miao, C.P. Guaianesesquiterpenes and isopimaranediterpenes from an endophytic fungus Xylaria sp. Phytochemistry 2014, 105, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Xie, X.S.; Fang, X.W.; Ma, K.X.; Wu, S.H. Five new guaianesesquiterpenes from the endophytic fungus Xylaria sp. YM 311647 of Azadirachta indica. Chem. Biodivers. 2015, 12, 1281–1286. [Google Scholar] [CrossRef] [PubMed]
- Baraban, E.G.; Morin, J.B.; Phillips, G.M.; Phillips, A.J.; Strobel, S.A.; Handelsman, J. Xyolide, a bioactive nonenolide from an Amazonian endophytic fungus, Xylaria feejeensis. Tetrahedron Lett. 2013, 54, 4058–4060. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Xiao, J.; Sun, Q.Q.; Qin, J.C.; Pescitelli, G.; Gao, J.M. Characterization of cytochalasins from the endophytic Xylaria sp. and their biological functions. J. Agric. Food. Chem. 2014, 62, 10962–10969. [Google Scholar] [CrossRef] [PubMed]
- Rukachaisirikul, V.; Buadam, S.; Sukpondma, Y.; Phongpaichit, S.; Sakayaroj, J.; Hutadilok-Towatana, N. Indanone and mellein derivatives from the Garcinia-derived fungus Xylaria sp. PSU-G12. Phytochem. Lett. 2013, 6, 135–138. [Google Scholar] [CrossRef]
- Sica, V.P.; Rees, E.R.; Tchegnon, E.; Bardsley, R.H.; Raja, H.A.; Oberlies, N.H. Spatial and temporal profiling of griseofulvin production in Xylaria cubensis using mass spectrometry mapping. Front. Microbiol. 2016, 7, 544. [Google Scholar] [CrossRef] [PubMed]
- Richardson, S.N.; Walker, A.K.; Nsiama, T.K.; McFarlane, J.; Sumarah, M.W.; Ibrahim, A.; Miller, J.D. Griseofulvin producing Xylaria endophytes of Pinus strobus and Vaccinium angustifolium: Evidence for a conifer-understory species endophyte ecology. Fungal Ecol. 2014, 11, 107–113. [Google Scholar] [CrossRef]
- Zhang, Q.; Li, H.Q.; Zong, S.C.; Gao, J.M.; Zhang, A.L. Chemical and bioactive diversities of the genus Chaetomium secondary metabolites. Mini-Rev. Med. Chem. 2012, 12, 127–148. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Y.; Yan, X.; Fang, M.J.; Wu, Z.; Wang, D.; Qiu, Y.K. Two new cytochalasan derivatives from Chaetomium globosum SNSHI-5, a fungus derived from extreme environment. Nat. Prod. Res. 2017, 31, 1669–1675. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.S.; Zhang, Y.Y.; Yan, W.; Cao, L.L.; Xiao, Y.; Ye, Y.H.; Zhao, S.S.; Zhang, Y.Y.; Yan, W.; Cao, L.L.; et al. Chaetomium globosum CDW7, a potential biological control strain and its antifungal metabolites. FEMS Microbiol. Lett. 2017, 364. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Yang, X.; Yang, Y.; Duang, R.; Chen, G.; Li, X.; Li, Q.; Qin, S.; Li, S.; Zhao, L.; et al. Anti-phytopathogen, multi-target acetylcholinesterase inhibitory and antioxidant activities of metabolites from endophytic Chaetomium globosum. Nat. Prod. Res. 2016, 30, 2616–2619. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, Y.Y.; Ding, D.D.; Yu, S.P.; Wang, L.W. A study on the secondary metabolites of endophytic fungus Chaetomium cupreum ZJWCF079 in Macleaya cordata. Health Res. 2013, 33, 94–96. [Google Scholar]
- Zhang, G.; Zhang, Y.; Qin, J.; Qu, X.; Liu, J.; Li, X.; Pan, H. Antifungal metabolites produced by Chaetomiumglobosum No.04, an endophytic fungus isolated from Ginkgo biloba. Indian J. Microbiol. 2013, 53, 175–180. [Google Scholar] [PubMed]
- Chen, Y.M.; Yang, Y.H.; Li, X.N.; Zou, C.; Zhao, P.J. Diterpenoids from the endophytic fungus Botryosphaeria sp. P483 of the Chinese herbal medicine Huperzia serrata. Molecules 2015, 20, 16924–16932. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Zhang, Q.; Gao, Y.Q.; Tang, J.J.; Zhang, A.L.; Gao, J.M. Secondary metabolites from the endophytic Botryosphaeria dothidea of Melia azedarach and their antifungal, antibacterial, antioxidant, and cytotoxic activities. J. Agric. Food Chem. 2014, 62, 3584–3590. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.B.; de Oliveira, D.M.; Hughes, A.F.S.; Kohlhoff, M.; Vieira, M.L.A.; Martins Vaz, A.B.; Ferreira, M.C.; Carvalho, C.R.; Rosa, L.H.; Rosa, C.A.; et al. Endophytic fungal compounds active against Cryptococcus neoformans and C. gattii. J. Antibiot. 2015, 68, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.; de Sá, N.P.; Borelli, B.M.; Rosa, C.A.; Barbeira, P.J.S.; Cota, B.B.; Johann, S. Antifungal activity of eicosanoic acids isolated from the endophytic fungus Mycosphaerella sp. against Cryptococcus neoformans and C. gattii. Microb. Pathog. 2016, 100, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Li, T.X.; Yang, M.H.; Wang, X.B.; Wang, Y.; Kong, L.Y. Synergistic antifungal meroterpenes and dioxolanone derivatives from the endophytic fungus Guignardia sp. J. Nat. Prod. 2015, 78, 2511–2520. [Google Scholar] [CrossRef] [PubMed]
- Mao, Z.; Lai, D.; Liu, X.; Fu, X.; Meng, J.; Wang, A.; Wang, X.; Sun, W.; Liu, Z.L.; Zhou, L.; et al. Dibenzo-α-pyrones: A new class of larvicidal metabolites against Aedes aegypti from the endophytic fungus Hyalodendriella sp. Ponipodef12. Pest Manag. Sci. 2017, 73, 1478–1485. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Mao, Z.; Lou, J.; Xu, L.; Zhong, L.; Peng, Y.; Zhou, L.; Wang, M. Benzopyranones from the endophytic fungus Hyalodendriella sp. Ponipodef12 and their bioactivities. Molecules 2012, 17, 11303–11314. [Google Scholar] [CrossRef] [PubMed]
- Rosseto, P.; Costa, A.T.; Polonio, J.C.; da Silva, A.A.; Pamphile, J.A.; Azevedo, J.L. Investigation of mycoviruses in endophytic and phytopathogenic strains of Colletotrichum from different hosts. Genet. Mol. Res. 2016, 15, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, G.; Zhang, Y.; Zheng, B.; Zhang, C.; Wang, L. Isolation and identification of an endophytic fungus Pezicula sp. in Forsythia viridissima and its secondary metabolites. World. J. Microbiol. Biotechnol. 2014, 30, 2639–2644. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.L.; Zhang, Y.Y.; Liu, Y.J.; Yang, T.T.; Zhang, J.L.; Zhang, Z.G.; Shen, L.; Liu, J.Y.; Ye, Y.H. Anti-phytopathogenic activity of sporothriolide, a metabolite from endophyte Nodulisporium sp. A21 in Ginkgo biloba. Pestic. Biochem. Physiol. 2016, 129, 7–13. [Google Scholar] [CrossRef] [PubMed]
- McMullin, D.R.; Green, B.D.; Miller, J.D. Antifungal sesquiterpenoids and macrolides from an endophytic Lophodermium species of Pinus strobus. Phytochem. Lett. 2015, 14, 148–152. [Google Scholar] [CrossRef]
- Li, R.; Chen, S.; Niu, S.; Guo, L.; Yin, J.; Che, Y. Exserolides A-F, new isocoumarin derivatives from the plant endophytic fungus Exserohilum sp. Fitoterapia 2014, 96, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, C.R.; Wedge, D.E.; Cantrell, C.L.; Silva-Hughes, A.F.; Pan, Z.; Moraes, R.M.; Madoxx, V.L.; Rosa, L.H. Molecular phylogeny, diversity, and bioprospecting of endophytic fungi associated with wild ethnomedicinal North American plant Echinacea purpurea (Asteraceae). Chem. Biodivers. 2016, 13, 918–930. [Google Scholar] [CrossRef] [PubMed]
- Burgess, K.M.N.; Ibrahim, A.; Soerensen, D.; Sumarah, M.W. Trienylfuranol A and trienylfuranone A.B: Metabolites isolated from an endophytic fungus, Hypoxylon submonticulosum, in the raspberry Rubus idaeus. J. Antibiot. 2017, 70, 721–725. [Google Scholar] [CrossRef] [PubMed]
- Nalli, Y.; Mirza, D.N.; Wani, Z.A.; Wadhwa, B.; Mallik, F.A.; Raina, C.; Chaubey, A.; Riyaz-Ul-Hassan, S.; Ali, A. Phialomustin A-D, new antimicrobial and cytotoxic metabolites from an endophytic fungus, Phialophora mustea. RSC Adv. 2015, 5, 95307–95312. [Google Scholar] [CrossRef]
- Hussain, H.; Jabeen, F.; Krohn, K.; Al-Harras, A.; Ahmad, M.; Mabood, F.; Shah, A.; Badshah, A.; Rehman, N.U.; Green, I.R.; et al. Antimicrobial activity of two mellein derivatives isolated from an endophytic fungus. Med. Chem. Res. 2015, 24, 2111–2114. [Google Scholar] [CrossRef]
- Hussain, H.; Kliche-Spory, C.; Al-Harrasi, A.; Al-Rawahi, A.; Abbas, G.; Green, I.R.; Schulz, B.; Krohn, K.; Shah, A. Antimicrobial constituents from three endophyticfungi. Asian Pac. J. Trop. Med. 2014, 7, S224–S227. [Google Scholar] [CrossRef]
- Shan, T.; Tian, J.; Wang, X.; Mou, Y.; Mao, Z.; Lai, D.; Dai, J.; Peng, Y.; Zhou, L.; Wang, M. Bioactive spirobisnaphthalenes from the endophytic fungus Berkleasmium sp. J. Nat. Prod. 2014, 77, 2151–2160. [Google Scholar] [CrossRef] [PubMed]
- Siriwach, R.; Kinoshita, H.; Kitani, S.; Igarashi, Y.; Pansuksan, K.; Panbangred, W.; Nihira, T. Bipolamides A and B, triene amides isolated from the endophytic fungus Bipolaris sp. MU34. J. Antibiot. 2014, 67, 167–170. [Google Scholar] [CrossRef] [PubMed]
- Phaopongthai, J.; Wiyakrutta, S.; Meksuriyen, D.; Sriubolmas, N.; Suwanborirux, K. Azole-synergistic anti-candidal activity of altenusin, a biphenyl metabolite of the endophytic fungus Alternaria alternata isolated from Terminalia chebula Retz. J. Microbiol. 2013, 51, 821–828. [Google Scholar] [CrossRef] [PubMed]
- Johann, S.; Rosa, L.H.; Rosa, C.A.; Perez, P.; Cisalpino, P.S.; Zani, C.L.; Cota, B.B. Antifungal activity of altenusin isolated from the endophytic fungus Alternaria sp. against the pathogenic fungus Paracoccidioides brasiliensis. Rev. Iberoam. Micol. 2012, 29, 205–209. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Radwan, M.M.; Tarawneh, A.H.; Gao, J.; Wedge, D.E.; Rosa, L.H.; Cutler, H.G.; Cutler, S.J. Antifungal activity against plant pathogens of metabolites from the endophytic fungus Cladosporium cladosporioides. J. Agric. Food Chem. 2013, 61, 4551–4555. [Google Scholar] [CrossRef] [PubMed]
- Talontsi, F.M.; Dittrich, B.; Schueffler, A.; Sun, H.; Laatsch, H. Epicoccolides: Antimicrobial and antifungal polyketides from an endophytic fungus Epicoccum sp. associated with Theobroma cacao. Eur. J. Org. Chem. 2013, 2013, 3174–3180. [Google Scholar] [CrossRef]
- Silva-Hughes, A.F.; Carvalho, C.R.; Wedge, D.E.; Cantrell, C.L.; Pan, Z.; Moraes, R.M.; Madoxx, V.L.; Rosa, L.H. Diversity and antifungal activity of the endophytic fungi associated with the native medicinal cactus Opuntia humifusa (Cactaceae) from the United States. Microbiol. Res. 2015, 175, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Pang, X.J.; Xu, L.L.; Zhao, T.; Long, X.Y.; Zhang, Q.Y.; Qin, H.L.; Yang, D.F.; Yang, X.L. Two new alkylated furan derivatives with antifungal and antibacterial activities from the plant endophytic fungus Emericella sp. XL029. Nat. Prod. Res. 2017, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Mousa, W.K.; Schwan, A.L.; Raizada, M.N. Characterization of antifungal natural products isolated from endophytic fungi of finger millet (Eleusine coracana). Molecules 2016, 21, 1171. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Yang, Y.B.; Chen, J.L.; Miao, C.P.; Wang, Q.; Zhou, H.; Chen, Y.W.; Li, Y.Q.; Ding, Z.T.; Zhao, L.X. Koninginins N-Q, polyketides from the endophytic fungus Trichoderma koningiopsis Harbored in Panax notoginseng. Nat. Prod. Bioprospect. 2016, 6, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Yang, Y.; Miao, C.P.; Zheng, Y.K.; Chen, J.L.; Chen, Y.W.; Xu, L.H.; Guang, H.L.; Ding, Z.T.; Zhao, L.X. Koningiopisins A-H, polyketides with synergistic antifungal activities from the endophytic fungus Trichoderma koningiopsis. Planta Medica 2016, 82, 371–376. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Xu, J.; Li, F.; Xu, L.; Li, C. A New antifungal isocoumarin from the endophytic fungus Trichoderma sp. 09 of Myoporum bontioides A. Gray. Pharmacogn. Mag. 2016, 12, 259–261. [Google Scholar] [PubMed]
- Shentu, X.; Zhan, X.; Ma, Z.; Yu, X.; Zhang, C. Antifungal activity of metabolites of the endophytic fungus Trichoderma brevicompactum from garlic. Braz. J. Microbiol. 2014, 45, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Li, Q.L.; Yang, Y.B.; Liu, K.; Miao, C.P.; Zhao, L.X.; Ding, Z.T. Koninginins R-S from the endophytic fungus Trichoderma koningiopsis. Nat. Prod. Res. 2017, 31, 835–839. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.R.M.; Elkhayat, E.S.; Mohamed, G.A.; Khedr, A.I.M.; Fouad, M.A.; Kotb, M.H.R.; Ross, S.A. Aspernolides F and G, new butyrolactones from the endophytic fungus Aspergillus terreus. Phytochem. Lett. 2015, 14, 84–90. [Google Scholar] [CrossRef]
- Xiao, J.; Zhang, Q.; Gao, Y.Q.; Shi, X.W.; Gao, J.M. Antifungal and antibacterial metabolites from an endophytic Aspergillus sp. associated with Melia azedarach. Nat. Prod. Res. 2014, 28, 1388–1392. [Google Scholar] [PubMed]
- Budhiraja, A.; Nepali, K.; Sapra, S.; Gupta, S.; Kumar, S.; Dhar, K.L. Bioactive metabolites from an endophytic fungus of Aspergillus species isolated from seeds of Gloriosa superba Linn. Med. Chem. Res. 2013, 22, 323–329. [Google Scholar] [CrossRef]
- Ma, Y.M.; Qiao, K.; Kong, Y.; Li, M.Y.; Guo, L.X.; Miao, Z.; Fan, C. A new isoquinolone alkaloid from an endophytic fungus R22 of Nerium indicum. Nat. Prod. Res. 2017, 31, 951–958. [Google Scholar] [CrossRef] [PubMed]
- Kajula, M.; Ward, J.M.; Turpeinen, A.; Tejesvi, M.V.; Hokkanen, J.; Tolonen, A.; Hakkanen, H.; Picart, P.; Ihalainen, J.; Sahl, H.G.; et al. Bridged epipolythiodiketopiperazines from Penicillium raciborskii, an endophytic fungus of Rhododendron tomentosum Harmaja. J. Nat. Prod. 2016, 79, 685–690. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.R.M.; Abdallah, H.M.; Elkhayat, E.S.; Al Musayeib, N.M.; Asfour, H.Z.; Zayed, M.F.; Mohamed, G.A. Fusaripeptide A: New antifungal and anti-malarial cyclodepsipeptide from the endophytic fungus Fusarium sp. J. Asian Nat. Prod. Res. 2018, 20, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.R.M.; Elkhayat, E.S.; Mohamed, G.A.A.; Fat’hi, S.M.; Ross, S.A. Fusarithioamide A, a new antimicrobial and cytotoxic benzamide derivative from the endophytic fungus Fusarium chlamydosporium. Biochem. Biophys. Res. Commun. 2016, 479, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.A.; Ma, Y.M.; Zhang, H.C.; Liu, R.A. new helvolic acid derivative from an endophytic Fusarium sp. of Ficus carica. Nat. Prod. Res. 2016, 30, 2407–2412. [Google Scholar] [CrossRef] [PubMed]
- Hussain, H.; Drogies, K.H.; Al-Harrasi, A.; Hassan, Z.; Shah, A.; Ran, U.A.; Green, I.R.; Draeger, S.; Schulz, B.; Krohn, K. Antimicrobial constituents from endophytic fungus Fusarium sp. Asian Pac. J. Trop. Dis. 2015, 5, 186–189. [Google Scholar] [CrossRef]
- Mondol, M.A.M.; Farthouse, J.; Islam, M.T.; Schueffler, A.; Laatsch, H. Metabolites from the endophytic fungus Curvularia sp. M12 act as motility inhibitors against Phytophthora capsici Zoospores. J. Nat. Prod. 2017, 80, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Silva, G.H.; Zeraik, M.L.; de Oliveira, C.M.; Teles, H.L.; Trevisan, H.C.; Pfenning, L.H.; Nicolli, C.P.; Young, M.C.M.; Mascarenhas, Y.P.; Abreu, L.M.; et al. Lactone derivatives produced by a Phaeoacremonium sp., an endophytic fungus from Senna spectabilis. J. Nat. Prod. 2017, 80, 1674–1678. [Google Scholar] [CrossRef] [PubMed]
- Taware, R.; Abnave, P.; Patil, D.; Rajamohananan, P.R.; Raja, R.; Soundararajan, G.; Kundu, G.C.; Ahmad, A. Isolation, purification and characterization of Trichothecinol-A produced by endophytic fungus Trichothecium sp. and its antifungal, anticancer and antimetastatic activities. Sustain. Chem. Process. 2014, 2, 8. [Google Scholar] [CrossRef]
- Taware, R.; Abnave, P.; Patil, D.; Rajamohananan, P.R.; Raja, R.; Soundararajan, G.; Kundu, G.C.; Kharat, M.K.D.; Pai, K.; Ahmad, A. Trichothecin from endophytic fungus Trichothecium sp. and its anticancer effect on murine melanoma and breast cancer cell lines. Curr. Biochem. Eng. 2015, 2, 73–80. [Google Scholar] [CrossRef]
- Morandini, L.M.B.; Neto, A.T.; Pedroso, M.; Antoniolli, Z.I.; Burrow, R.A.; Bortoluzzi, A.J.; Mostardeiro, M.A.; da Silva, U.F.; Dalcol, I.I.; Morel, A.F. Lanostane-type triterpenes from the fungal endophyte Scleroderma UFSMSc1 (Persoon) Fries. Bioorg. Med. Chem. Lett. 2016, 26, 1173–1176. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.L.; Wang, G.P.; Mao, L.J.; Komon-Zelazowska, M.; Yuan, Z.L.; Lin, F.C.; Druzhinina, I.S.; Kubicek, C.P. Muscodor fengyangensis sp. nov. from southeast China: Morphology, physiology and production of volatile compounds. Fungal Biol. 2010, 114, 797–808. [Google Scholar] [CrossRef] [PubMed]
- Woropong, J.; Strobel, G.A.; Ford, E.J.; Li, J.Y.; Baird, G.; Hess, W.M. Muscodor albusanam. nov., an endophyte from Cinnamomum zeylanicum. Mycotaxon 2001, 79, 67–79. [Google Scholar]
- Strobel, G.A. Muscodor species-endophytes with biological promise. Phytochem. Rev. 2011, 10, 165–172. [Google Scholar] [CrossRef]
- DiFranscesco, A.; Ugolini, L.; Lazzeri, L.; Mari, M. Production of volatile organic compounds by Aureobasidium pullulans as a potential mechanism against post-harvest fruit pathogens. Biol. Control 2015, 81, 8–14. [Google Scholar] [CrossRef]
- Oro, L.; Feliziani, E.; Ciani, M.; Romanazzi, G.; Comitini, F. Volatile organic compounds of Wickerhamomyces anomalus, Metschinikowia pulcherrima and Saccharomyces cerevisiae inhibit growth of decay causing fungi and post-harvest disease of strawberries. J. Food Microbiol. 2018, 265, 18–22. [Google Scholar] [CrossRef] [PubMed]
- Pimenta, R.S.; da Silva, J.F.M.; Buyer, J.S.; Janiesiewicz, W.J. Endophytic fungi from Plums (Prunis domestica) and their antifungal activity against Monilinia fruticola. J. Food Prot. 2012, 75, 1883–1889. [Google Scholar] [CrossRef] [PubMed]
- Medina-Romaro, Y.M.; Roque-Flores, G.; Macias-Rubalcava, M.L. Volatile organic compounds from endophytic fungi as innovative post-harvest control of Fusarium oxysporum in cherry tomato fruits. Appl. Microbiol. Biotechnol. 2017, 101, 8209–8222. [Google Scholar] [CrossRef] [PubMed]
- Strobel, G.A.; Dirkse, E.; Sears, J.; Markworth, C. Volatile antimicrobials from Muscodor albus, a novel endophytic fungus. Microbiology 2001, 147, 2943–2950. [Google Scholar] [CrossRef] [PubMed]
- Daisy, B.H.; Strobel, G.A.; Castillo, U.; Ezra, D.; Sears, J.; Weaver, D.K.; Runyon, J.B. Naphthalene, an insect repellent, is produced by Muscodor vitigenus, a novel endophytic fungus. Microbiology 2002, 148, 3737–3741. [Google Scholar] [CrossRef] [PubMed]
- Grimme, E.; Zidack, N.K.; Sikora, R.A.; Strobel, G.A.; Jacobsen, B.J. Comparison of Muscodor albus volatiles with abiorational mixture for control of seedling diseases of sugar beet and root knot nematode on tomato. Plant Dis. 2007, 91, 220–225. [Google Scholar] [CrossRef]
- Strobel, G.A.; Knighton, B.; Kluck, K.; Ren, Y.; Livinghouse, T.; Griffin, M.; Spakowicz, D.; Sears, J. The production of myco-diesel hydrocarbons and their derivatives by the endophytic fungus Gliocladiumroseum (NRRL 50072). Microbiology 2008, 154, 3319–3328. [Google Scholar] [CrossRef] [PubMed]
- Worapong, J.; Strobel, G.; Daisy, B.; Castillo, U.F.; Baird, G.; Hess, M.W. Muscodor roseousanam. Sp. Nov. and endophyte from Grevillea pteridifolia. Mycotaxon 2002, 81, 463–475. [Google Scholar]
- González, M.C.; Anaya, A.L.; Glenn, A.E.; Macías-Rubalcava, M.L.; Hernández-Bautista, B.E.; Hanlin, R.T. Muscodor yucatanensis, a new endophytic ascomycete from Mexican chakah, Bursera simaruba. Mycotaxon 2009, 110, 363–372. [Google Scholar] [CrossRef]
- Mitchell, A.M.; Strobel, G.A.; Moore, E.; Robison, R.; Sears, J. Volatile antimicrobials from Muscodor crispans, a novel endophytic fungus. Microbiology 2010, 156, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Kudalkar, P.; Strobel, G.; Riyaz-Ul-Hassan, S.; Geary, B.; Sears, J. Muscodor sutura, a novel endophytic fungus with volatile antibiotic activities. Mycoscience 2012, 53, 319–325. [Google Scholar] [CrossRef]
- Suwannarach, N.; Kumla, J.; Bussaban, B.; Hyde, K.D.; Matsui, K.; Lumyong, S. Molecular and morphological evidence support four new species in the genus Muscodor from northern Thailand. Ann. Microbiol. 2013, 63, 1341–1351. [Google Scholar] [CrossRef]
- Meshram, V.; Kapoor, N.; Saxena, S. Muscodor kashayum sp. nov.—A new volatile anti-microbial producing endophytic fungus. Mycology 2013, 4, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Saxena, S.; Meshram, V.; Kapoor, N. Muscodor darjeelingensis, a new endophytic fungus of Cinnamomum camphora collected from north eastern Himalayas. Sydowia 2014, 66, 55–67. [Google Scholar]
- Meshram, V.; Saxena, S.; Kapoor, N. Muscodor strobelii, a new endophytic species from South India. Mycotaxon 2014, 128, 93–104. [Google Scholar] [CrossRef]
- Saxena, S.; Meshram, V.; Kapoor, N. Muscodor tigerii sp. nov. Volatile antibiotic producing endophytic fungus from the North eastern Himalayas. Ann. Microbiol. 2015, 65, 47–57. [Google Scholar] [CrossRef]
- Siri-udom, S.; Suwannarach, N.; Lumyong, S. Existence of Muscodor vitigenus, M. equiseti and M. heveae sp. nov.in leaves of the rubber tree (Hevea brasiliensis Müll. Arg.), and their biocontrol potential. Ann. Microbiol. 2015, 66, 437–448. [Google Scholar] [CrossRef]
- Meshram, V.; Gupta, M.; Saxena, S. Muscodor ghoomensis and Muscodor indica: New endophytic species based on morphological features, molecular and volatile organic analysis from northeast India. Sydowia 2016, 67, 133–146. [Google Scholar]
- Meshram, V.; Kapoor, N.; Chopra, G.; Saxena, S. Muscodor camphora, a new endophytic species from Cinnamomum camphora. Mycosphere 2017, 8, 568–582. [Google Scholar] [CrossRef]
- Scherlach, K.; Hertweck, C. Triggering cryptic natural product biosynthesis in microorganisms. Org. Biomol. Chem. 2009, 7, 1753–1760. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.B.; Henrikson, J.C.; Hoover, A.R.; Lee, A.E.; Cichewicz, R.H. Epigenetic remodeling of the fungal secondary metabolome. Org. Biomol. Chem. 2008, 6, 1895–1897. [Google Scholar] [CrossRef] [PubMed]
- Cichewicz, R.H. Epigenome manipulation as a pathway to new natural product scaffolds and their congeners. Nat. Prod. Rep. 2010, 27, 11–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fox, E.M.; Howlett, B.J. Secondary metabolism, regulation and role in fungal biology. Curr. Opin. Microbiol. 2008, 11, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Fisch, K.M.; Gillaspy, A.F.; Gipson, M.; Henrikson, J.C.; Hoover, A.R.; Jackson, L.; Najar, F.Z.; Wägele, H.; Cichewicz, R.H. Chemical induction of silent pathway transcription in Aspergillus niger. J. Ind. Microbiol. Biotechnol. 2009, 36, 1199–1213. [Google Scholar] [CrossRef] [PubMed]
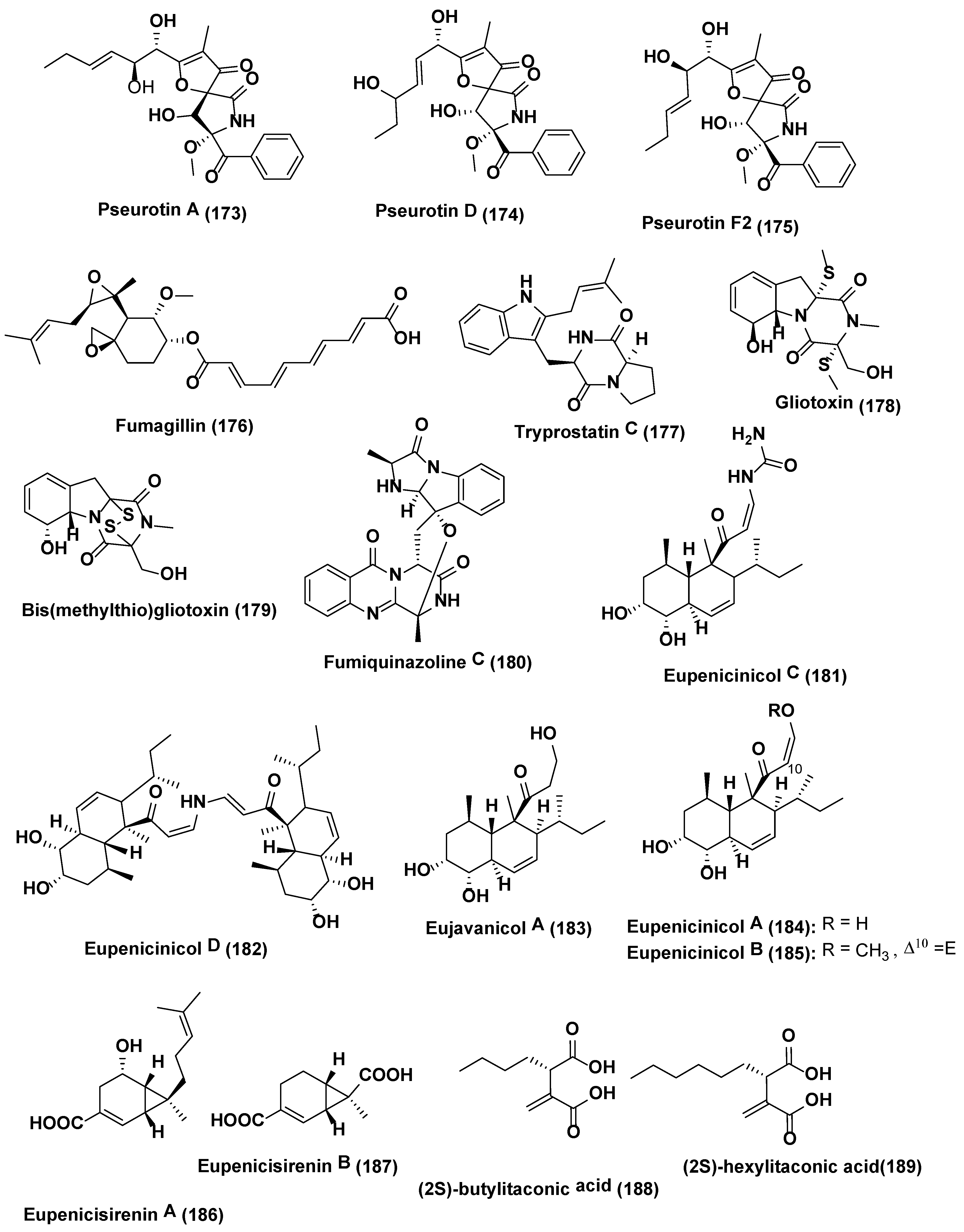
- Magotra, A.; Kumar, M.; Kushwaha, M.; Awasthi, P.; Raina, C.; Gupta, A.P.; Shah, B.A.; Gandhi, S.G.; Chaubey, A. Epigenetic modifier induced enhancement of fumiquinazoline C production in Aspergillus fumigatus (GA-L7): An endophytic fungus from Grewia asiatica L. AMB Express 2017, 7, 43. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Kusari, S.; Golz, C.; Laatsch, H.; Strohmann, C.; Spiteller, M. Epigenetic modulation of endophytic Eupenicillium sp. LG41 by a histone deacetylase inhibitor for production of decalin-containing compounds. J. Nat. Prod. 2017, 80, 983–988. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Kusari, S.; Lamshöft, M.; Schüffler, A.; Laatsch, H.; Spiteller, M. Antibacterial secondary metabolites from an endophytic fungus, Eupenicillium sp. LG41. J. Nat. Prod. 2014, 77, 2335–2341. [Google Scholar] [CrossRef] [PubMed]
- Vasanthakumari, M.M.; Jadhav, S.S.; Sachin, N.; Vinod, G.; Singh, S.; Manjunatha, B.L.; Kumara, P.M.; Ravikanth, G.; Nataraja, K.N.; Uma Shaanker, R. Restoration of camptothecine production in attenuated endophytic fungus on re-inoculation into host plant and treatment with DNA methyltransferase inhibitor. World. J. Microbiol. Biotechnol. 2015, 31, 1629–1639. [Google Scholar] [CrossRef] [PubMed]
- Gubiani, J.R.; Wijeratne, E.M.K.; Shi, T.; Araujo, A.R.; Arnold, A.E.; Chapman, E.; Gunatilaka, A.A.L. An epigenetic modifier induces production of (10′ S)-verruculide B, an inhibitor of protein tyrosine phosphatases by Phoma sp. nov. LG0217, a fungal endophyte of Parkinsonia microphylla. Bioorg. Med. Chem. 2017, 25, 1860–1866. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.; Zhu, G.; Hao, J.; Wang, Y.; Zhu, W. Chemical-epigenetic Method to Enhance the Chemodiversity of the Marine Algicolous Fungus, Aspergillus terreus OUCMDZ-2739. Tetrahedron 2018, 74, 83–87. [Google Scholar] [CrossRef]
- Akone, S.H.; Mandi, A.; Kurtan, T.; Hartmann, R.; Lin, W.; Daletos, G.; Proksch, P. Inducing secondary metabolite production by the endophytic fungus Chaetomium sp. through fungal-bacterial co-culture and epigenetic modification. Tetrahedron 2016, 72, 6340–6347. [Google Scholar] [CrossRef]
- Asai, T.; Otsuki, S.; Sakurai, H.; Yamashita, K.; Ozeki, T.; Oshima, Y. Benzophenones from an endophytic fungus, Graphiopsis chlorocephala, from Cultivated in the Presence of an NAD+-Dependent HDAC Inhibitor. Org. Lett. 2013, 15, 2058–2061. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Menendez, V.; Perez-Bonilla, M.; Perez-Victoria, I.; Martin, J.; Munoz, F.; Reyes, F.; Tormo, J.R.; Genilloud, O. Multicomponent analysis of the differential induction of secondary metabolite profiles in fungal endophytes. Molecules 2016, 21, 234. [Google Scholar] [CrossRef] [PubMed]
- Rico-Gray, V. Interspecific interaction. Encycl. Life Sci. 2001. [Google Scholar] [CrossRef]
- Sandland, G.J.; Rodgers, J.K.; Minchella, D.J. Interspecific antagonism and virulence in hostsex-posed to two parasite species. J. Invertebr. Pathol. 2007, 96, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Marmann, A.; Aly, A.H.; Lin, W.; Wang, B.; Proksch, P. Co-Cultivation—A powerful emerging tool for enhancing the chemical diversity of microorganisms. Mar. Drugs 2014, 12, 1043–1065. [Google Scholar] [CrossRef] [PubMed]
- Brakhage, A.A.; Schroeckh, V. Fungal secondary metabolites–strategies to activate silent gene clusters. Fungal Genet. Biol. 2011, 48, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Ola, A.R.; Thomy, D.; Lai, D.; Brötz-Oesterhelt, H.; Proksch, P. Inducing secondary metabolite production by the endophytic fungus Fusarium tricinctum through co-culture with Bacillus subtilis. J. Nat. Prod. 2013, 76, 2094–2099. [Google Scholar] [CrossRef] [PubMed]
- Kusari, S.; Singh, S.; Jayabaskaran, C. Rethinking production of Taxol® (paclitaxel) using endophyte biotechnology. Trends Biotechnol. 2014, 32, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Ebrahim, W.; El-Neketi, M.; Lewald, L.I.; Orfali, R.S.; Lin, W.; Rehberg, N.; Kalscheuer, R.; Daletos, G.; Proksch, P. Metabolites from the Fungal Endophyte Aspergillus austroafricanus in Axenic Culture and in Fungal-Bacterial Mixed Cultures. J. Nat. Prod. 2016, 79, 914–922. [Google Scholar] [CrossRef] [PubMed]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
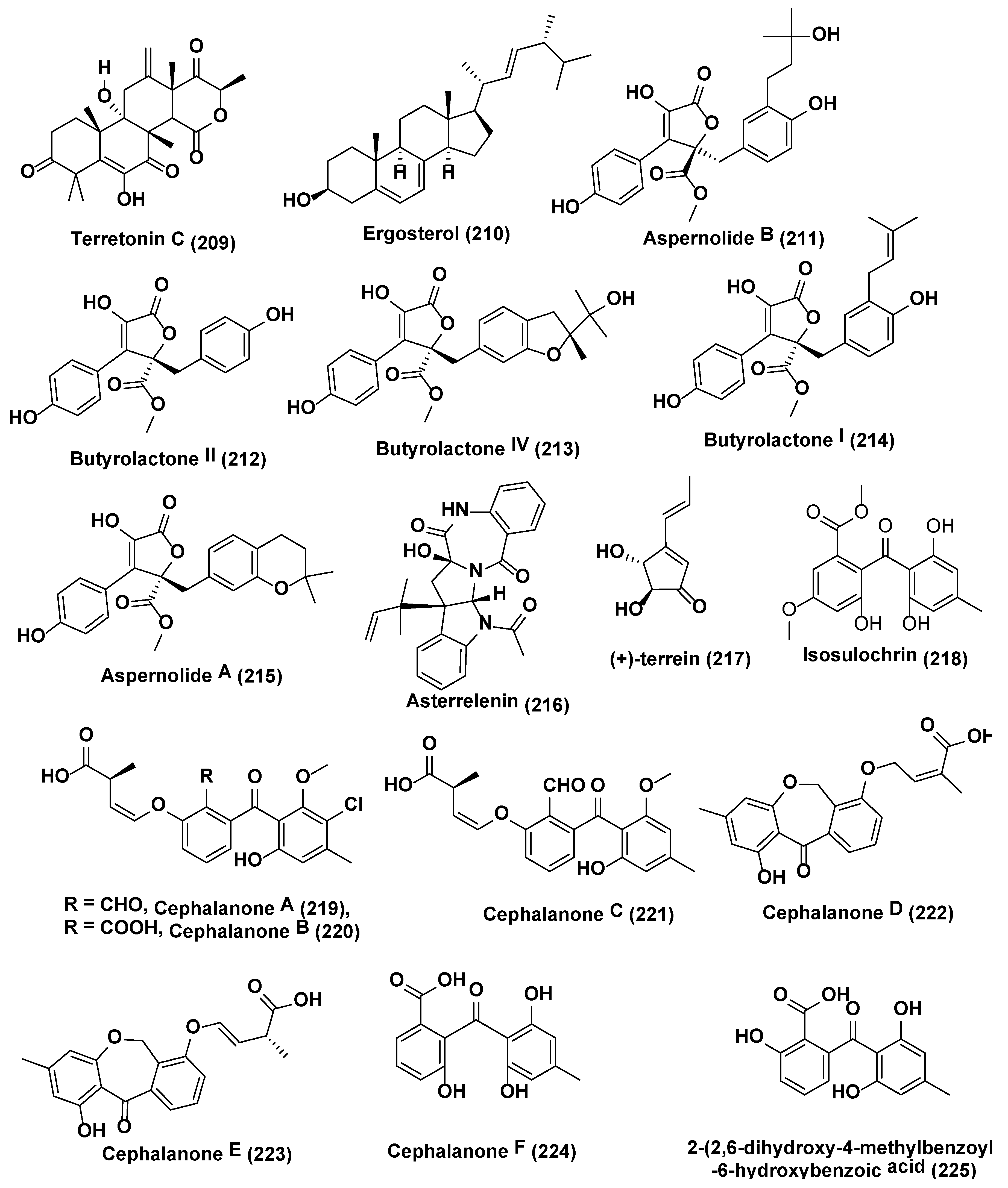{kind=link}
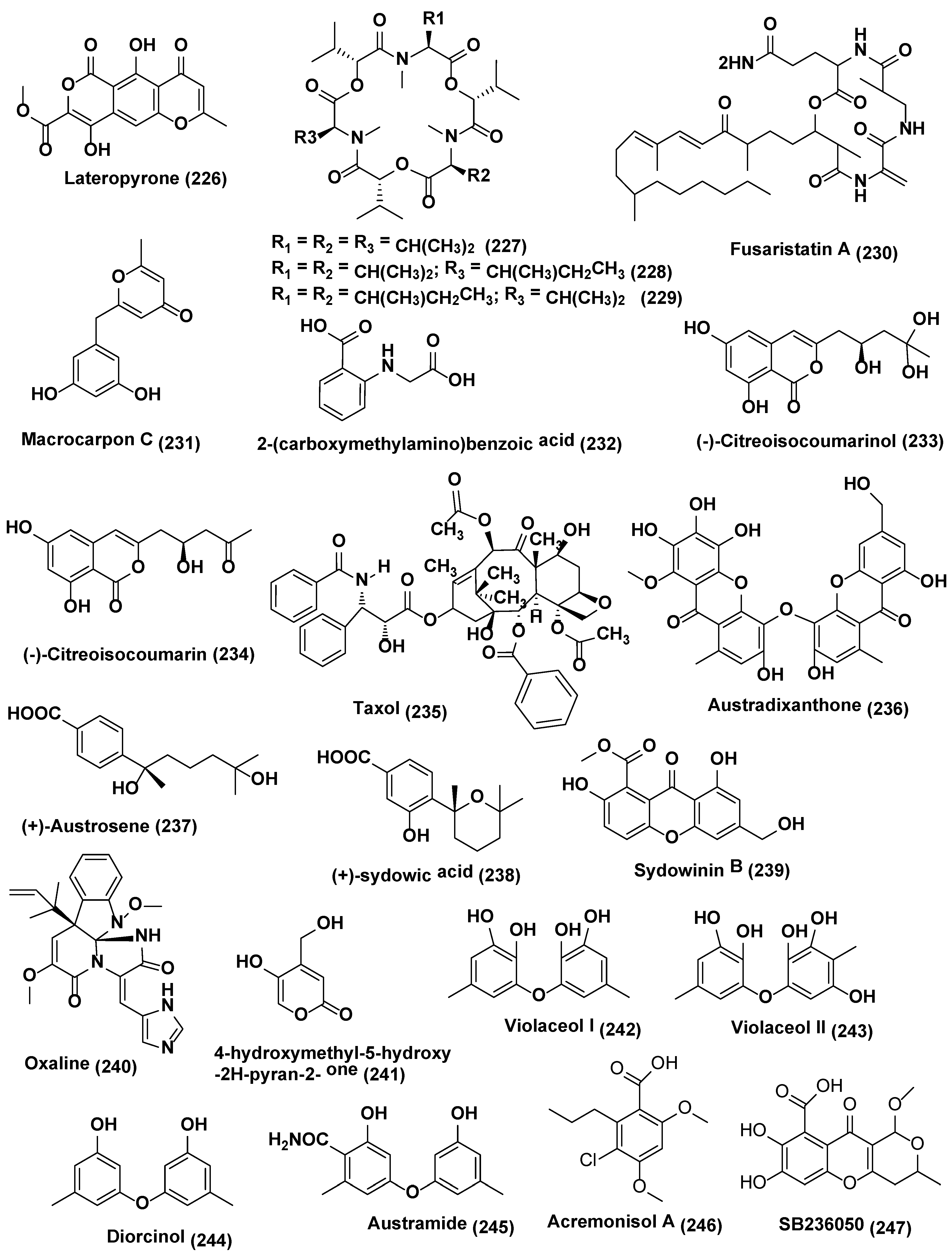{kind=link}
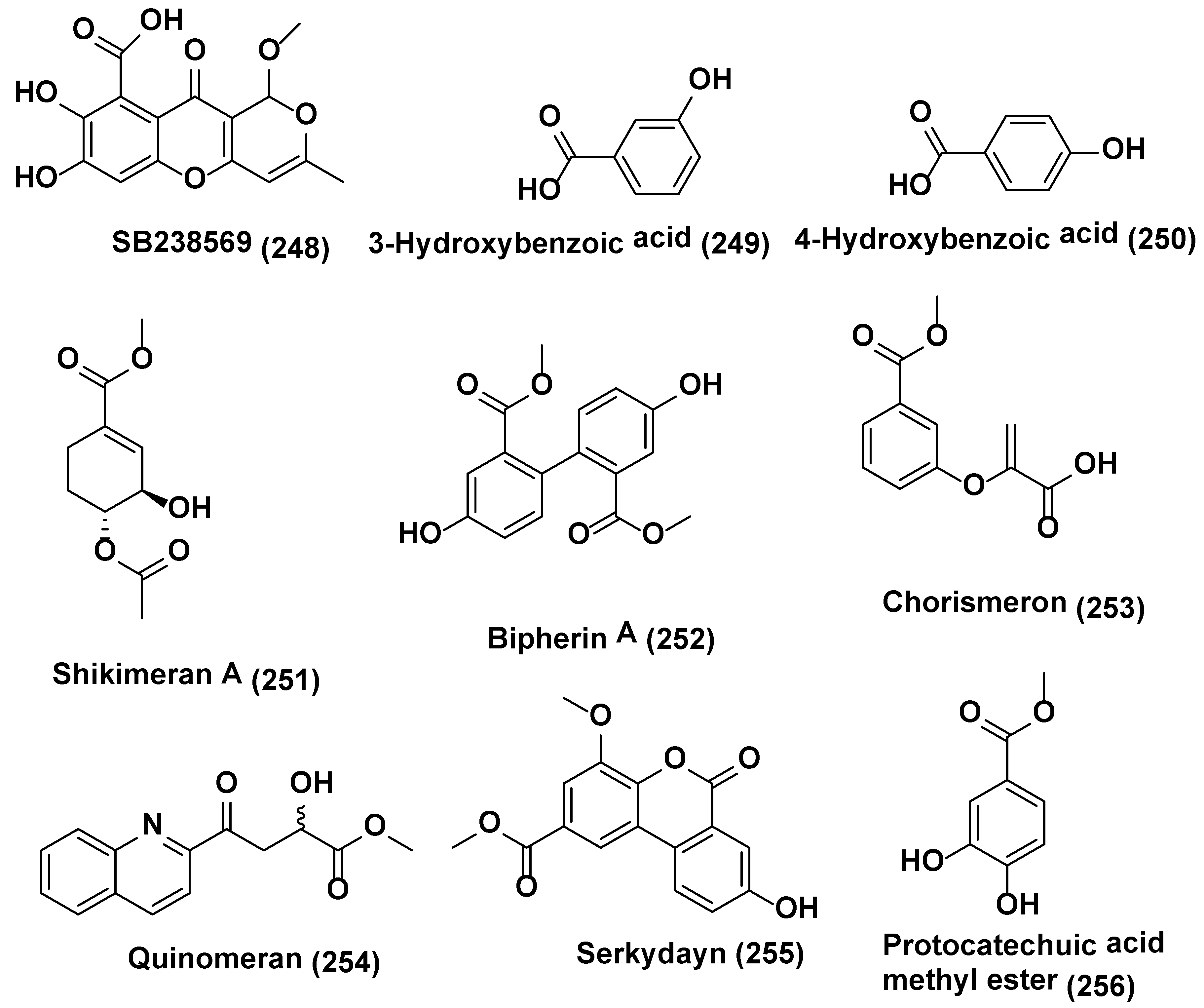{kind=link}
| Sr. No. | Fungus | Plantsource | Compounds Isolated | Biologicalactivity * | Refs. |
|---|---|---|---|---|---|
| Comounds Produced by Coelomycetes | |||||
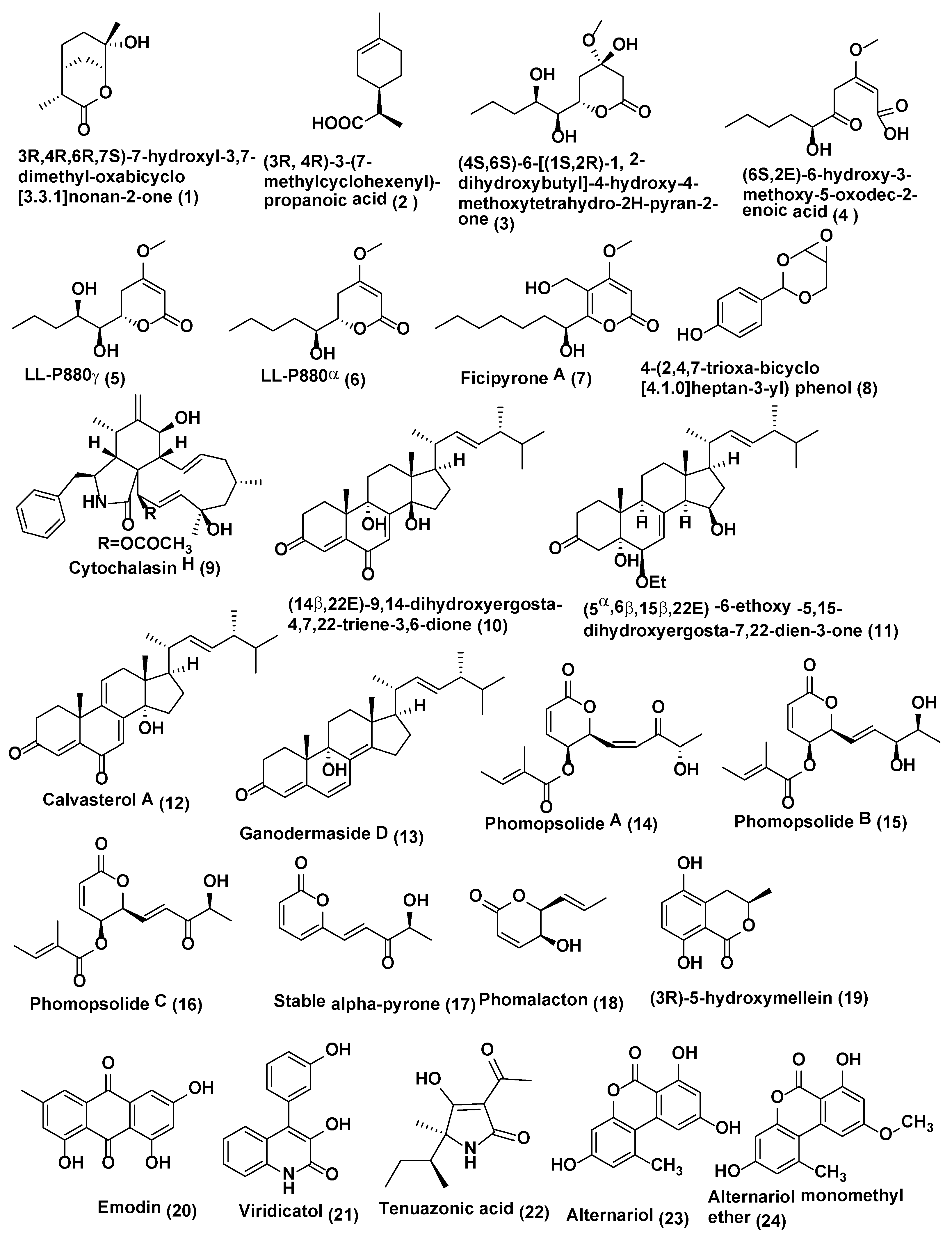
| 1 | Pestalotiopsis foedan | Bruguiera sexangula Hainan, China | (3R,4R,6R,7S)-7-hydroxyl-3,7-dimethyl-oxabicyclo[3.3.1]nonan-2-one (1), (3R,4R)-3-(7-methylcyclohexenyl)-propanoic acid (2) | Compound 1 B. cinerea and P. nicotianae (MIC 3.1 and 6.3 µg/mL), ketoconazole (MIC 3.1 µg/mL each) Compound 2 C. albicans MIC 50 µg/mL) ketoconazole (MIC 6.3 µg/mL) | [15] |
| 2 | Pestalotiopsis sp. DO14 | Dendrobium officinale, Yandang Mountain, Zhejiang Province, China. | (4S,6S)-6-[(1S,2R)-1,2-dihydroxybutyl]-4-hydroxy-4-methoxytetrahydro-2H-pyran-2-one (3) and (6S,2E)-6-hydroxy-3-methoxy-5-oxodec-2-enoic acid (4), LL-P880γ (5), LL-P880α (6) | Compounds 3–6 active against C. albicans, C. neoformans, T. rubrum, and A. fumigates (MIC ≤ 50 µg/mL) Compounds 3–4 active against C. albicans, C. neoformans, T. rubrum, and A. fumigatus (MIC, ≤ 25 µg/mL) | [16] |
| 3 | Pestalotiopsis fici | Camellia sinensis Hangzhou, China. | Ficipyrone A (7) | Compound 7 active against G. zeae (IC50 15.9 µM), ketoconazole (IC50 6.02 µM) | [17] |
| 4 | Pestalotiopsis mangiferae | Mangifera indica Maduravoyal, Tamil Nadu Province, India. | 4-(2,4,7-trioxa-bicyclo[4.1.0]heptan-3-yl) phenol (8) | Compound 8 active against C. albicans (MIC, 0.039 µg/mL), Nystatin (MIC 10.0 µg/mL) | [18] |
| 5 | Phomopsis sp. | Senna spectabilis São Paulo, Brazil | Cytochalasin H (9) | Compound 9 active against C. cladosporioides and C. sphaerosphermum (MIC 10.0 and 25.0 µg, respectively), nystatin (MIC = 1.0 µg) | [19] |
| 6 | Phomopsis sp. | Aconitum carmichaeli, Huize County, Yunnan Province, China. | (14β,22E)-9,14-dihydroxyergosta-4,7,22-triene-3,6-dione (10), (5α,6β,15β,22E)-6-ethoxy-5,15-dihydroxyergosta-7,22-dien-3-one (11), calvasterols A (12), and ganodermaside D (13) | Compound 10 active against C. albicans, H. compactum, and A. niger, (MIC, 64, 64, and 128 µg/mL, respectively). Compound 11 active against C. albicans and F. avenaceum (MIC = 128 µg/mL). Compounds 12 and 13 active against F. avenaceum. (MIC, 64 µg/mL), Compound 12 activie against P. oryzae and T. gypseum (MIC 128 and 256 µg/mL) | [20] |
| 7 | Diaporthe maritima | Picea sp., Acadian forest of Eastern Canada. | Phomopsolide A (14), B (15), and C (16), and a stable alpha-pyrone (17) | Compound 14 active against M. violaceum and S. cerevisiae at 25 µM, Compounds 15–17 demonstrated growth inhibition at 250 µM | [21] |
| 8 | Phoma sp. | Fucus serratus, | Phomalacton (18), (3R)-5-hydroxymellein (19) and emodin (20) | Compounds 18–20 active against M. violaceum with 5, 6 and 5mm zone of inhibition. | [23] |
| 9 | Phoma sp. WF4 | Eleusine coracana Arkell Field Station, Arkell, ON, Canada | Viridicatol (21), tenuazonic acid (22), alternariol (23), and alternariol monomethyl ether (24) | Compounds 21–24 caused dramatic breakage of F. graminearum hyphae in vitro | [24] |
| 10 | Rhizopycnis vagum Nitaf 22 | Nicotiana tabacum, China Agricultural University. Beijing 100193, China. | Rhizopycnin D (25) and TMC-264 (26) | Compounds 25–26 inhibited the spore germination of M. oryzae with IC50 values of 9.9 and 12.0 µg/mL, respectively | [25] |
| 11 | Microsphaeropsis sp. Seimatosporium sp. | Salsola oppositifolia, Playa del Ingles, Gomera, Spain | Microsphaerol (27) Seimatorone (28) | Compounds 27 and 28 active against M. violaceum with 9 and 5 mm zone of inhibition. In addition, there was some growth with in zone of inhibition | [26] |
| 12 | Colletotrichum gloeosporioides | Michelia champaca São Paulo State University, Araraquara, São Paulo, Brazil. | 2-phenylethyl 1H-indol-3-yl-acetate (29) | Compound 29 active against C. cladosporioides and C. sphaerospermum comparable to that of the positive control nystatin | [27] |
| 13 | Colletotrichum sp. | Gomera (Spain). | Colletonoic acid (30) | Compound 30 active against M. violaceum with 7 mm zone of inhibition | [28] |
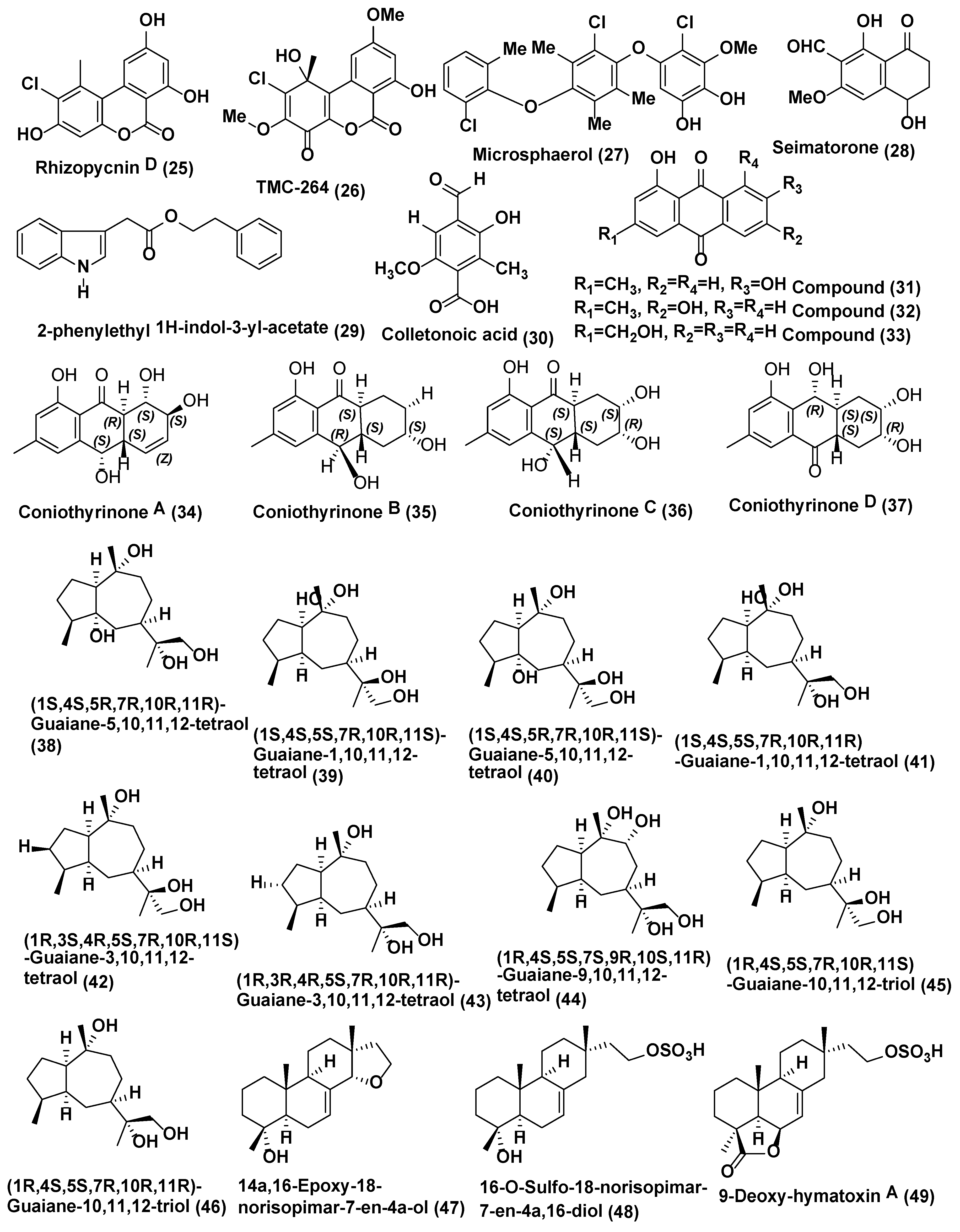
| 14 | Coniothyrium sp., | Salsola oppostifolia Gomera in the Canary Islands. | 1,7-dihydroxy3-methyl-9,10-anthraquinone (31), 1,6-dihydroxy-3 -methyl-9,10-anthraquinone (phomarin) (32), and 1-hydroxy-3-hydroxymethyl-9,10-anthraquinone (33) coniothyrinones A-D (34–37) | Compounds 31–37 active against M. violaceum with 7, 10, 8, 7.5, 6, 8 and 7.5 mm zone of inhibition. Compounds 32–34 active against M. violaceum (10 and 9 mm zone of inhibition) and B. cinerea (7.5 and 12.5 mm zone of inhibition) when tested under similar conditions | [29,30] |
| Comounds Produced by Acsomycetes | |||||
| 15 | Xylaria sp. YM 311647 | Azadirachta indica,Yuanjiang County, Yunnan Province, China, | (1S,4S,5R,7R,10R,11R)-Guaiane-5,10,11,12-tetraol (38) (1S,4S,5S,7R,10R,11S) -Guaiane-1,10,11,12-tetraol (39) (1S,4S,5R,7R,10R,11S)-Guaiane-5,10,11,12-tetraol (40) (1S,4S,5S,7R,10R,11R)-Guaiane-1,10,11,12-tetraol (41) (1R,3S,4R,5S,7R,10R,11S) -Guaiane-3,10,11,12-tetraol (42) (1R,3R,4R,5S,7R,10R,11R)-Guaiane-3, 10,11,12-tetraol (43) (1R,4S,5S,7S,9R,10S,11R)-Guaiane-9,10,11,12-tetraol (44) (1R,4S,5S,7R,10R,11S) -Guaiane-10,11,12-triol (45) (1R,4S,5S,7R,10R,11R)-Guaiane-10,11,12-triol (46), 14a,16-Epoxy-18-norisopimar-7-en-4a-ol (47),16-O-Sulfo-18-norisopimar-7-en-4a,16-diol (48), and 9-Deoxy-hymatoxin A (49) | Compounds 38–46 active against C. albicans and H. compactum (MIC in the range of 32 to 256 µg/mL), compounds 47–49 active against C. albicans, A. niger, P. oryzae, F. avenaceum, and H. compactum (MIC in the range of 16 to 256 µg/mL). Compound 49 exhibited the potent inhibitory activity against C. albicans and P. oryzae with MIC values of 16 µg/mL | [32] |
| 16 | Xylaria sp. YM 31164 | Azadirachta indica, Yuanjiang County, Yunnan Province, China | (1S,2S,4S,5S,7R,10R)-Guaiane-2,10,11,12-tetraol (50), (1S,2S,4R,5R,7R,10R)-Guaiane-2,4,10,11,12-pentaol (51), (1S,4R,5S,7R,10R)-Guaiane-4,5,10,11,12-pentaol (52), (1R,4S,5R,7R,10R)-Guaiane-1,5,10,11,12-pentaol (53), (1R,4R,5R,7R,10R)-11-Methoxyguaiane-4,10,12-triol (54), | Compounds 50–54 active against P. oryzae and H. compactum (MIC in the range of 32–256 µg/mL). Compound 53 active against P. oryzae (MIC 32 µg/mL). Compounds 52 and 53 active against H. compactum with (MIC, 64 µg/mL), Compound 53 and 54 active against C. albicans (MIC 32 µg/mL). Compound 52 active against C. albicans, A. niger, and H. compactum (MIC, 64 µg/mL). | [33] |
| 17 | X. feejeensis | Croton lechleri. | Xyolide (55), | Compound 55 active against P. ultimum (MIC 425 µM) | [34] |
| 18 | Xylaria sp. XC-16 | Toona sinensis Yangling, Shaanxi Province, China | Cytochalasin Z28 (56) | Compound 56 active against G. saubinetti (MIC of 12.5 µM), Hymexazol (MIC = of 25 µM) | Zhang et al. [35] |
| 19 | Xylaria sp. strain F0010, Xylaria sp. PSU-G12X. cubensis, 13 strains of Xylaria sp. | Abies holophylla; Garcinia hombroniana ; Asimina triloba; Pinus strobus; Vaccinium angustifolium, New Brunswick and Nova Scotia, Canada | Griseofulvin (57) | Griseofulvin (57) Inhibits A. mali, B. cinerea, Colletotrichum gloeosporioides, Corticium sasaki, F. oxysporum and M. grisea in in vitro (IC50 values of 18.0, 5.0, 1.7, 11.0, 30.0, and 1.7 µg/mL, respectively, Compound 57 active against M. grisea, C. sasaki, B. cinerea, P. recondite and B .graminis f. sp. hordei in in vivo, with % of fungal control of 95, 100, 60, 90 and 90, respectively, at 150 µg/mL. | [36,37,38] |
| 20 | Xylaria sp. | Dechlorogriseofulvin (58) | Compound 58 showed weak antifungal activity, with an IC50 value, 200 µg/mL against M. grisea, C. sasaki, B. cinerea, P. recondite and B .graminis f. sp. hordei in in vivo, | [36,38] | |
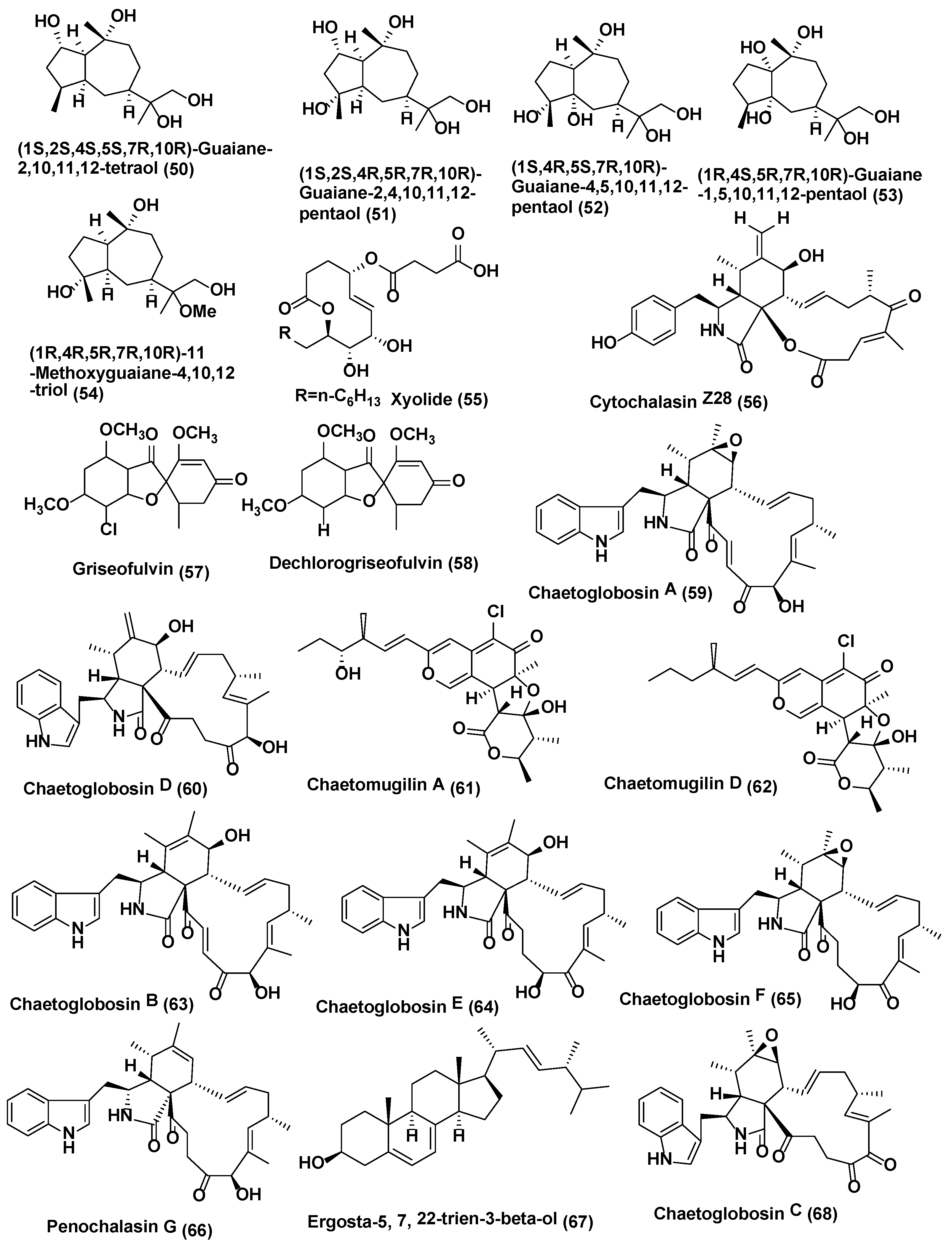
| 21 | Chaetomium globosum CDW7 | Ginkgo biloba China | Chaetoglobosin A (59) and D (60) | Compounds 59–60 active against S. sclerotiorum with IC50 values of 0.35 and 0.62 µg/mL, respectively, carbendazim (0.17 µg/mL) | [41] |
| 22 | Chaetomium globosum | Seeds of Panax notoginseng collected at the Wenshan, Yunnan, China | Chaetoglobosin A (59), Chaetomugilin A (61), Chaetomugilin D (62), Chaetoglobosin B (63), Chaetoglobosin E (64), Chaetoglobosin F (65) and Penochalasin G (66) | Compounds 59 and 61–66 active against P. herbarum (MIC in the range of 16–128 µg/mL) and, E. nigrum (MIC in the range of <1–16 µg/mL). | [42] |
| 23 | Chaetomium cupreum ZJWCF079 | Macleaya cordata. | Ergosta-5, 7, 22-trien-3-beta-ol (67) | Compound 67 against S. sclerotiorum and B. cinerea with EC50 values of 125 µg/mL and 190 µg/mL respectively, | [43] |
| 24 | Chaetomium globosum No.04 | Barks of Ginkgo biloba, Linyi, Shandong Province, China. | Chaetoglobosin A (59), D (60), E (64), C (68), G (69), R (70) | Compounds 59–60, 64, and 68–70 active against R. stolonifer and C. diplodiella at a concentration of 20 µg/disk | [44] |
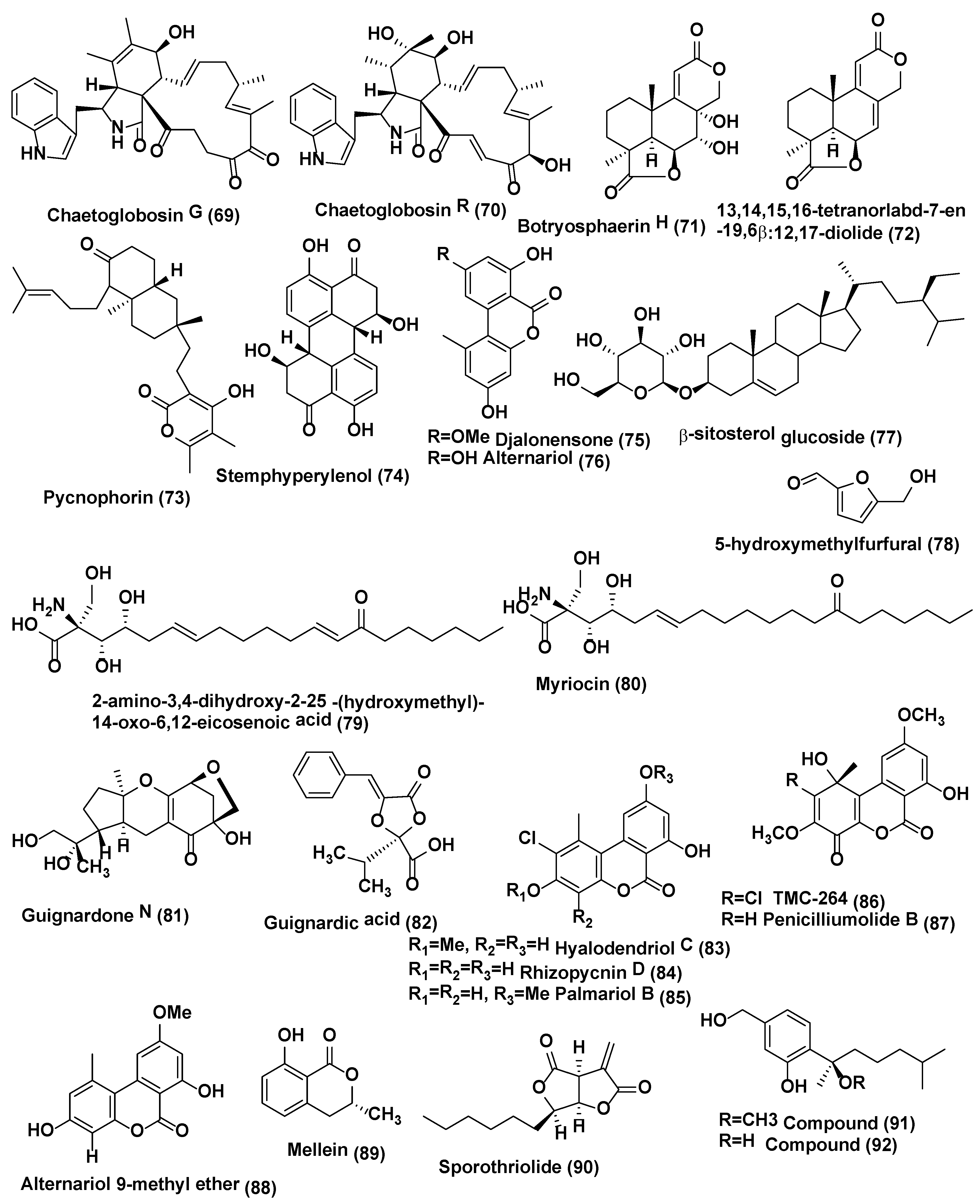
| 25 | Botryosphaeria sp. P483 | Huperzia serrata, Xichou County, Yunnan Province, China | Botryosphaerin H (71) 13,14,15,16-tetranorlabd-7-en-19,6β:12,17-diolide (72) | At 100 µg/disk, compound 71 showed zone of inhibition of 9, 7, 7, 8, and 8 mm, against G. graminis, F. solani, P. oryzae, F. moniliforme, and F. oxysporum; compound 72 showed zone of inhibition of 12, 10, 10, 11, and 13 mm against G. graminis, F. solani, P. oryzae, F. moniliforme, and F. oxysporum; carbendazim (50 µg/disk) showed the zone of inhibition of 14, 18, 15, 17, 15 mm against G. graminis F. solani P. oryzae F. moniliforme F. oxysporum, respectively | [45] |
| 26 | Botryosphaeriadothidea KJ-1, | Melia azedarach Yangling, Shaanxi Province, China. | Pycnophorin (73), stemphyperylenol (74), chaetoglobosin C (68), djalonensone (75), alternariol (76), β-sitosterol glucoside (77), 5-hydroxymethylfurfural (78) | Compound 74 active against A. solani (MICs of 1.57 µM) Compounds 68, 73, and 75–78 active against A. solani (MICs of 6.25−25 µM) | [46] |
| 27 | Mycosphaerella sp. | Eugenia bimarginata DC. Brazil (savannah). | 2-amino-3,4-dihydroxy-2-25-(hydroxymethyl)-14-oxo-6,12-eicosenoic acid (79), myriocin (80) | Compounds 79 active against several isolates of C. neoformans and C. gattii, with MIC values ranging from 1.3 to 2.50 µg/mL and 0.5 µg/mL, for compound 80 | [47] |
| Compounds 79 active against several isolates of C. neoformans and C. gattii, with MIC values ranging from 0.49 to 7.82 µM and 0.48–1.95 µM for compound 80. Compounds 79 and 80 cause deformities in cell shape, depressions on the surface, and withered cells. | [48] | ||||
| 28 | Guignardia sp., | Euphorbia sieboldiana collected from the campus of China Pharmaceutical University, Nanjing, Jiangsu, China | Guignardone N (81), guignardic acid (82) | At 6.3 µg/mL combined with 0.031 µg/mL of fluconazole, compounds 81 and 82 were found to have prominent inhibition on the growth of C. albicans with FIC index values of 0.23 and 0.19, respectively. Combined with fluconazole, both of them (40 µg/mL for (81) and 20 µg/mL for (82) could also inhibit C. albicans biofilms and reverse the tolerance of C. albicans biofilms to fluconazole | [49] |
| 29 | Hyalodendriella sp. Ponipodef 12 | “Neva” hybrid of Populus deltoides Marsh × P. nigra L., Longhua in Hebei Province of China. | hyalodendriol C (83), rhizopycnin D (84), palmariol B (85), TMC-264 (86), penicilliumolide B (87) and alternariol 9-methyl ether (88) | Compound 88 exhibited spore germination of M. oryzae with IC50 value of 11.6 µg/mL, positive control, carbendazim (IC50 6.9 µg/mL) Compounds 84–88 displayed antifungal effects against the spore germination of M. oryzae | [50,51,52] |
| 30 | Pezicula sp. | Forsythia viridissima, collected from Zhejiang Province, Southeast China | Mellein (89) | Compound 89 active against B. cinerea, P. ultimum, F. oxysporium f. sp. cucumerinum, C. orbiculare, V. dahliae, P. oryzae, P. diospyri, S. sclerotiorum and F. fulva, especially B. cinerea and F. fulva with EC50 values below 50 µg/mL | [53] |
| 31 | Nodulisporium sp. A21 | Leaves of Ginkgo biloba. Nanjing in Jiangsu Province, China | Sporothriolide, (90) | The EC50 of compound 90 against R.solani was 3.04 µg/mL (11.6 µM), while the EC50 of carbendazim was 1.84 µg/mL (9.6 µM). | [54] |
| 32 | Lopherdermium nitens DAOM 250027 | Pinus strobus Sussex, NB, Canada | Six phenolic bisabolane-type sesquiterpenoids (91–96), pyrenophorin (97) | Compound 97 significantly reduced the growth of M. violaceum and S. cerevisiae at 5 µM whereas sesquiterpenoids 91–96 active at 50 µM to both species tested | [55] |
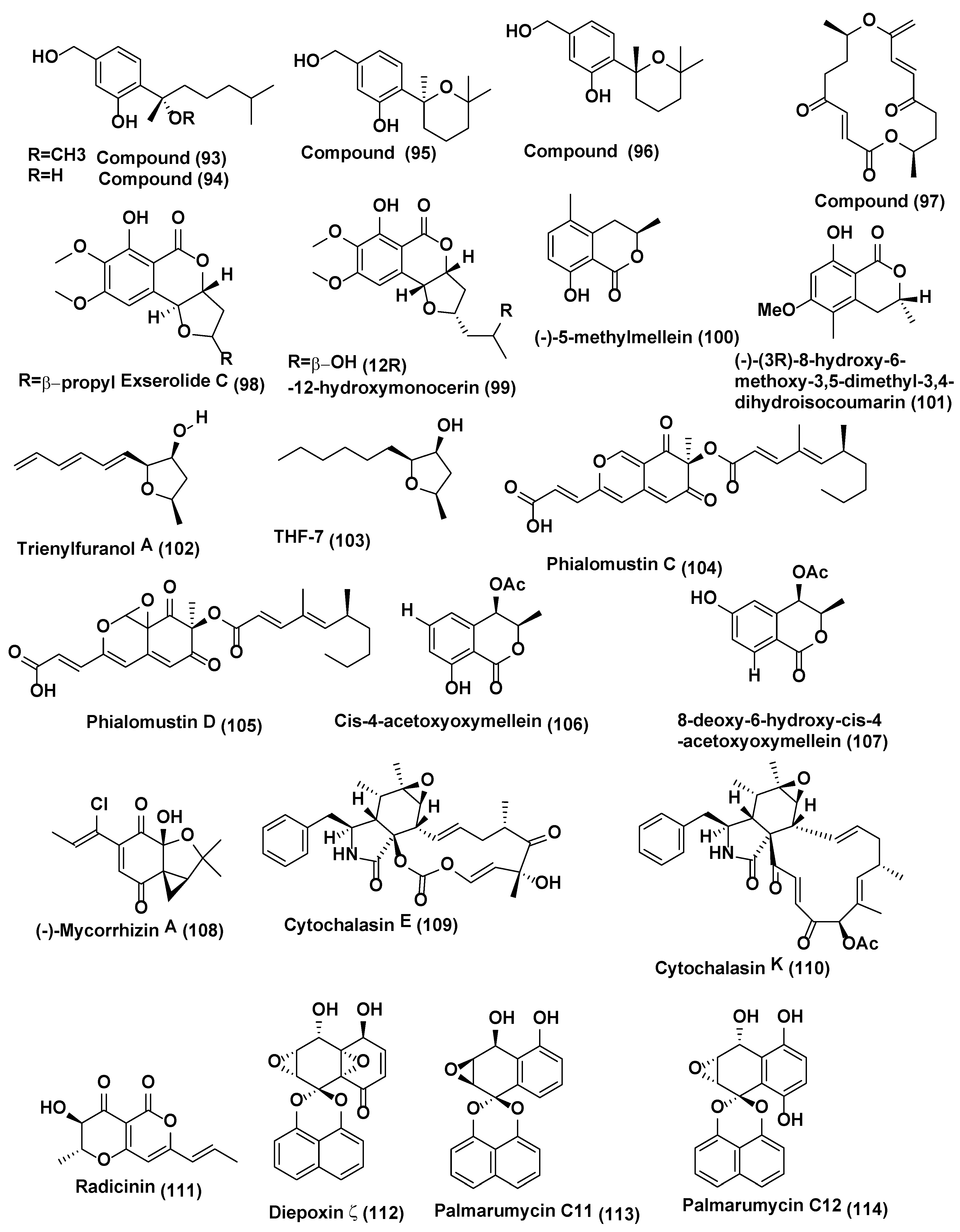
| 33 | Exserohilum sp. | Acer truncatum Beijing, China. | Exserolide C (98), (12R)-12-hydroxymonocerin (99) | Compounds 98 and 99 active against F. oxysporum, both showing a MIC value of 20 µg/mL, Amphotericin B (MIC, 0.63 µg/mL) | [56] |
| 34 | Biscogniauxia mediterranea EPU38CA | Echinacea purpurea Missouri, USA, | (−)-5-methylmellein (100) and (−)-(3R)-8-hydroxy-6-methoxy-3,5-dimethyl-3, 4-dihydroisocoumarin (101) | Compound 100 active against P. obscurans, P. viticola, and F.oxysporum, and caused growth stimulation of C. fragariae, C. acutatum, C. gloeosporioides, and B. cinerea. Compound 101 was found to be slightly more active in the microtiter environment than 5-methylmellein | [57] |
| 35 | Hypoxylon submonticulosum | Rubus idaeus collected from Jordan Station, ON, Canada. | Trienylfuranol A (102) Complete hydrogenation of (102) yielded THF 7 (103) | THF 7 (103) inhibited the growth of S. cervisiae (74 ± 4% inhibition) at a concentration of 250 µg/mL as compared with complete inhibition by nystatin at 10 µg/mL | [58] |
| 36 | Phialophoramustea | Crocus sativus. | Phialomustin C-D (104) (105) | Compounds 104–105 active against C. albicans (IC50, 14.3 and 73.6 µM) | [59] |
| 37 | unidentified Ascomycete, | Melilotus dentatus. | cis-4-acetoxyoxymellein (106) and 8-deoxy-6-hydroxy-cis-acetoxyoxymellein (107) | Compounds 106 and 107 displayed activities toward M. violaceum, B. cinerea, with 8 mm zone of inhibition for both fungi. | [60] |
| 38 | Plectophomella sp. | (−)-Mycorrhizin A (108) | Compound 108 active against U. violacea and E. repens. | [61] | |
| 39 | Physalospora sp. | Cytochalasin E (109) and K (110) | Compound 109–110 active against E. repens and M. microspora | [61] | |
| 40 | Crataegus monogyna. | Radicinin (111) | Radicinin (111) active against E. repens and M. microspora | [61] | |
| 41 | Berkleasmium sp., | Dioscorea zingiberensis. Hubei Province, China. | Diepoxin ζ (112), palmarumycin C11 (113), palmarumycin C12 (114), cladospirone B (115), palmarumycin C6 (116), 1,4,7β-trihydroxy-8-(spirodioxy-1′,8′-naphthyl)-7,8-dihydronaphthalene (117) and palmarumycin C8 (118) | Compounds 112–118 inhibited spore germination of M. oryzae (IC50 values in the range 9.1−124.5 µg/mL). Compound 118 showed the best inhibitory activity (IC50, 9.1 µg/mL) among the compounds tested. Carbendazim (IC50 6.3 µg/mL) | [62] |
| 42 | Bipolaris sp. MU34 | Gynura hispida Bangkok, Thailand. | Bipolamide B (119) | Bipolamide B (119) active against C. cladosporioides, C. cucumerinum, S. cerevisiae, A. niger and R. oryzae, with MIC values of 16, 32, 32, 64 and 64 µg/mL, respectively | [63] |
| 43 | Alternaria alternata Tche-153 | Terminalia chebula Rezt. Suanluang Rama IX Public Park, Bangkok, Thailand. | Altenusin (120) | Altenusin (120) in combination with each of three azole drugs, ketoconazole, fluconazole or itraconazole at their low sub-inhibitory concentrations exhibited potent synergistic activity against C. albicans with the FIC index range of 0.078 to 0.188 | [64] |
| 44 | Alternaria sp. UFMGCB 55, | Leaves of Trixis vauthieri DC (Asteraceae). | Altenusin (120) | The altenusin (120) exhibited strong activity against 11 strains P. brasiliensis with MIC values ranging between 1.9 and 31.2 µg/mL MIC values found for amphotericin B were between 0.031 and 0.12 µg/mL. Additionally, S. pombe cells treated with altenusin were more rounded in shape than untreated cellssuggeststhat altenusin could act through the inhibition of cell wall synthesis or assembly in P. brasiliensis and S. pombe | [65] |
| 45 | Cladosporium cladosporioides | Cladosporin (121), Isocladosporin (122) | At 30 µM compound 121 exhibited 92.7, 90.1, 95.4, and 79.9% growth inhibition against C. acutatum, C. fragariae, C. gloeosporioides and P. viticola respectively. Compound 122 showed 50.4, 60.2, and 83.0% growth inhibition at 30 µM against C. fragariae, C. gloeosporioides, and P. viticola, respectively | [66] | |
| 46 | Epicoccum sp. CAFTBO, | Theobroma cacao (Sterculiaceae) Mount Kala, near Yaoundé, Centre Province, Republic of Cameroon | Epicolactone (123), Epicoccolide A (124) and B (125) | Compounds 123–125 showed inhibitory effects on the mycelial growth of P. ultimum and A. cochlioides and R. solani (MIC in the range of 20–80 µg per paper disc) | [67] |
| 47 | Biscogniauxiamediterranea Ohu 19B | Opuntia humifusa (Cactaceae) from the United States | 5-methylmellein (100) | Compound 100 5-methylmellein was evaluated for antifungal activity against seven plant pathogens (C. acutatum, C. fragariae, C. gloeosporioides, F. oxysporum, B. cinerea, P. obscurans, and P. viticola) using an in vitro microdilution broth assay. | [68] |
| 48 | Emericella sp. XL029 | Leaves of Panax notoginseng Shijiazhuang, Hebei Province, China. | 5-(undeca-3′,5′,7′-trien-1′-yl)furan-2-ol (126) and 5-(undeca-3′,5′,7′-trien-1′-yl)furan-2-carbonate (127) | Compound 126 active against R. solani, V. dahliae, H. maydis, F. oxysporum, F. tricinctum, B. dothidea, and A. fragriae (MIC values from 25 to 3.1 µg/mL), while compound 127 displayed activity against V. dahliae, H. maydis, F. tricinctum, B. dothidea, and A. fragriae (MIC values from 50 to 12.5 µg/mL) | [69] |
| Comounds Produced by Hyphomycetes | |||||
| 49 | Fusarium fujikuroi (WF5), Penicilium chrysogenum WF6, and P. expensum WF7 | Finger millet Plants Arkell Field Station, Arkell, ON, Canada. | 5-hydroxy 2(3H)-benzofuranone (128), dehydrocostus lactone (129) and harpagoside (130) | Compounds 128–130 active against F. graminearum with MIC of 31.25, 250.00 and 31.25 µg/mL, respectively. | [70] |
| 50 | Trichoderma koningiopsis YIM PH30002 | Panax notoginseng. Wenshan, Yunnan Province, China. | Koninginin O (131), koninginin Q (132), 7-O-methylkoninginin D (133) | Compounds 131–132 active against F. oxysporum and P. cucumerina, with an MIC of 128 µg/mL. Compound 133 showed activity against P. cucumerina with an MIC of 128 µg/mL. Nystatin was active with MICs at 32 µg/mL | [71] |
| 51 | Trichoderma koningiopsis YIM PH30002 | Panax notoginseng. Wenshan, Yunnan Province, China. | Koningiopisin C (134) | Compound 134 exhibited in vitro antifungal activity against F. oxysporum, A. panax, F. solani and P. cucumerina with MICs at 32, 64, 32, and 16 µg/mL, respectively | [72] |
| 52 | Trichoderma sp. 09 | Myoporum bontioides | Dichlorodiaportinolide (135), dichlorodiaportin (136) | Compounds 135–136 active against C. musae and Rhizoctoniasolani (MIC values from 6.25 to 150 µg/mL) | [73] |
| 53 | Trichoderma brevicompactum 0248 | Allium sativum | Trichodermin (137) | Compound 137 active against R. solani, B. cinereal, C. lindemuthianum with an EC50 of 0.25, 2.02 and 25.60 µg/mL respectively. Carbendazim showed, antifungal activity against R. solani, B. cinereal, with an EC50 of 0.36 and 10.35 µg/mL respectively | [74] |
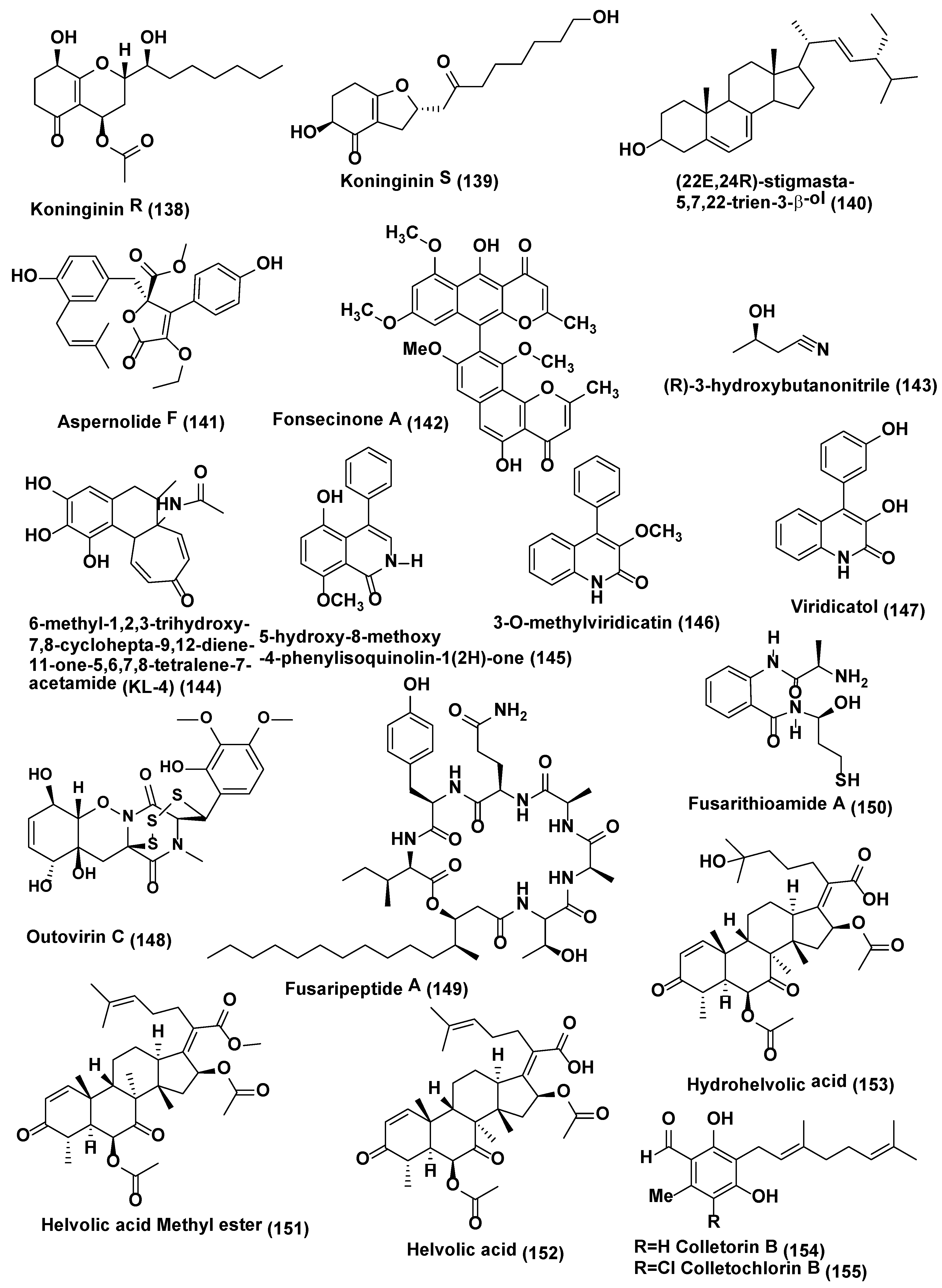
| 54 | Trichoderma koningiopsis YIM PH30002 | Wenshan, Yunnan Province of China. | Koninginin R (138) and S (139) | Compound 138 active against F. oxysporum and F. flocciferum with MICs at 128 µg/mL, while compound 139 displayed activity against F. oxysporum with MIC at 128 µg/mL | [75] |
| 55 | Aspergillus terreus | Carthamus lanatus Al-Azhar University campus, Assiut Branch, Assiut, Egypt. | (22E,24R)-stigmasta-5,7,22-trien-3-β-ol (140), aspernolides F (141) | Compound 140 active against C. neoformans with IC50 values of 4.38 µg/mL, amphotericin B (IC50 0.34 µg/mL). Compound 141 showed good activity against C. neoformans (IC50 5.19 µg/mL). | [76] |
| 56 | Aspergillus sp. KJ-9, | Melia azedarach which was collected at Yangling, Shaanxi Province, China | Fonsecinone A (142), (R)-3-hydroxybutanonitrile (143) | Compounds 142 and 143 were active against G. saubinetti, M. grisea, B. cinerea, C. gloeosporioides and A. solani with MIC range of 6.25–50 µM | [77] |
| 57 | Aspergillus sp. | Gloriosa superba Tirupati, India. | 6-methyl-1,2,3-trihydroxy-7,8-cyclohepta-9,12-diene-11-one-5,6,7,8-tetralene-7-acetamide (KL-4) (144) | KL-4 (144) active against S. cerevisiae, C. albicans and C. gastricus with MIC 25, 12.5, and 50 µg/mL respectively | [78] |
| 58 | Penicillium sp. R22 | Nerium indicum collected from Qinling Mountain, Shaanxi Province, China. | 5-hydroxy-8-methoxy-4-phenylisoquinolin-1(2H)-one (145), 3-O-methylviridicatin (146) and viridicatol (147) | Compound 145 active against A. brassicae, A. alternata and V. mali with MIC value of 31.2 µg/mL, compound 146 against A. brassicae, B. cinerea and V. male with MIC value of 31.2 µg/mL, compound 147 against A. brassicae, A. alternata and B. cinerea with MIC value of 31.2 µg/mL | [79] |
| 59 | Penicillium raciborskii, | Rhododendron tomentosum were collected at the test site of University of Oulu, Finland. | Outovirin C (148) | Outovirin C (148) inhibited growth of F. oxysporum, B. cinerea, and V. dahlia at the concentration of 0.38 µM. Compound 148 active against B. cinerea (57% inhibition) and slightly less effective against V. dahliae (45% inhibition) | [80] |
| 60 | Fusarium sp. | Mentha longifolia Saudi Arabia. | Fusaripeptide A (149) | Compound 149 active against C. albicans, C. glabrata, C. krusei, and A. fumigates with IC50 values of 0.11, 0.24, 0.19, and 0.14 µM, respectively. Amphotericin B exhibited antifungal activity toward C. albicans, C. glabrata, C. krusei, and A. fumigates with IC50 values of 0.3, 0.6, 0.5, 0.7 µM, respectively | [81] |
| 61 | Fusarium chlamydosporium | Anvillea garcinii Al Madinah Al Munawwarah, Saudi Arabia. | Fusarithioamide A (150) | Compound 150 active with inhibition zone diameters 16.2 mm and MIC 2.6 µg/mL towards C. albicans. Clotrimazole (inhibition zone diameters 18.5 mm and MIC 3.7 µg/mL) | [82] |
| 62 | Fusarium sp. | Ficus carica Qinling Mountain, Shaanxi Province, China | Helvolic acid Methyl ester (151), helvolic acid (152) and hydrohelvolic acid (153) | Compounds 151–153 active against B. cinerea, C. gloeosporioides, F. oxysporum f. sp. niveum, F. graminearum and P. capsici (MIC in the range of 12.5–25 µg/mL), Carbendazim (MIC in the range of 32.2–62.5 µg/mL) | [83] |
| 63 | Fusarium sp. | Colletorin B (154), colletochlorin B (155), LL-Z1272β (llicicolin B) (156) and 4,5-dihydrodechloroascochlorin (157) | Compounds 154–156 showed antifungal against U. violacea and F. oxysporum. Compound 157 showed antifungal activity towards E. repens, | Hussain et al. [84] | |
| 64 | Curvularia sp., strain M12, | Murraya koenigii Rajshahi University, Bangladesh | Murranolide A (158), murranopyrone (159), Curvularin (160), (S)-dehydrocurvularin (161), pyrenolide A (162), modiolide A (163), and 8-hydroxy-6-methoxy-3-methylisocoumarin (164) | Pyrenolide A (162) showed a strong motility impairing activity against Phytophthora capsici zoospores at a low concentration (100% at 0.5 µg/mL) in a short time (30 min). Compounds 158–161 and 163–164 exhibited zoospore motility impairment activity at higher concentrations (IC50: 50–100 µg/mL) | [85] |
| 65 | Phaeoacremonium sp., | Senna spectabilis AraraquaraCerrado area, Araraquara, Sao Paulo state, Brazil. | Isoaigialone B (165), and C (166), aigialone (167) | Compounds 165 and 167 exhibited antifungal activity, with a detection limit of 5 µg, for C. cladosporioides and C. sphaerospermum, compound 166 exhibited weak activity (detection limit > 5 µg), with a detection limit of 25 µg. Nystatin, positive control, showing a detection limit of 1 µg | [86] |
| 66 | Trichothecium sp. | Phyllanthus amarus Pune India. | Trichothecinol A (168) | Compound 168 active against C. albidus up to 20 µg/mL | [87] |
| 67 | Trichothecium sp. | Phyllanthus sp. Pune India. | Trichothecin (169) | Trichothecin (169) active against S. cerevisiae, C. albidus var diffluens (NCIM 3371), C. albidus var diffluens (NCIM 3372), F. oxysporum, P. expansum, T. viride, P. varioti and A. niger with MIC of 6.0, 20.0, 12.0, 10.0, 30.0, 40.0, 20.0 and 12.0 µg/mL respectively | [88] |
| Comounds Produced by Basidiomycetes | |||||
| 68 | Scleroderma UFSM Sc1(Persoon) Fries | Eucalyptus grandis. | Sclerodol A (170) and B (171) and related lanostane triterpenoid (172) | Compound 170 active against C. albicans, C. tropicalis, C. crusei, C. parapsiosis (MIC of 25.0, 25.0, 6.25 and 12.5 MFC 25.0 25.0, 12.5, 25.0 µg/mL) Compound 170 and 172 were active against tested strain (MIC in range of 12.5–100 µg/mL). Nystatin active against test strains (MIC in the range of 0.77–1.52 µg/mL). | [89] |
| No. | Name of the Endophytic Fungi | Geographic Area of Isolation | Major VOCs Produced | Anti-Fungal Activity | Refs. |
|---|---|---|---|---|---|
| 1 | Muscodor albus | Central America (Honduras) | 2-methylpropanoic acid; 3-methyl-1-butanol; ethanol; acetic acid (methyl ester) | Rhizoctonia solani; Phytophthora cinnamomi; Sclerotinia sclerotiorum; Fusarium solani, Verticillium dahliae | [91] |
| 2 | M. vitigenus | South America (Peru) | naphthalene; caryophyllene; azulene | R. solani; Phoma sp.; C. coefficola | [98] |
| 3 | M. roseus | Australia | 2-butenoic acid (ethyl ester); 1,2,4-tri-methyl-benzene; 2-nonadiene | Antifungal spectrum not reported | [101] |
| 4 | M. yucatensis | South America (Mexico) | caryophyllene; aromadendrene | Botrytis cineria; R. solani; C. coefficola; Phoma sp. | [102] |
| 5 | M. fengyangensis | China | 2-methylpropionic acid; β-phellendrene | B. cineria; Aspergillus clavatus; Colletotrichum fragiae; Sclerotium rolfsii | [90] |
| 6 | M. crispans | South America (Bolivia) | 2-methylpropanoic acid; ethanol; ethyl acetate | B. cineria; Curvularia lunata; P. cinnamomi; S. sclerotiorum | [103] |
| 7 | M. sutura | USA (Columbia) | butylated hydroxytoleuene; octacecanoic acid; thujopsene; 2-methylpropanoic acid; naphthalene | Aspergillus fumigatus; Colletotrichum lagenarium; B. cineria; Cercospora beticola; Phytophthora palmivora; Fusarium solani | [104] |
| 8 | M. musae | Thailand | 3-methylbutanol acetate 2-methylpropanoic acid | Alternaria porri; Alternaria solani; Colletotrichum gloereosporioides; Nigrospora oryzae | [105] |
| 9 | M. oryzae | Thailand | 3-methylbutan-1-ol; 2-methylpropanoic acid | A. porri; A. solani; Aspergillus flavus; B. cineria; C. gloereosporioides; N. oryzae | [105] |
| 10 | M. suthepensis | Thailand | 3-methylpropanoic acid 3-methylbutan-1-ol | A. porri; Alternaria alternata; Aspergillus flavus; B. cineria; C. gloereosporioides; Fusarium oxysporum; Fusarium solani; N. oryzae | [105] |
| 11 | M. equiseti | Thailand | 3-methylbutan-1-ol; 3-methylbutanoyl acetate; 2-methylpropanoic acid | A. porri; A. solani; B. cineria; C. gloereosporioides; F. oxysporum; F. solani; N. oryzae | [105] |
| 12 | M. cinnamomi | Thailand | 2-methylpropanoic acid;2-methyl butanoic acid; azulene | Rhizoctonia solani | [105] |
| 13 | M. kashayum | India | 1-methyl-4(1-methylethhylidene)-cyclohexane; 2(4-morpholinyl)ethylamine; 9-octadecanoic acid (methyl ester); 4-octadecylmorpholine | Bionectria ochroleuca; Cercospora beticola; Chaetomium heterosporum; C. gloereosporioides; F. oxysporum; Fusarium equiseti; Curvularia lunata | [106] |
| 14 | M. darjeelingensis | India | 4-octadecylmorpholine; 2,6-bis(1,1-dimethylethyl)-4-(1-oxopropyl)phenol; beta-aminoethyl-morpholine | Lasiodiplodia theobromae; A. alternata; Rhizoctonia solani; Cercospora beticola | [107] |
| 15 | M. strobelii | India | 4-octadecylmorpholine; tetraoxapropellan; aspidofractanine-3-methanol;viridiflorol | Rhizoctonia solani; Colletotrichum gloereosporioides; Fusarium oxysporum; Lasiodiplodia theobromae | [108] |
| 16 | M. tigrii | India | 4-octadecylmorpholine; 1-tetradecamine n, n-dimethyl 1,2-benzidicarboxylic acid mono(2-ethylhexyl)ester | Alternaria alternate Cercospora beticola | [109] |
| 17 | M. heavae | Thailand | 2-phenylethanol; azulene | Aspergillus niger; Phellinus noxius; Rigidoporus microporus | [110] |
| 18 | M. ghoomensis | India | n,n-dimethyl-1-nonadecamine; 4-octadecylmorpholine | Cercospora beticola | [111] |
| 19 | M. indica | India | n, n-dimethyl-1-pentadecamine; 4-morpholinethanamine | Cercospora beticola Penicillium marnaeffi | [111] |
| 20 | M. camphora | India | tetracontane; 4-octadecylmorpholine; n, n-dimethyl-1-pentadecamine | Colletotrichum gloereosporioides; Lasiodiplodia theobromae | [112] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deshmukh, S.K.; Gupta, M.K.; Prakash, V.; Saxena, S. Endophytic Fungi: A Source of Potential Antifungal Compounds. J. Fungi 2018, 4, 77. https://doi.org/10.3390/jof4030077
Deshmukh SK, Gupta MK, Prakash V, Saxena S. Endophytic Fungi: A Source of Potential Antifungal Compounds. Journal of Fungi. 2018; 4(3):77. https://doi.org/10.3390/jof4030077
Chicago/Turabian StyleDeshmukh, Sunil K., Manish K. Gupta, Ved Prakash, and Sanjai Saxena. 2018. "Endophytic Fungi: A Source of Potential Antifungal Compounds" Journal of Fungi 4, no. 3: 77. https://doi.org/10.3390/jof4030077
APA StyleDeshmukh, S. K., Gupta, M. K., Prakash, V., & Saxena, S. (2018). Endophytic Fungi: A Source of Potential Antifungal Compounds. Journal of Fungi, 4(3), 77. https://doi.org/10.3390/jof4030077