Molecular Phylogeny and Morphology of Amphisphaeria (= Lepteutypa) (Amphisphaeriaceae)

 ,
,  ,
,  ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection, Isolation and Morphological Studies
2.2. DNA Extraction, PCR Amplification and Sequencing
2.3. Phylogenetic Analyses
2.4. Cluster Analysis
3. Results
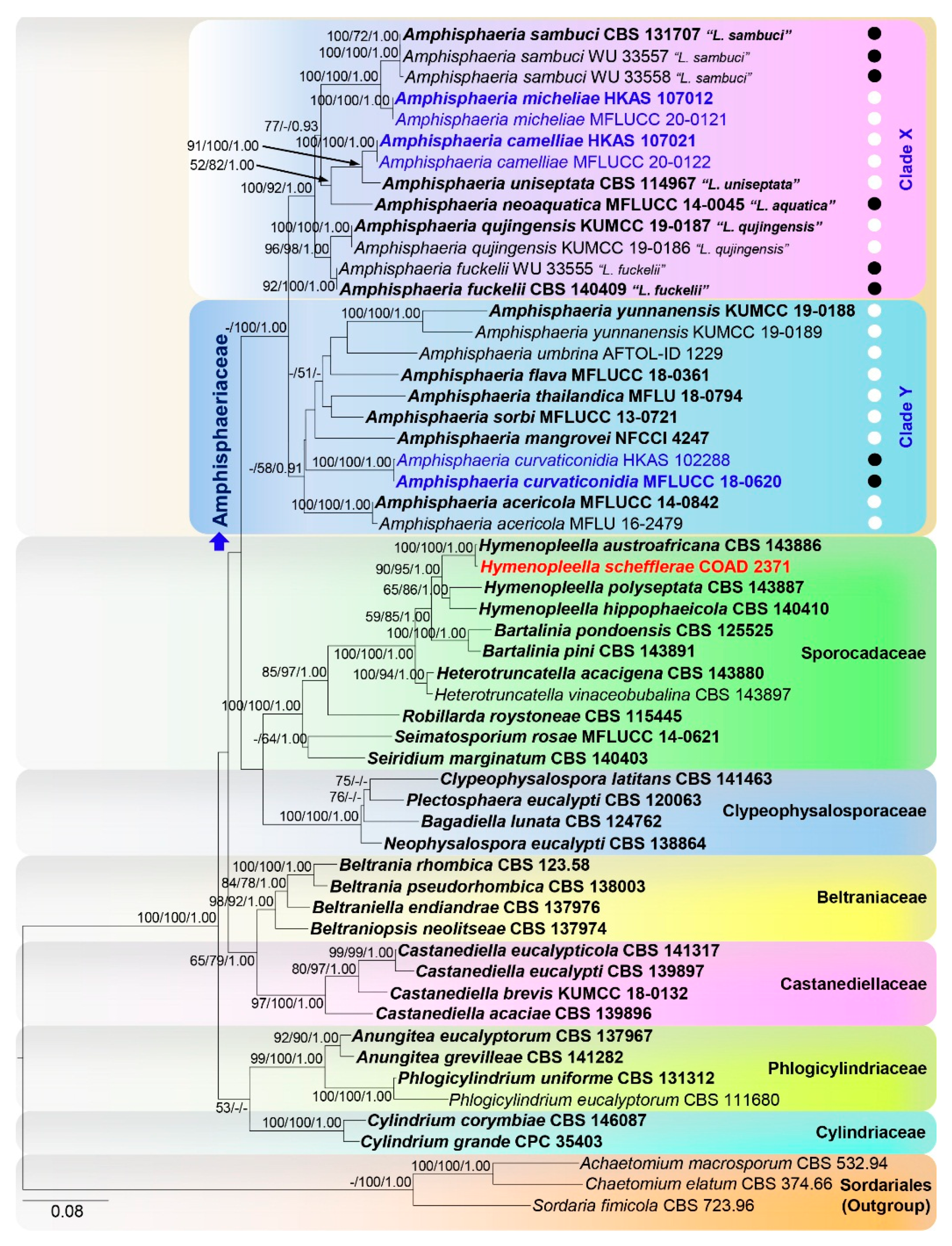
3.1. Topology of Phylogenetic Analyses
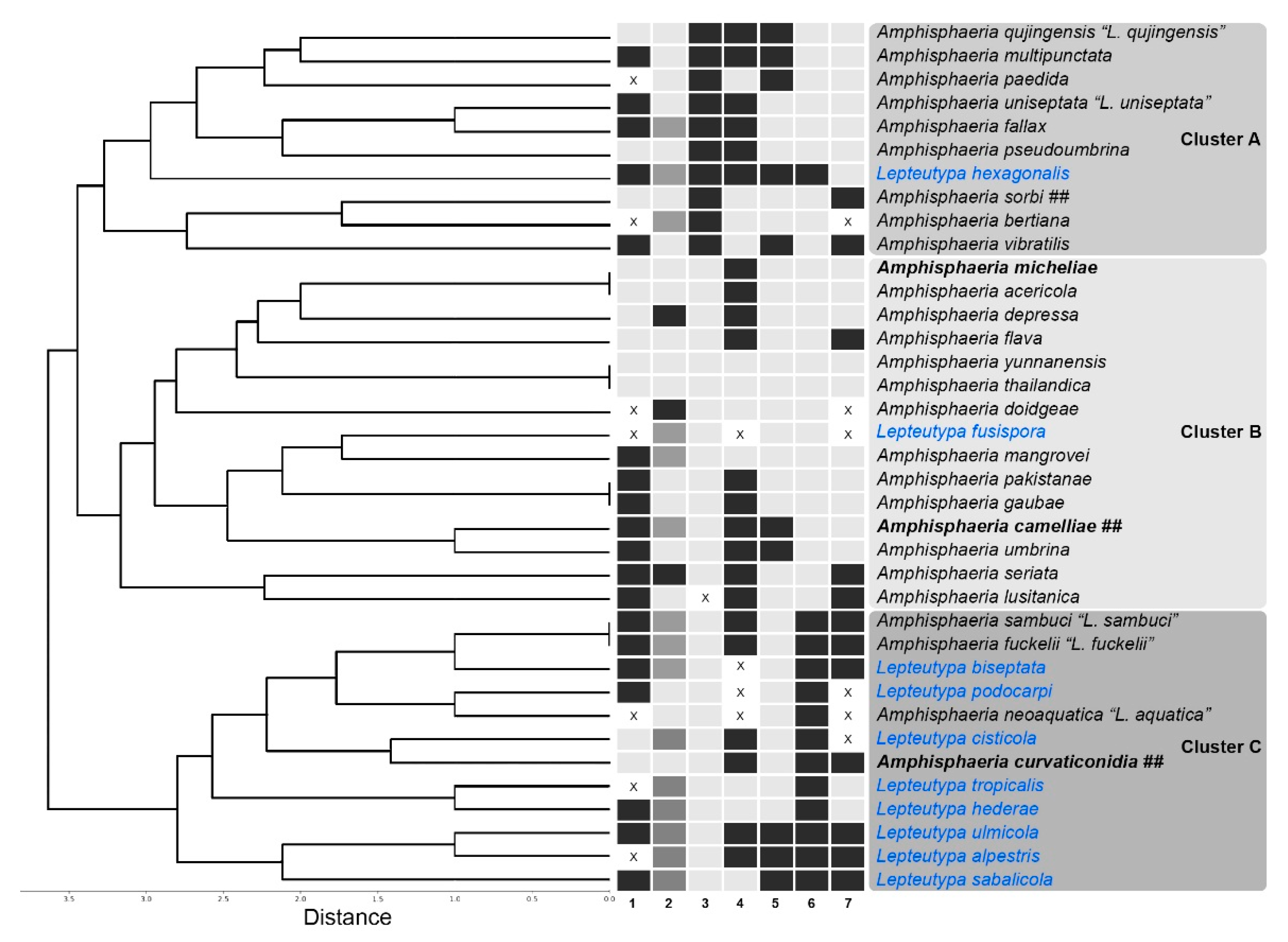
3.2. Cluster Analysis
3.3. Taxonomy
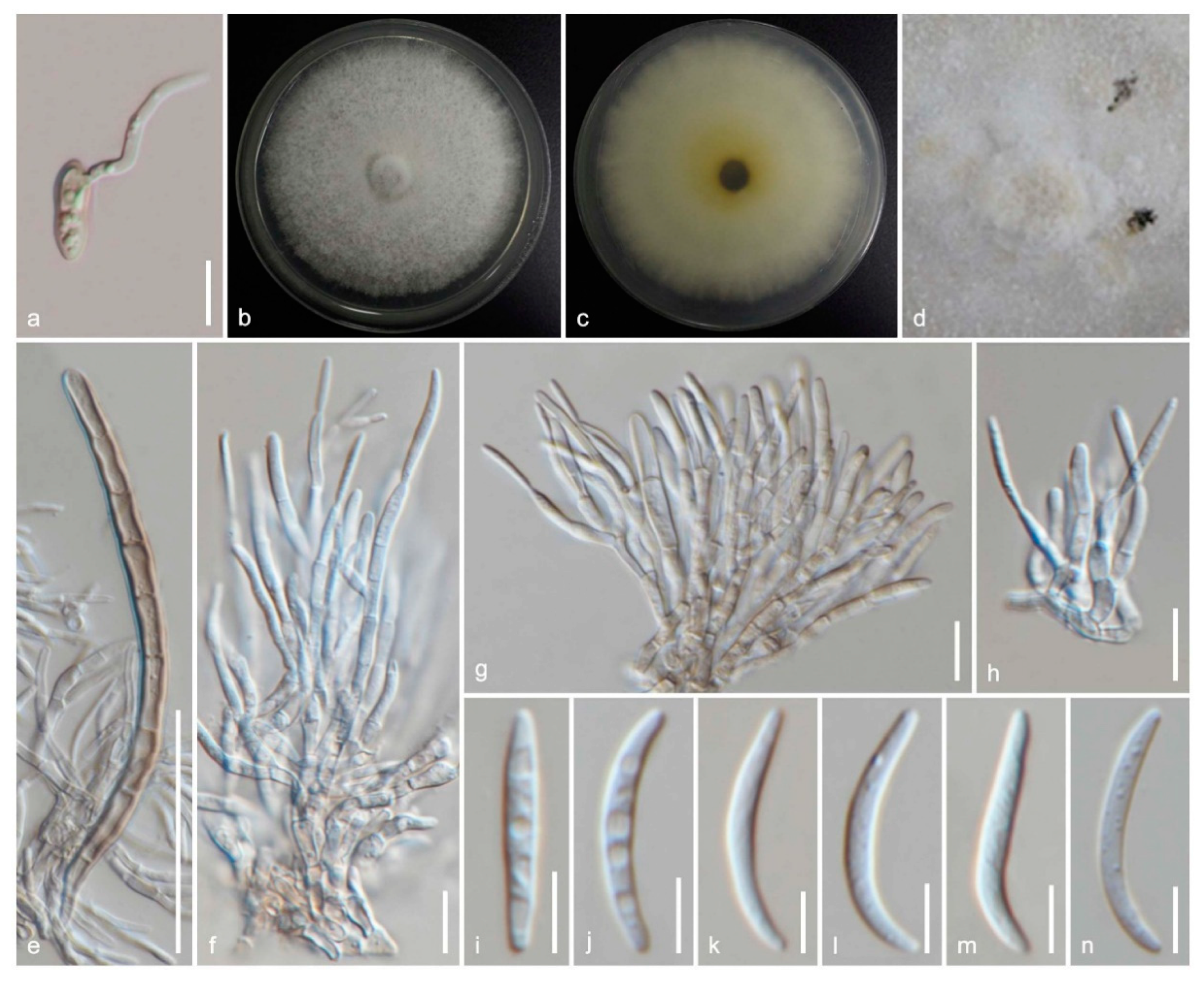
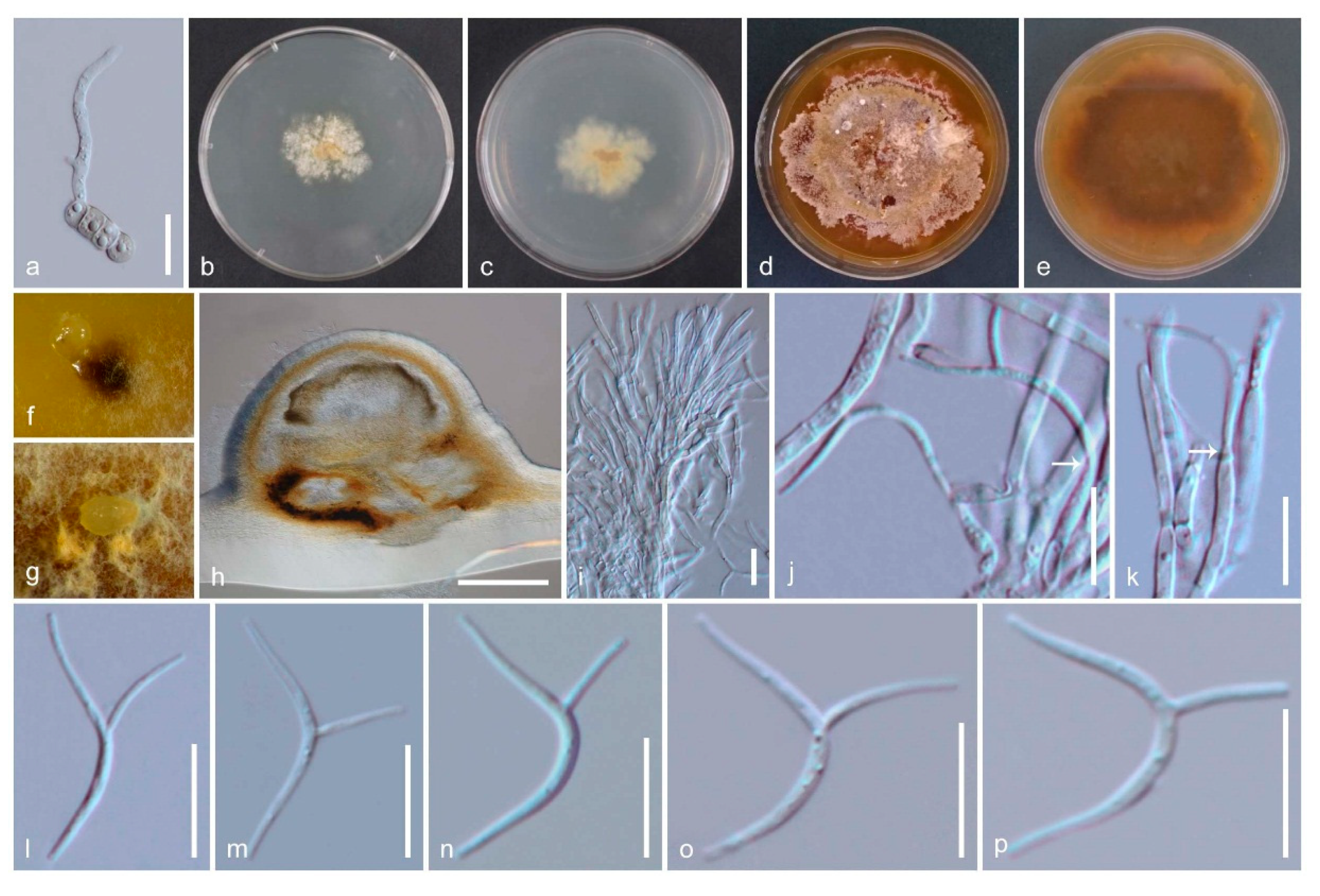
3.3.1. Amphisphaeria
3.3.2. Additional Accepted Species in Amphisphaeria
3.3.3. Taxa Needing Further Revisions
3.3.4. Hymenopleella
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cesati, V.; de Notaris, G. Schema di classificazione degli sferiacei italici aschigeri più o meno appartenenti al genere Sphaeria nell’antico significato attribuitoglda Persoon. Comment. Soc Crittogamia Ital. 1863, 1, 177–240. [Google Scholar]
- Wang, Y.Z.; Aptroot, A.; Hyde, K.D. Revision of the Ascomycete Genus Amphisphaeria; Fungal Diversity Press: Hong Kong, China, 2004; ISBN 962-86765-5-5. [Google Scholar]
- Senanayake, I.C.; Maharachchikumbura, S.S.N.; Hyde, K.D.; Bhat, J.D.; Jones, E.B.G.; McKenzie, E.H.C.; Dai, D.Q.; Daranagama, D.A.; Dayarathne, M.C.; Goonasekara, I.D.; et al. Towards unraveling relationships in Xylariomycetidae (Sordariomycetes). Fungal Divers. 2015, 73, 73–144. [Google Scholar] [CrossRef]
- Marincowitz, S.; Gryzenhout, M.; Wingfield, M.J. New and rare coelomycetes with appendage-bearing conidia from Pondoland, South Africa. Mycotaxon 2010, 111, 309–322. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.K.; Hyde, K.D.; Jones, E.B.G.; Ariyawansa, H.A.; Bhat, D.J.; Boonmee, S.; Maharachchikumbura, S.S.N.; McKenzie, E.H.C.; Phookamsak, R.; Phukhamsakda, C.; et al. Fungal diversity notes 1–110: Taxonomic and phylogenetic contributions to fungal species. Fungal Divers. 2015, 72, 1–197. [Google Scholar] [CrossRef]
- Phookamsak, R.; Hyde, K.D.; Jeewon, R.; Bhat, D.J.; Jones, E.B.G.; Maharachchikumbura, S.S.N.; Raspé, O.; Karunarathna, S.C.; Wanasinghe, D.N.; Hongsanan, S.; et al. Fungal diversity notes 929–1035: Taxonomic and phylogenetic contributions on genera and species of fungi. Fungal Divers. 2019, 95, 1–273. [Google Scholar] [CrossRef] [Green Version]
- Samarakoon, M.C.; Jack Liu, J.-K.; Hyde, K.D.; Promputtha, I. Two new species of Amphisphaeria (Amphisphaeriaceae) from Northern Thailand. Phytotaxa 2019, 391, 207–217. [Google Scholar] [CrossRef]
- Senanayake, I.C.; Lian, T.-T.; Mai, X.-M.; Camporesi, E.; Zeng, Y.-J.; Tian, S.-L.; Xie, N. Taxonomy and phylogeny of Amphisphaeria acericola sp. nov. from Italy. Phytotaxa 2019, 403, 285–292. [Google Scholar] [CrossRef]
- Dissanayake, L.S.; Samarakoon, M.C.; Mortimer, P.E.; Lu, Y.-Z.; Li, Q.-R.; Hyde, K.D.; Kang, J.-C. Morpho-molecular characterization of two novel amphisphaeriaceous species from Yunnan, China. Phytotaxa 2020, 446, 144–158. [Google Scholar] [CrossRef]
- Petrak, F. Mykologische Notizen VI. Ann. Mycol. 1923, 21, 182–335. [Google Scholar]
- Petrak, F. Beiträge zur pilzflora von Hawaii. Sydowia 1953, 7, 381–409. [Google Scholar]
- Petrak, F. Beiträge zur Pilzflora Australiens. Sydowia 1954, 8, 192–220. [Google Scholar]
- Swart, H.J. The fungus causing cypress canker. Trans. Br. Mycol. Soc. 1973, 61, 71–82. [Google Scholar] [CrossRef]
- Barr, M.E. Redisposition of some taxa described by J. B. Ellis. Mycotaxon 1993, 46, 45–76. [Google Scholar]
- Goh, T.-K.; Hyde, K.D. Lepteutypa hexagonalis sp. nov. from Pinanga sp. in Ecuador. Mycol. Res. 1997, 101, 85–88. [Google Scholar] [CrossRef]
- Jaklitsch, W.M.; Gardiennet, A.; Voglmayr, H. Resolution of morphology-based taxonomic delusions: Acrocordiella, Basiseptospora, Blogiascospora, Clypeosphaeria, Hymenopleella, Lepteutypa, Pseudapiospora, Requienella, Seiridium and Strickeria. Persoonia 2016, 37, 82–105. [Google Scholar] [CrossRef] [Green Version]
- Luo, Z.-L.; Hyde, K.D.; Liu, J.-K.; Maharachchikumbura, S.S.N.; Jeewon, R.; Bao, D.-F.; Bhat, D.J.; Lin, C.-G.; Li, W.-L.; Yang, J.; et al. Freshwater Sordariomycetes. Fungal Divers. 2019, 99, 451–660. [Google Scholar] [CrossRef] [Green Version]
- Petrak, F. Mykologische Notizen VIII. Ann. Mycol. 1925, 23, 1–143. [Google Scholar]
- Kirk, P.M.; Cannon, P.F.; Minter, D.W.; Stalpers, J.A. Ainsworth & Bisby’s Dictionary of the Fungi, 10th ed.; CABI Publishing: Wallingford, UK, 2008; ISBN 978-0-85199-826-8. [Google Scholar]
- Hyde, K.D.; Tennakoon, D.S.; Jeewon, R.; Bhat, D.J.; Maharachchikumbura, S.S.N.; Rossi, W.; Leonardi, M.; Lee, H.B.; Mun, H.Y.; Houbraken, J.; et al. Fungal diversity notes 1036–1150: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2019, 96, 1–242. [Google Scholar] [CrossRef]
- Samarakoon, M.C.; Thongbai, B.; Hyde, K.D.; Brönstrup, M.; Beutling, U.; Lambert, C.; Miller, A.N.; Liu, J.-K.; Promputtha, I.; Stadler, M. Elucidation of the life cycle of the endophytic genus Muscodor and its transfer to Induratia in Induratiaceae fam. nov., based on a polyphasic taxonomic approach. Fungal Divers. 2020, 101, 177–210. [Google Scholar] [CrossRef]
- Chomnunti, P.; Hongsanan, S.; Aguirre-Hudson, B.; Tian, Q.; Peršoh, D.; Dhami, M.K.; Alias, A.S.; Xu, J.; Liu, X.; Stadler, M.; et al. The sooty moulds. Fungal Divers. 2014, 66, 1–36. [Google Scholar] [CrossRef]
- Jayasiri, S.C.; Hyde, K.D.; Ariyawansa, H.A.; Bhat, J.; Buyck, B.; Cai, L.; Dai, Y.-C.; Abd-Elsalam, K.A.; Ertz, D.; Hidayat, I.; et al. The Faces of Fungi database: Fungal names linked with morphology, phylogeny and human impacts. Fungal Divers. 2015, 74, 3–18. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols; Elsevier: Amsterdam, The Netherlands, 1990; pp. 315–322. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerase II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- O’Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Nylander, J.A.A. MrModeltest v2. Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Swofford, D.L. PAUP* Version 4.0 b10. Phylogenetic Analysis Using Parsimony (* and Other Methods); Sinauer Associates: Sunderland, UK, 2002. [Google Scholar]
- Kishino, H.; Hasegawa, M. Evaluation of the maximum likelihood estimate of the evolutionary tree topologies from DNA sequence data, and the branching order in Hominoidea. J. Mol. Evol. 1989, 29, 170–179. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; IEEE: Piscataway, NJ, USA; pp. 1–8. [Google Scholar]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [Green Version]
- Zhaxybayeva, O.; Gogarten, J.P. Bootstrap, Bayesian probability and maximum likelihood mapping: Exploring new tools for comparative genome analyses. BMC Genom. 2002, 3, 4. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A. FigTree v. 1.4.0. Available online: http://tree.bio.ed.ac.uk/%0Asoftware/figtree/ (accessed on 15 August 2020).
- Wang, X.W.; Houbraken, J.; Groenewald, J.Z.; Meijer, M.; Andersen, B.; Nielsen, K.F.; Crous, P.W.; Samson, R.A. Diversity and taxonomy of Chaetomium and chaetomium-like fungi from indoor environments. Stud. Mycol. 2016, 84, 145–224. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Bonthond, G.; Groenewald, J.Z.; Cai, L.; Crous, P.W. Sporocadaceae, a family of coelomycetous fungi with appendage-bearing conidia. Stud. Mycol. 2019, 92, 287–415. [Google Scholar] [CrossRef] [PubMed]
- Schoch, C.L.; Sung, G.-H.; López-Giráldez, F.; Townsend, J.P.; Miadlikowska, J.; Hofstetter, V.; Robbertse, B.; Matheny, P.B.; Kauff, F.; Wang, Z.; et al. The Ascomycota tree of life: A phylum-wide phylogeny clarifies the origin and evolution of fundamental reproductive and ecological traits. Syst. Biol. 2009, 58, 224–239. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Shivas, R.G.; Quaedvlieg, W.; van der Bank, M.; Zhang, Y.; Summerell, B.A.; Guarro, J.; Wingfield, M.J.; Wood, A.R.; Alfenas, A.C.; et al. Fungal Planet description sheets: 214–280. Persoonia 2014, 32, 184–306. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Wingfield, M.J.; Guarro, J.; Cheewangkoon, R.; van der Bank, M.; Swart, W.J.; Stchigel, A.M.; Cano-Lira, J.F.; Roux, J.; Madrid, H.; et al. Fungal Planet description sheets: 154–213. Persoonia 2013, 31, 188–296. [Google Scholar] [CrossRef] [PubMed]
- Cheewangkoon, R.; Groenewald, J.Z.; Summerell, B.A.; Hyde, K.D.; To-anun, C.; Crous, P.W. Myrtaceae, a cache of fungal biodiversity. Persoonia 2009, 23, 55–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vu, D.; Groenewald, M.; de Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Guarro, J.; Hernández-Restrepo, M.; Sutton, D.A.; Acharya, K.; Barber, P.A.; Boekhout, T.; Dimitrov, R.A.; Dueñas, M.; et al. Fungal Planet description sheets: 320–370. Persoonia 2015, 34, 167–266. [Google Scholar] [CrossRef]
- Lin, C.-G.; Bhat, D.J.; Liu, J.-K.; Hyde, K.D.; Wang, Y. The genus Castanediella. MycoKeys 2019, 51, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Giraldo, A.; Crous, P.W.; Schumacher, R.K.; Cheewangkoon, R.; Ghobad-Nejhad, M.; Langer, E. The Genera of Fungi—G3: Aleurocystis, Blastacervulus, Clypeophysalospora, Licrostroma, Neohendersonia and Spumatoria. Mycol. Prog. 2017, 16, 325–348. [Google Scholar] [CrossRef] [Green Version]
- Crous, P.W.; Wingfield, M.J.; Schumacher, R.K.; Akulov, A.; Bulgakov, T.S.; Carnegie, A.J.; Jurjević, Ž.; Decock, C.; Denman, S.; Lombard, L.; et al. New and Interesting Fungi. 3. Fungal Syst. Evol. 2020, 6, 157–231. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Carnegie, A.J.; Wingfield, M.J.; Sharma, R.; Mughini, G.; Noordeloos, M.E.; Santini, A.; Shouche, Y.S.; Bezerra, J.D.P.; Dima, B.; et al. Fungal Planet description sheets: 868–950. Persoonia 2019, 42, 291–473. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Wingfield, M.J.; Schumacher, R.K.; Summerell, B.A.; Giraldo, A.; Gené, J.; Guarro, J.; Wanasinghe, D.N.; Hyde, K.D.; Camporesi, E.; et al. Fungal Planet description sheets: 281–319. Persoonia 2014, 33, 212–289. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Summerell, B.A.; Shivas, R.G.; Romberg, M.; Mel’nik, V.A.; Verkley, G.J.M.; Groenewald, J.Z. Fungal Planet description sheets: 92–106. Persoonia 2011, 27, 130–162. [Google Scholar] [CrossRef]
- Quaedvlieg, W.; Verkley, G.J.M.; Shin, H.-D.; Barreto, R.W.; Alfenas, A.C.; Swart, W.J.; Groenewald, J.Z.; Crous, P.W. Sizing up Septoria. Stud. Mycol. 2013, 75, 307–390. [Google Scholar] [CrossRef] [Green Version]
- Summerell, B.A.; Groenewald, J.Z.; Carnegie, A.J.; Summerbell, R.C.; Crous, P. Eucalyptus microfungi known from culture. 2. Alysidiella, Fusculina and Phlogicylindrium genera nova, with notes on some other poorly known taxa. Fungal Divers. 2006, 23, 323–350. [Google Scholar]
- Crous, P.W.; Carris, L.M.; Giraldo, A.; Groenewald, J.Z.; Hawksworth, D.L.; Hemández-Restrepo, M.; Jaklitsch, W.M.; Lebrun, M.-H.; Schumacher, R.K.; Stielow, J.B.; et al. The Genera of Fungi—Fixing the application of the type species of generic names—G2: Allantophomopsis, Latorua, Macrodiplodiopsis, Macrohilum, Milospium, Protostegia, Pyricularia, Robillarda, Rotula, Septoriella, Torula, and Wojnowicia. IMA Fungus 2015, 6, 163–198. [Google Scholar] [CrossRef]
- Bonthond, G.; Sandoval-Denis, M.; Groenewald, J.Z.; Crous, P.W. Seiridium (Sporocadaceae): An important genus of plant pathogenic fungi. Persoonia 2018, 40, 96–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, A.M.C.; Jeewon, R.; Hyde, K.D. Phylogenetic utility of protein (RPB2, β-tubulin) and ribosomal (LSU, SSU) gene sequences in the systematics of Sordariomycetes (Ascomycota, Fungi). Antonie Leeuwenhoek 2007, 91, 327–349. [Google Scholar] [CrossRef]
- Fries, E.M. Systema Mycologicum: Sistens Fungorum Ordines, Genera et Species, huc Usque Cognitas, Quas ad Normam Methodi Naturalis Determinavit/Disposuit Atque Descripsit Elias Fries; Ex Officina Berlingiana: Lund, Sweden, 2018; Volume 2, p. 1823. [Google Scholar]
- Clements, F.E.; Shear, C.L. The Genera of Fungi; Wilson: New York, NY, USA, 1931. [Google Scholar]
- Müller, E.; von Arx, J.A. Die Gattungen der didymosporen Pyrenomyceten. Beiträge Kryptogamenflora Schweiz 1962, 11, 1–922. [Google Scholar]
- Korf, R.P. Report (N.S. 1) of the Committee for Fungi and Lichens on proposals to conserve and/or reject names. Taxon 1988, 37, 450–463. [Google Scholar] [CrossRef]
- Hyde, K.D.; Kang, J.C.; Kong, R.Y.C. Fungi from palms. XXX. Notes on Amphisphaeria species described from palms and a description of A. umbrina. Nov. Hedwig. 1996, 63, 101–108. [Google Scholar]
- Saccardo, P.A.; Traverso, G.B.; Trotter, A. Sylloge Fungorum Omnium Hucusque Cognitorum. Digessit P. A. Saccardo; Sumptibus Auctoris: Patavi, Padova, Italy, 1882. [Google Scholar]
- Ridley, G.S.; Dobbie, K.; Dick, M.A. Mycological records. 5: Lepteutypa podocarpi (Butin) van der Aa. N. Z. J. For. Sci. 1997, 27, 76–78. [Google Scholar]
- Aptroot, A. Redisposition of some species excluded from Didymosphaeria (Ascomycotina). Nov. Hedwig. 1995, 60, 325–379. [Google Scholar]
- Samuels, G.J.; Müller, E.; Petrini, O. Studies in the Amphisphaeriaceae (sensu lato) 3. New species of Monographella and Pestalosphaeria, and two new genera. Mycotaxon 1987, 28, 473–499. [Google Scholar]
- Saccardo, P.A. Fungi Veneti novi vel critici. Series I. Nuovo G. Bot. Ital. 1873, 5, 269–298. [Google Scholar]
- Tsui, C.K.M.; Hyde, K.D.; Hodgkiss, I.J. Paraniesslia tuberculata gen. et sp. nov., and new records or species of Clypeosphaeria, Leptosphaeria and Astrosphaeriella in Hong Kong Freshwater Habitats. Mycologia 2001, 93, 1002–1009. [Google Scholar] [CrossRef] [Green Version]
- Ellis, J.B.; Everhart, B.M. New species of fungi from various localities. Proc. Acad. Nat. Sci. Phila. 1894, 46, 322–384. [Google Scholar]
- Marin-Felix, Y.; Hernández-Restrepo, M.; Iturrieta-González, I.; García, D.; Gené, J.; Groenewald, J.Z.; Cai, L.; Chen, Q.; Quaedvlieg, W.; Schumacher, R.K.; et al. Genera of phytopathogenic fungi: GOPHY 3. Stud. Mycol. 2019, 94, 1–124. [Google Scholar] [CrossRef]
- Nag Raj, T.R.; Kendrick, B. Ellurema gen. nov., with notes on Lepteutypa cisticola and Seiridium canariense. Sydowia Ann. Mycol. Ser. II 1985, 38, 178–193. [Google Scholar]
- Petrak, F. Mykologische Beiträge zur Flora der kanarischen Inseln. Bot. Engler’s Jahrb. Syst. Beibl. 1929, 142, 93–160. [Google Scholar]
- Rappaz, F. Anthostomella and related xylariaceous fungi on hardwoord from Europe and North America. Mycol. Helv. 1995, 7, 99–168. [Google Scholar]
- Butin, H. Beitrag zur Ascomyceten Flora von Chile. Sydowia 1973, 27, 267–292. [Google Scholar]
- van der Aa, H.A. Revision of Keissleriella podocarpi BUTIN. Sydowia 1986, 39, 1–7. [Google Scholar]
- Dulymamode, R.; Cannon, P.F.; Sivanesan, A.; Peerally, A. Fungi from Mauritius: Four new ascomycetes on native plants. Mycol. Res. 2001, 105, 247–254. [Google Scholar] [CrossRef]
- Barr, M.E. Notes on the Amphisphaeriaceae and related families. Mycotaxon 1994, 51, 191–224. [Google Scholar]
- Hirayama, K.; Tanaka, K. Taxonomic revision of Lophiostoma and Lophiotrema based on reevaluation of morphological characters and molecular analyses. Mycoscience 2011, 52, 401–412. [Google Scholar] [CrossRef]
- Jaklitsch, W.M.; Olariaga, I.; Voglmayr, H. Teichospora and the Teichosporaceae. Mycol. Prog. 2016, 15, 31. [Google Scholar] [CrossRef] [Green Version]
- Hyde, K.D.; Norphanphoun, C.; Maharachchikumbura, S.S.N.; Bhat, D.J. Refined families of Sordariomycetes. Mycosphere 2020, 11, 305–1059. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Al-Ani, L.K.T.; Tedersoo, L. Outline of Fungi and fungus-like taxa. Mycosphere 2020, 11, 1060–1456. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Code | GenBank Accession Numbers | References | |||
|---|---|---|---|---|---|---|
| LSU | ITS | RPB2 | TUB2 | |||
| Achaetomium macrosporum | CBS 532.94 | KX976699 | KX976574 | KX976797 | KX976915 | [38] |
| Amphisphaeria acericola | MFLU 16-2479 | MK640424 | MK640423 | - | - | [8] |
| Amphisphaeria acericola | MFLUCC 14-0842 * | MF614131 | MF614128 | - | - | [8] |
| Amphisphaeria camelliae | HKAS 107021 *,a | MT756615 | MT756621 | MT789850 | MT774368 | This study |
| Amphisphaeria camelliae | MFLUCC 20-0122 b | MT756616 | MT756622 | MT789851 | MT774369 | This study |
| Amphisphaeria curvaticonidia | HKAS 102288 a | MT756618 | MT756624 | MT789853 | - | This study |
| Amphisphaeria curvaticonidia | MFLUCC 18-0620 *,b | MT756617 | MT756623 | MT789852 | - | This study |
| Amphisphaeria flava | MFLUCC 18-0361 * | MH971234 | MH971224 | - | MK033638 | [7] |
| Amphisphaeria fuckelii | WU 33555 | KT949903 | KT949903 | - | - | [16] |
| Amphisphaeria fuckelii | CBS 140409 * | KT949902 | KT949902 | MH554918 | MH554677 | [16,39] |
| Amphisphaeria mangrovei | NFCCI-4247 * | MG844275 | MG844283 | - | - | [6] |
| Amphisphaeria micheliae | HKAS 107012 *,a | MT756619 | MT756625 | MT789854 | MT774370 | This study |
| Amphisphaeria micheliae | MFLUCC 20-0121b | MT756620 | MT756626 | MT789855 | MT774371 | This study |
| Amphisphaeria neoaquatica | MFLUCC 14-0045 * | MK835805 | MK828607 | - | - | [17] |
| Amphisphaeria qujingensis | KUMCC 19-0186 | MN707567 | MN707568 | - | - | [9] |
| Amphisphaeria qujingensis | KUMCC 19-0187 * | MN556316 | MN477033 | - | - | [9] |
| Amphisphaeria sambuci | CBS 131707 * | KT949904 | KT949904 | MH554911 | MH704632 | [16,39] |
| Amphisphaeria sambuci | WU 33557 | KT949905 | KT949905 | - | - | [16] |
| Amphisphaeria sambuci | WU 33558 | KT949906 | KT949906 | - | - | [16] |
| Amphisphaeria sorbi | MFLUCC 13-0721 * | KP744475 | KR092797 | - | - | [5] |
| Amphisphaeria thailandica | MFLU 18-0794 * | MH971235 | MH971225 | MK033640 | MK033639 | [7] |
| Amphisphaeria umbrina | AFTOL-ID 1229 | FJ176863 | FJ176809 | - | - | [40] |
| Amphisphaeria uniseptata | CBS 114967 * | MH554197 | - | MH554878 | MH554638 | [39] |
| Amphisphaeria yunnanensis | KUMCC 19-0188 * | MN556306 | MN477177 | - | - | [9] |
| Amphisphaeria yunnanensis | KUMCC 19-0189 | MN550992 | MN550997 | - | - | [9] |
| Anungitea eucalyptorum | CBS 137967 | KJ869176 | KJ869118 | - | - | [41] |
| Anungitea grevilleae | CBS 141282 | KX228304 | KX228252 | - | - | [42] |
| Bagadiella lunata | CBS 124762 * | GQ303300 | GQ303269 | - | - | [43] |
| Bartalinia pini | CBS 143891 * | MH554330 | MH554125 | MH555033 | MH554797 | [39] |
| Bartalinia pondoensis | CBS 125525 * | MH875078 | MH863602 | MH554904 | MH554663 | [39,40] |
| Beltrania pseudorhombica | CBS 138003 * | KJ869215 | KJ869158 | MH555032 | - | [39,41] |
| Beltrania rhombica | CBS 123.58 | MH869260 | MH857718 | MH554899 | MH704631 | [39,44] |
| Beltraniella endiandrae | CBS 137976 * | KJ869185 | KJ869128 | - | - | [41] |
| Beltraniopsis neolitseae | CBS 137974 * | KJ869183 | KJ869126 | - | - | [41] |
| Castanediella acaciae | CBS 139896 * | KR476763 | KR476728 | - | - | [45] |
| Castanediella brevis | KUMCC 18-0132 * | MH806358 | MH806361 | - | - | [46] |
| Castanediella eucalypti | CBS 139897 * | KR476758 | KR476723 | - | - | [45] |
| Castanediella eucalypticola | CBS 141317 * | KX228317 | KX228266 | - | KX228382 | [42] |
| Chaetomium elatum | CBS 374.66 | MH870466 | KC109758 | KF001820 | KC109776 | [38,44] |
| Clypeophysalospora latitans | CBS 141463 * | KX820261 | KX820250 | - | - | [47] |
| Cylindrium corymbiae | CBS 146087 * | MT223887 | MT223792 | MT223679 | MT223732 | [48] |
| Cylindrium grande | CPC 35403 * | MK876425 | MK876384 | MK876481 | MK876502 | [49] |
| Heterotruncatella acacigena | CBS 143880 * | MH554295 | MH554084 | MH554996 | MH554756 | [39] |
| Heterotruncatella vinaceobubalina | CBS 143897 | MH554341 | MH554139 | MH555045 | MH554812 | [39] |
| Hymenopleella hippophaeicola | CBS 140410 * | KT949901 | KT949901 | MH554919 | MH554678 | [16,39] |
| Hymenopleella polyseptata | CBS 143887 * | MH554321 | MH554116 | MH555024 | MH554789 | [39] |
| Hymenopleella schefflerae | COAD 2371 * | MH084761 | MH128360 | - | MH231215 | [20] |
| Hymenopleella austroafricana | CBS 143886 * | MH554320 | MH554115 | MH555023 | MH554788 | [39] |
| Neophysalospora eucalypti | CBS 138864 * | KP004490 | KP004462 | - | - | [50] |
| Phlogicylindrium uniforme | CBS 131312 * | JQ044445 | JQ044426 | MH554910 | MH704634 | [39,51] |
| Phlogicylindrium eucalyptorum | CBS 111680 | KF251707 | KF251204 | KF252209 | KF252698 | [52] |
| Plectosphaera eucalypti | CBS 120063 * | DQ923538 | DQ923538 | - | - | [53] |
| Robillarda roystoneae | CBS 115445 * | MH874545 | KR873254 | MH554880 | KR873317 | [39,44,54] |
| Seimatosporium rosae | MFLUCC 14-0621 * | MH823070 | LT853105 | LT853153 | LT853253 | [3,55] |
| Seiridium marginatum | CBS 140403 * | MH878679 | KT949914 | LT853149 | LT853249 | [16,44,55] |
| Sordaria fimicola | CBS 723.96 | MH874231 | - | DQ368647 | - | [44,56] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Samarakoon, M.C.; Maharachchikumbura, S.S.N.; Liu, J.-K.; Hyde, K.D.; Promputtha, I.; Stadler, M. Molecular Phylogeny and Morphology of Amphisphaeria (= Lepteutypa) (Amphisphaeriaceae). J. Fungi 2020, 6, 174. https://doi.org/10.3390/jof6030174
Samarakoon MC, Maharachchikumbura SSN, Liu J-K, Hyde KD, Promputtha I, Stadler M. Molecular Phylogeny and Morphology of Amphisphaeria (= Lepteutypa) (Amphisphaeriaceae). Journal of Fungi. 2020; 6(3):174. https://doi.org/10.3390/jof6030174
Chicago/Turabian StyleSamarakoon, Milan C., Sajeewa S. N. Maharachchikumbura, Jian-Kui (Jack) Liu, Kevin D. Hyde, Itthayakorn Promputtha, and Marc Stadler. 2020. "Molecular Phylogeny and Morphology of Amphisphaeria (= Lepteutypa) (Amphisphaeriaceae)" Journal of Fungi 6, no. 3: 174. https://doi.org/10.3390/jof6030174
APA StyleSamarakoon, M. C., Maharachchikumbura, S. S. N., Liu, J. -K., Hyde, K. D., Promputtha, I., & Stadler, M. (2020). Molecular Phylogeny and Morphology of Amphisphaeria (= Lepteutypa) (Amphisphaeriaceae). Journal of Fungi, 6(3), 174. https://doi.org/10.3390/jof6030174