Designed Antimicrobial Peptides Against Trauma-Related Cutaneous Invasive Fungal Wound Infections

Abstract
:1. Introduction
2. Materials and Methods
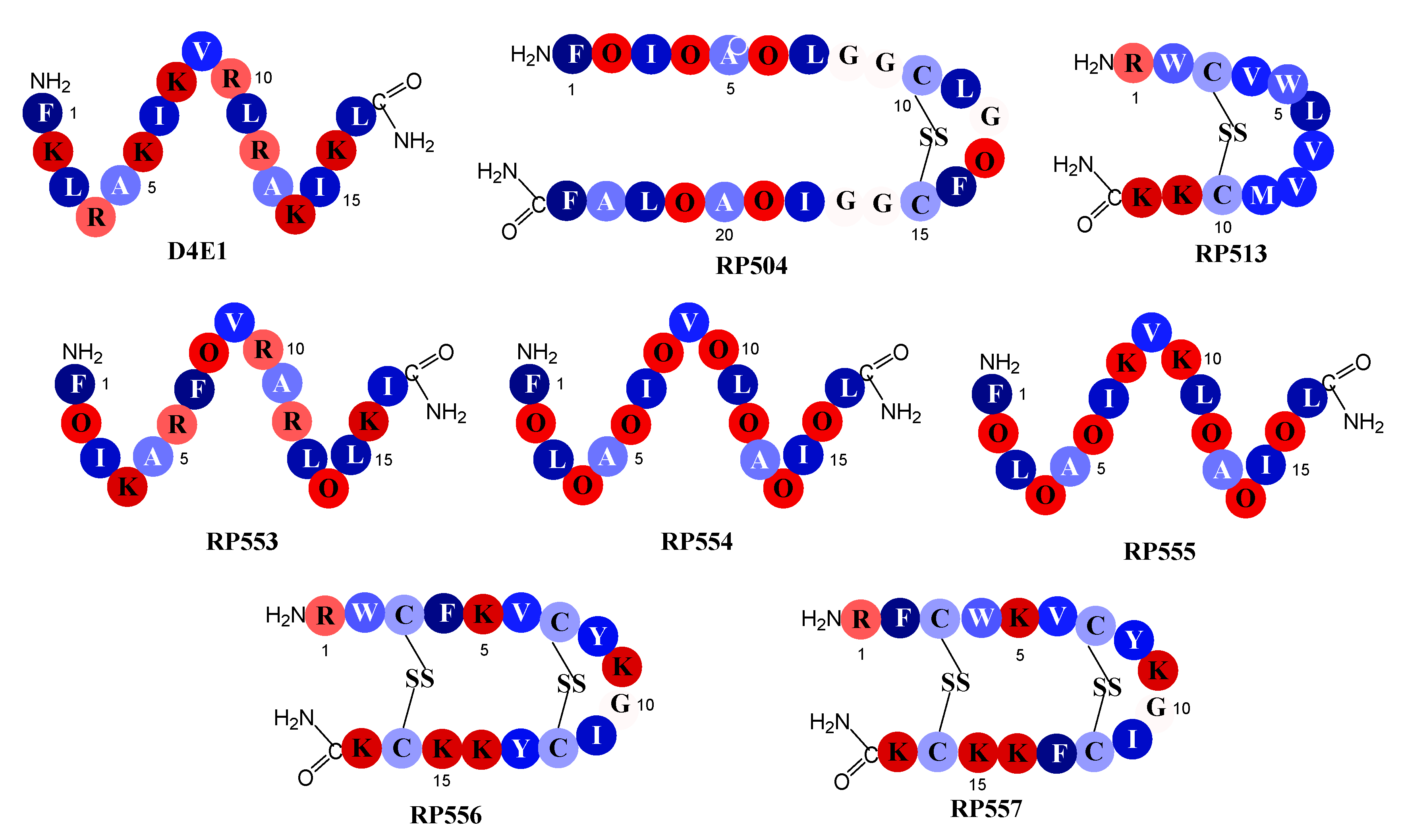
2.1. Designed Antimicrobial Peptides
2.2. Pathogens
2.3. Minimum Inhibitory Concentration (MIC) Assays
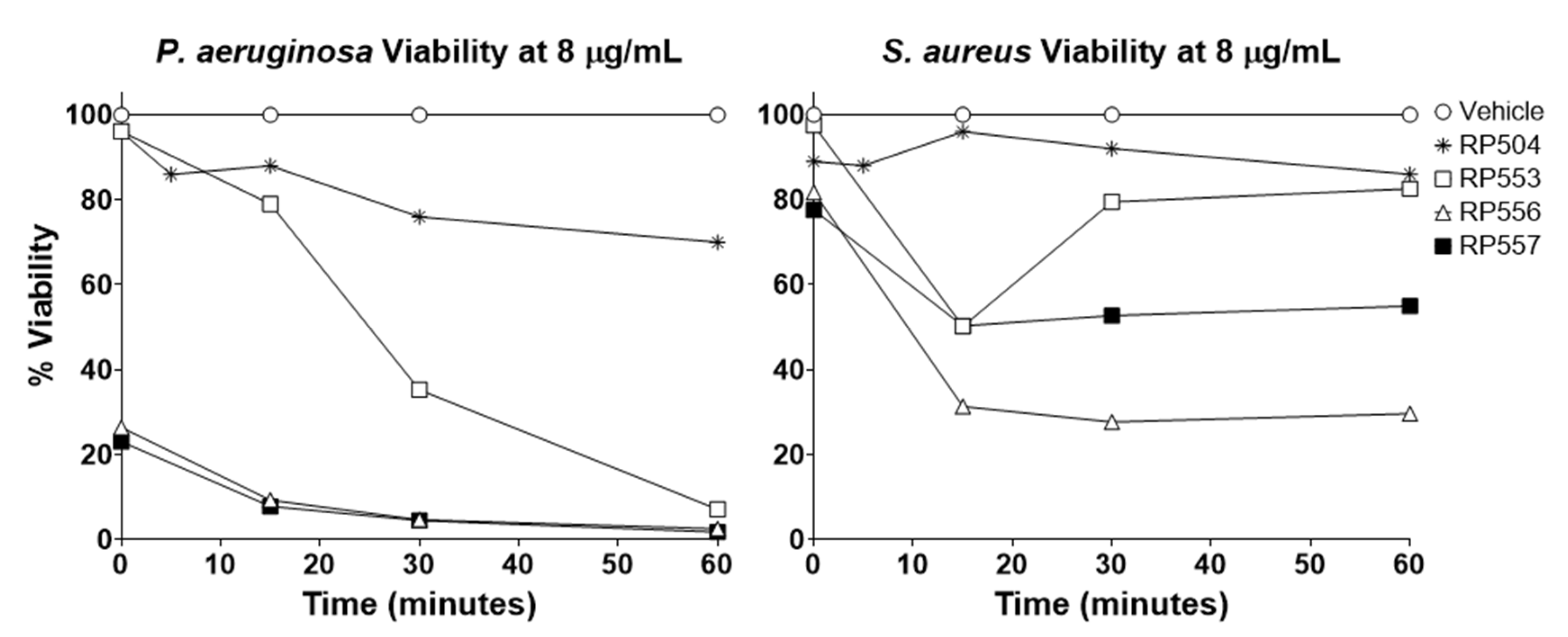
2.4. Bioluminescent Time-kill Assays
2.5. Statistical Analysis
3. Results
3.1. dAMP Antifungal Activity
3.2. dAMP Antibacterial Activity
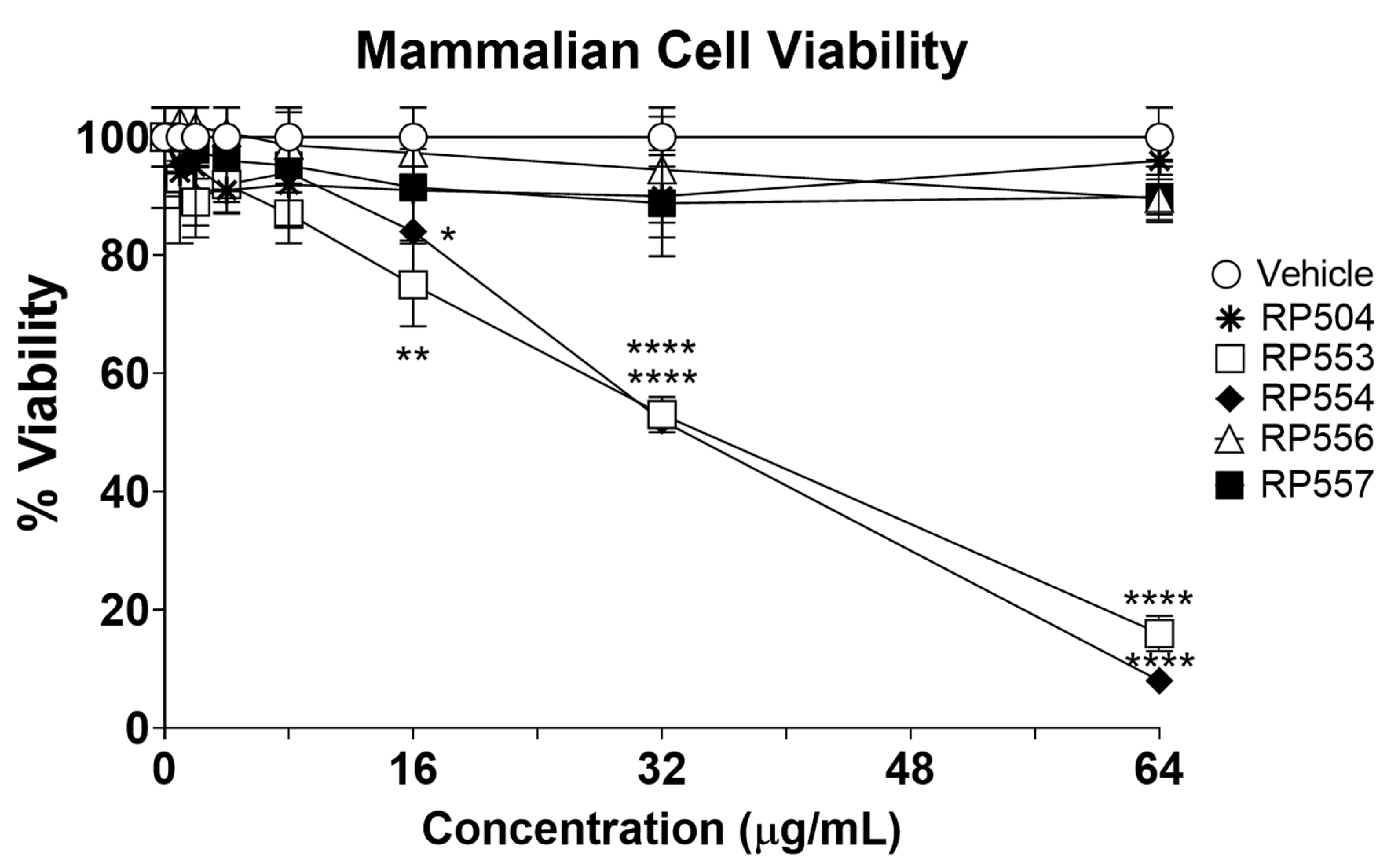
3.3. Limited Mammalian Cell Toxicity
4. Discussion
5. Patents
Author Contributions
Funding
Conflicts of Interest
References
- Rodriguez, C.J.; Weintrob, A.C.; Shah, J.; Malone, D.; Dunne, J.R.; Weisbrod, A.B.; Lloyd, B.A.; Warkentien, T.E.; Murray, C.K.; Wilkins, K.; et al. Risk factors associated with invasive fungal infections in combat trauma. Surg. Infect. 2014, 15, 521–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tribble, D.R.; Rodriguez, C.J. Combat-Related Invasive Fungal Wound Infections. Curr. Fungal Infect. Rep. 2014, 8, 277–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blyth, D.M.; Yun, H.C.; Tribble, D.R.; Murray, C.K. Lessons of war: Combat-related injury infections during the Vietnam War and Operation Iraqi and Enduring Freedom. J. Trauma Acute Care Surg. 2015, 79, S227–S235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganesan, A.; Wells, J.; Shaikh, F.; Peterson, P.; Bradley, W.; Carson, M.L.; Petfield, J.L.; Klassen-Fischer, M.; Akers, K.S.; Downing, K.; et al. Molecular Detection of Filamentous Fungi in Formalin-Fixed Paraffin-Embedded Specimens in Invasive Fungal Wound Infections Is Feasible with High Specificity. J. Clin. Microbiol. 2019, 58, e01259-19. [Google Scholar] [CrossRef]
- Tribble, D.R.; Rodriguez, C.J.; Weintrob, A.C.; Shaikh, F.; Aggarwal, D.; Carson, M.L.; Murray, C.K.; Masuoka, P. Environmental Factors Related to Fungal Wound Contamination after Combat Trauma in Afghanistan, 2009–2011. Emerg. Infect. Dis. 2015, 21, 1759–1769. [Google Scholar] [CrossRef] [Green Version]
- Andersen, R.C.; Fleming, M.E.; Forsberg, J.A.; Gordon, W.T.; Nanos, G.P.; Charlton, M.T.; Ficke, J.R. Dismounted complex blast injury. J. Surg. Orthop. Adv. 2012, 21, 2–7. [Google Scholar]
- Cannon, J.W.; Hofmann, L.J.; Glasgow, S.C.; Potter, B.K.; Rodriguez, C.J.; Cancio, L.C.; Rasmussen, T.E.; Fries, C.A.; Davis, M.R.; Jezior, J.R.; et al. Dismounted Complex Blast Injuries: A Comprehensive Review of the Modern Combat Experience. J. Am. Coll. Surg. 2016, 223, 652–664. [Google Scholar] [CrossRef] [PubMed]
- Lewandowski, L.R.; Weintrob, A.C.; Tribble, D.R.; Rodriguez, C.J.; Petfield, J.L.; Lloyd, B.A.; Murray, C.K.; Stinner, D.J.; Aggarwal, D.; Shaikh, F.; et al. Early Complications and Outcomes in Combat Injury-Related Invasive Fungal Wound Infections: A Case-Control Analysis. J. Orthop. Trauma 2016, 30, e93–e99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganesan, A.; Shaikh, F.; Bradley, W.; Blyth, D.M.; Bennett, D.; Petfield, J.L.; Carson, M.L.; Wells, J.M.; Tribble, D.R. Classification of Trauma-Associated Invasive Fungal Infections to Support Wound Treatment Decisions. Emerg. Infect. Dis. 2019, 25, 1639–1647. [Google Scholar] [CrossRef]
- Warkentien, T.; Rodriguez, C.; Lloyd, B.; Wells, J.; Weintrob, A.; Dunne, J.R.; Ganesan, A.; Li, P.; Bradley, W.; Gaskins, L.J.; et al. Invasive mold infections following combat-related injuries. Clin. Infect. Dis. 2012, 55, 1441–1449. [Google Scholar] [CrossRef] [Green Version]
- Fanfair, R.N.; Benedict, K.; Bos, J.; Bennett, S.D.; Lo, Y.; Adebanjo, T.; Etienne, K.A.; Deak, E.; Derado, G.; Shieh, W.J.; et al. Necrotizing cutaneous mucormycosis after a tornado in Joplin, Missouri, in 2011. N. Engl. J. Med. 2012, 367, 2214–2225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheean, A.J.; Tintle, S.M.; Rhee, P.C. Soft tissue and wound management of blast injuries. Curr. Rev. Musculoskelet. Med. 2015, 8, 265–271. [Google Scholar] [CrossRef] [Green Version]
- Barsoumian, A.E.; Sanchez, C.J.; Mende, K.; Tully, C.C.; Beckius, M.L.; Akers, K.S.; Wenke, J.C.; Murray, C.K. In vitro toxicity and activity of Dakin’s solution, mafenide acetate, and amphotericin B on filamentous fungi and human cells. J. Orthop. Trauma 2013, 27, 428–436. [Google Scholar] [CrossRef]
- Akers, K.S.; Rowan, M.P.; Niece, K.L.; Graybill, J.C.; Mende, K.; Chung, K.K.; Murray, C.K. Antifungal wound penetration of amphotericin and voriconazole in combat-related injuries: Case report. BMC Infect. Dis. 2015, 15, 184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zasloff, M. Antimicrobial peptides of multicellular organisms. Nature 2002, 415, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Clemens, L.E.; Jaynes, J.; Lim, E.; Kolar, S.S.; Reins, R.Y.; Baidouri, H.; Hanlon, S.; McDermott, A.M.; Woodburn, K.W. Designed Host Defense Peptides for the Treatment of Bacterial Keratitis. Investig. Ophthalmol. Vis. Sci. 2017, 58, 6273–6281. [Google Scholar] [CrossRef] [PubMed]
- Woodburn, K.W.; Clemens, L.E.; Jaynes, J.; Joubert, L.M.; Botha, A.; Nazik, H.; Stevens, D.A. Designed Antimicrobial Peptides for Recurrent Vulvovaginal Candidiasis Treatment. Antimicrob. Agents Chemother. 2019, 63, e02690-18. [Google Scholar] [CrossRef] [Green Version]
- Woodburn, K.W.; Jaynes, J.M.; Clemens, L.E. Evaluation of the Antimicrobial Peptide, RP557, for the Broad-Spectrum Treatment of Wound Pathogens and Biofilm. Front. Microbiol. 2019, 10, 1688. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Li, X.; Wang, Z. APD3: The antimicrobial peptide database as a tool for research and education. Nucleic Acids Res. 2016, 44, D1087–D1093. [Google Scholar] [CrossRef] [Green Version]
- de Lucca, A.J.; Bland, J.M.; Grimm, C.; Jacks, T.J.; Cary, J.W.; Jaynes, J.M.; Cleveland, T.E.; Walsh, T.J. Fungicidal properties, sterol binding, and proteolytic resistance of the synthetic peptide D4E1. Can. J. Microbiol. 1988, 44, 514–520. [Google Scholar] [CrossRef]
- Gordon, Y.J.; Romanowski, E.G.; McDermott, A.M. A review of antimicrobial peptides and their therapeutic potential as anti-infective drugs. Curr. Eye Res. 2005, 30, 505–515. [Google Scholar] [CrossRef] [PubMed]
- Heyden, M.; Freites, J.A.; Ulmschneider, M.B.; White, S.H.; Tobias, D.J. Assembly and Stability of α-Helical Membrane Proteins. Soft Matter 2012, 8, 7742–7752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadurugamuwa, J.L.; Sin, L.; Albert, E.; Yu, J.; Francis, K.; DeBoer, M.; Rubin, M.; Bellinger-Kawahara, C.; Parr, T.R., Jr.; Contag, P.R. Direct continuous method for monitoring biofilm infection in a mouse model. Infect. Immun. 2003, 71, 882–890. [Google Scholar] [CrossRef] [Green Version]
- Woodburn, K.W.; Jaynes, J.; Clemens, L.E. Designed Antimicrobial Peptides for Topical Treatment of Antibiotic Resistant Acne Vulgaris. Antibiotics 2020, 9, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berthold, N.; Czihal, P.; Fritsche, S.; Sauer, U.; Schiffer, G.; Knappe, D.; Alber, G.; Hoffmann, R. Novel apidaecin 1b analogs with superior serum stabilities for treatment of infections by gram-negative pathogens. Antimicrob. Agents Chemother. 2013, 57, 402–409. [Google Scholar] [CrossRef] [Green Version]
- Akers, K.S.; Mende, K.; Cheatle, K.A.; Zera, W.C.; Yu, X.; Beckius, M.L.; Aggarwal, D.; Li, P.; Sanchez, C.J.; Wenke, J.C.; et al. Biofilms and persistent wound infections in United States military trauma patients: A case–control analysis. BMC Infect. Dis. 2014, 14, 190. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Fungi | D4E1 | RP504 | RP513 | RP553 | RP554 | RP555 | RP556 | RP557 |
|---|---|---|---|---|---|---|---|---|
| Candida albicans Y-326 | 64 | 8 | 64 | 64 | 32 | 32 | 16 | 16 |
| Candida albicans Y-6359 | 64 | 4 | 64 | 64 | 32 | 32 | 8 | 4 |
| Candida parapsilosis Y-1761 | 64 | 8 | >128 | 64 | 16 | 32 | 32 | 16 |
| Candida parapsilosis Y-1763 | 64 | 2 | >128 | 32 | 8 | 16 | 2 | 2 |
| Candida krusei Y-27803 | 64 | 8 | 64 | 32 | 16 | 32 | 16 | 16 |
| Candida krusei Y-27825 | 64 | 8 | 64 | 32 | 16 | 32 | 32 | 16 |
| Aspergillus fumigatus 9648 | 128 | >128 | 128 | 128 | 64 | 64 | 64 | 64 |
| Aspergillus fumigatus 9651 | 64 | >128 | >128 | 64 | 32 | 64 | 64 | 64 |
| Aspergillus flavus MYA-3651 | 128 | >128 | 128 | 128 | 32 | 128 | 64 | 64 |
| Aspergillus flavus MYA-1004 | 128 | >128 | >128 | 128 | 64 | >128 | 64 | 64 |
| Absidia corymbifera NRRL 6251 | 2 | 8 | 32 | 2 | 4 | 4 | 2 | 2 |
| Fusarium solani NRRL 28548 | 2 | 2 | 16 | 2 | 2 | 2 | 2 | 2 |
| Mucor circinelloides NRRL 3631 | 128 | >128 | 32 | 64 | 32 | 64 | 64 | 64 |
| dAMPs | D4E1 | RP504 | RP513 | RP553 | RP554 | RP555 | RP556 | RP557 |
|---|---|---|---|---|---|---|---|---|
| Gram-negative bacteria | ||||||||
| A. baumannii 6043 | >128 | 32 | 64 | 16 | 8 | 16 | 8 | 8 |
| A. baumannii 6838 | 8 | 64 | 128 | 8 | 4 | 4 | 8 | 8 |
| A. baumannii ATCC 17978 | 16 | 32 | >128 | 4 | 4 | 4 | 8 | 16 |
| E. cloacae 6053 | 8 | 32 | >128 | 4 | 4 | 2 | 8 | 8 |
| E. cloacae 6054 | 4 | 64 | >128 | 2 | 2 | 4 | 8 | 8 |
| K. pneumonia 6069 | 16 | 32 | 128 | 4 | 16 | 16 | 8 | 4 |
| K. pneumonia ATCC 10031 | 32 | 32 | 128 | 4 | 8 | 4 | 8 | 8 |
| P. aeruginosa 6186 | 16 | 16 | >128 | 8 | 8 | 4 | 4 | 4 |
| P. aeruginosa ATCC 19660 | 32 | 16 | >128 | 8 | 8 | 8 | 8 | 4 |
| P. aeruginosa ATCC 27853 | 16 | 16 | >128 | 4 | 4 | 4 | 8 | 4 |
| Gram-positive bacteria | ||||||||
| S. aureus B-767 | 4 | 32 | >128 | 4 | 4 | 8 | 8 | 4 |
| S. aureus 6061 | 8 | 64 | >128 | 8 | 32 | 16 | 16 | 8 |
| MRSA 6313 | 8 | 64 | >128 | 4 | 32 | 16 | 8 | 4 |
| MRSA 6381 | 8 | 64 | >128 | 8 | 32 | 16 | 4 | 4 |
| MRSA ATCC 33592 | 8 | 32 | >128 | 4 | 32 | 8 | 4 | 4 |
| S. epidermis ATCC 51625 | 4 | 16 | >128 | 2 | 2 | 2 | 4 | 8 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Woodburn, K.W.; Jaynes, J.M.; Clemens, L.E. Designed Antimicrobial Peptides Against Trauma-Related Cutaneous Invasive Fungal Wound Infections. J. Fungi 2020, 6, 184. https://doi.org/10.3390/jof6030184
Woodburn KW, Jaynes JM, Clemens LE. Designed Antimicrobial Peptides Against Trauma-Related Cutaneous Invasive Fungal Wound Infections. Journal of Fungi. 2020; 6(3):184. https://doi.org/10.3390/jof6030184
Chicago/Turabian StyleWoodburn, Kathryn W., Jesse M. Jaynes, and L. Edward Clemens. 2020. "Designed Antimicrobial Peptides Against Trauma-Related Cutaneous Invasive Fungal Wound Infections" Journal of Fungi 6, no. 3: 184. https://doi.org/10.3390/jof6030184
APA StyleWoodburn, K. W., Jaynes, J. M., & Clemens, L. E. (2020). Designed Antimicrobial Peptides Against Trauma-Related Cutaneous Invasive Fungal Wound Infections. Journal of Fungi, 6(3), 184. https://doi.org/10.3390/jof6030184