Histidine-Rich Defensins from the Solanaceae and Brasicaceae Are Antifungal and Metal Binding Proteins

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of Additional HRDs
2.2. Sequence Analysis
2.3. Confirmation of Expression of HRDs in Planta
2.4. Cloning, Expression, and Purification of HRDs
2.5. Antifungal Activity Assays
2.6. Metal-induced Precipitation
2.7. Microscale Thermophoresis
2.8. NMR Spectroscopy and Structural Analysis
2.9. Molecular Modeling
3. Results
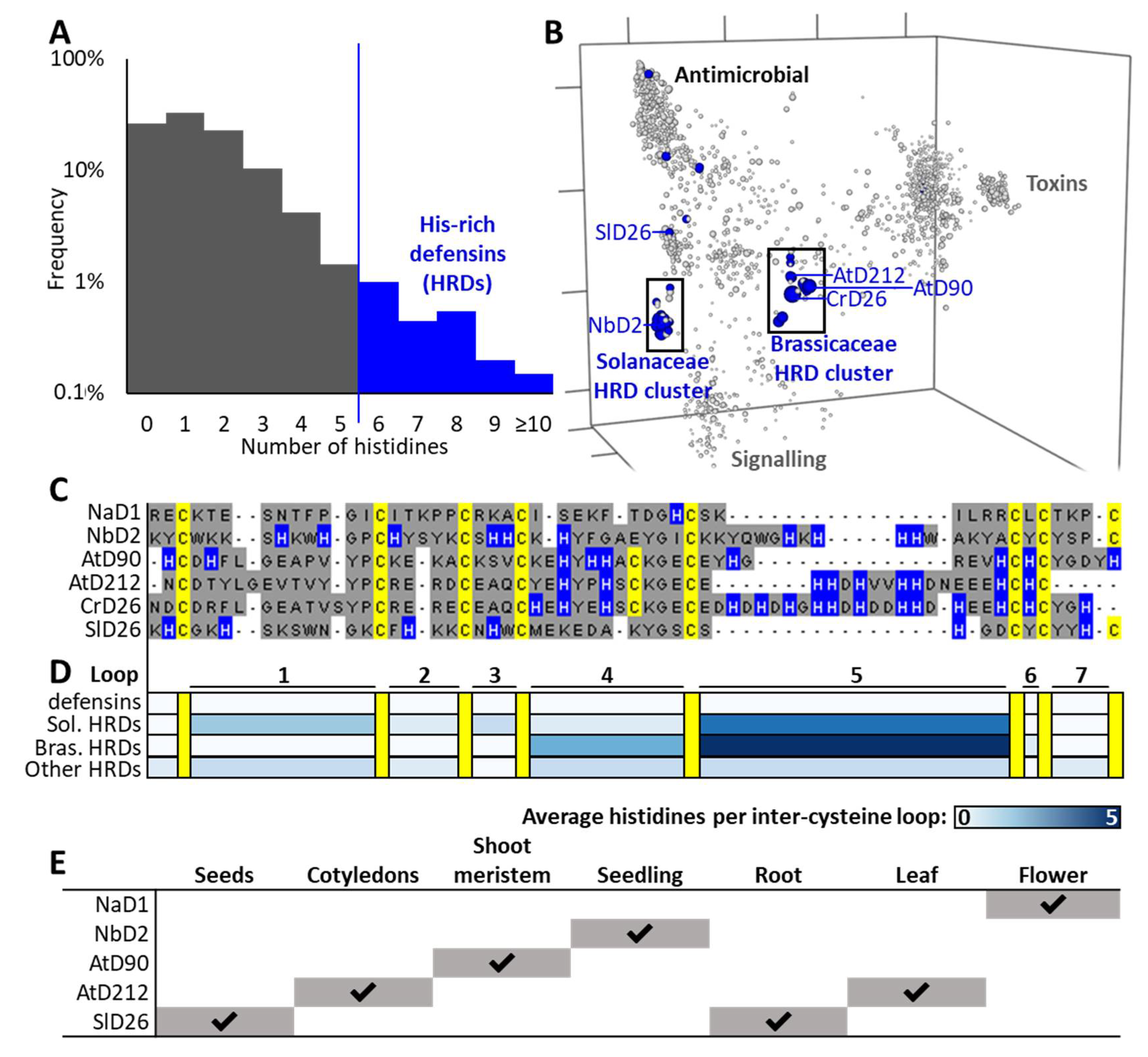
3.1. Identification of Histidine-Rich Defensins
3.2. Evolutionary Analyses
3.3. Expression and Purification of HRDs
3.4. Antifungal Activity of HRDs
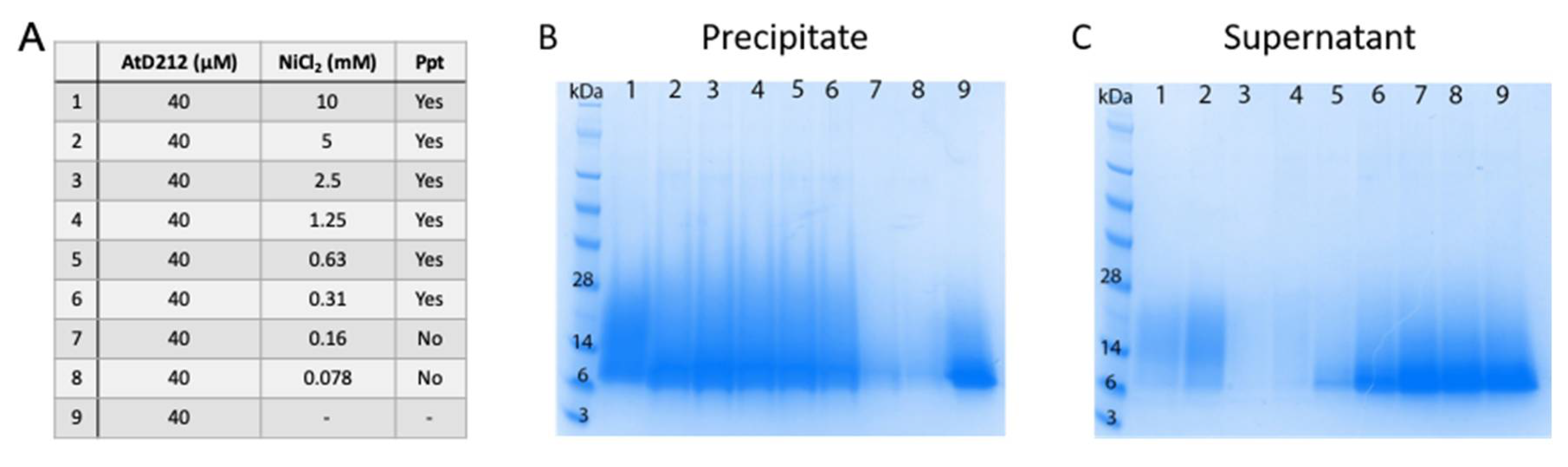
3.5. Metal Binding by HRDs
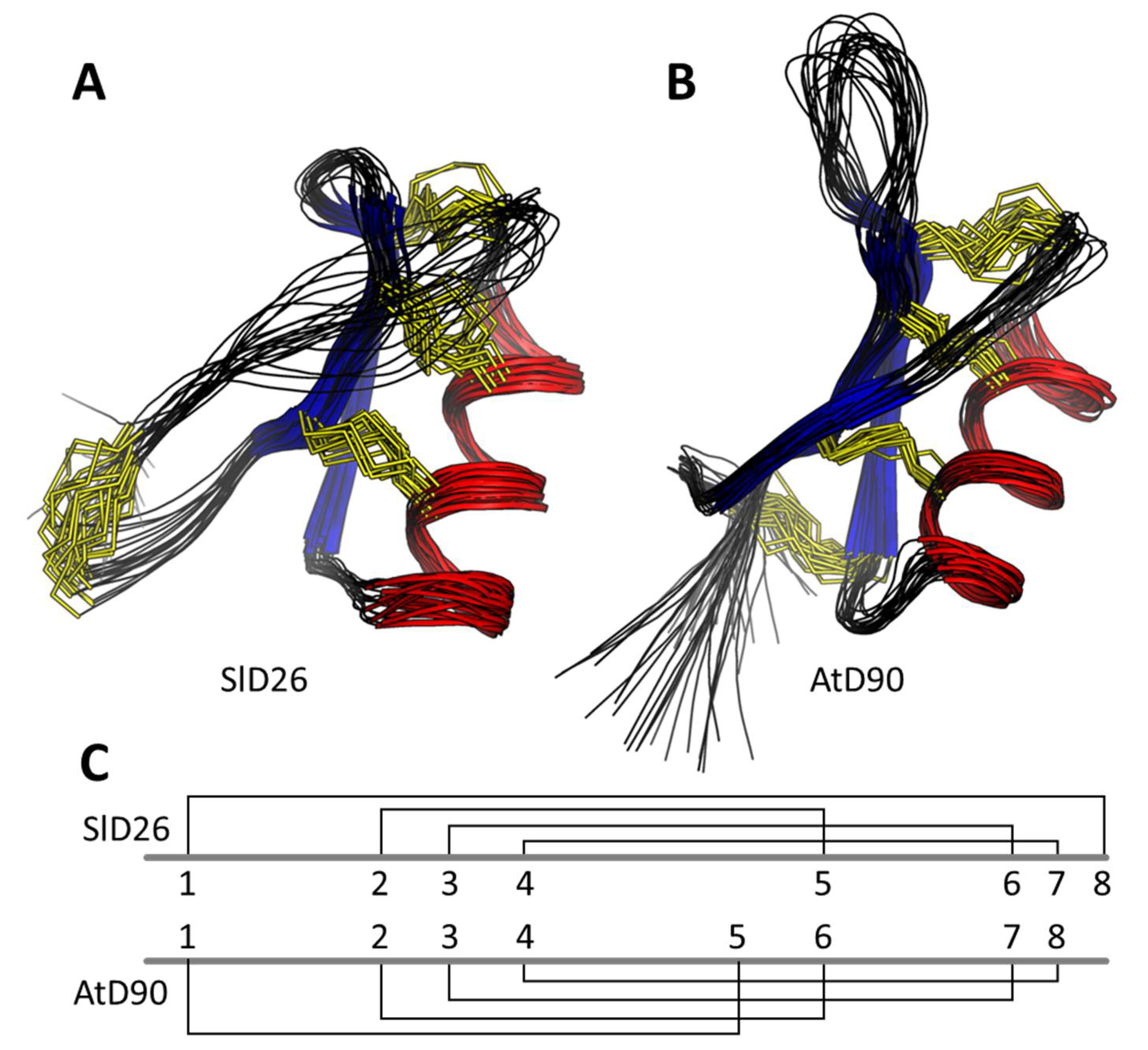
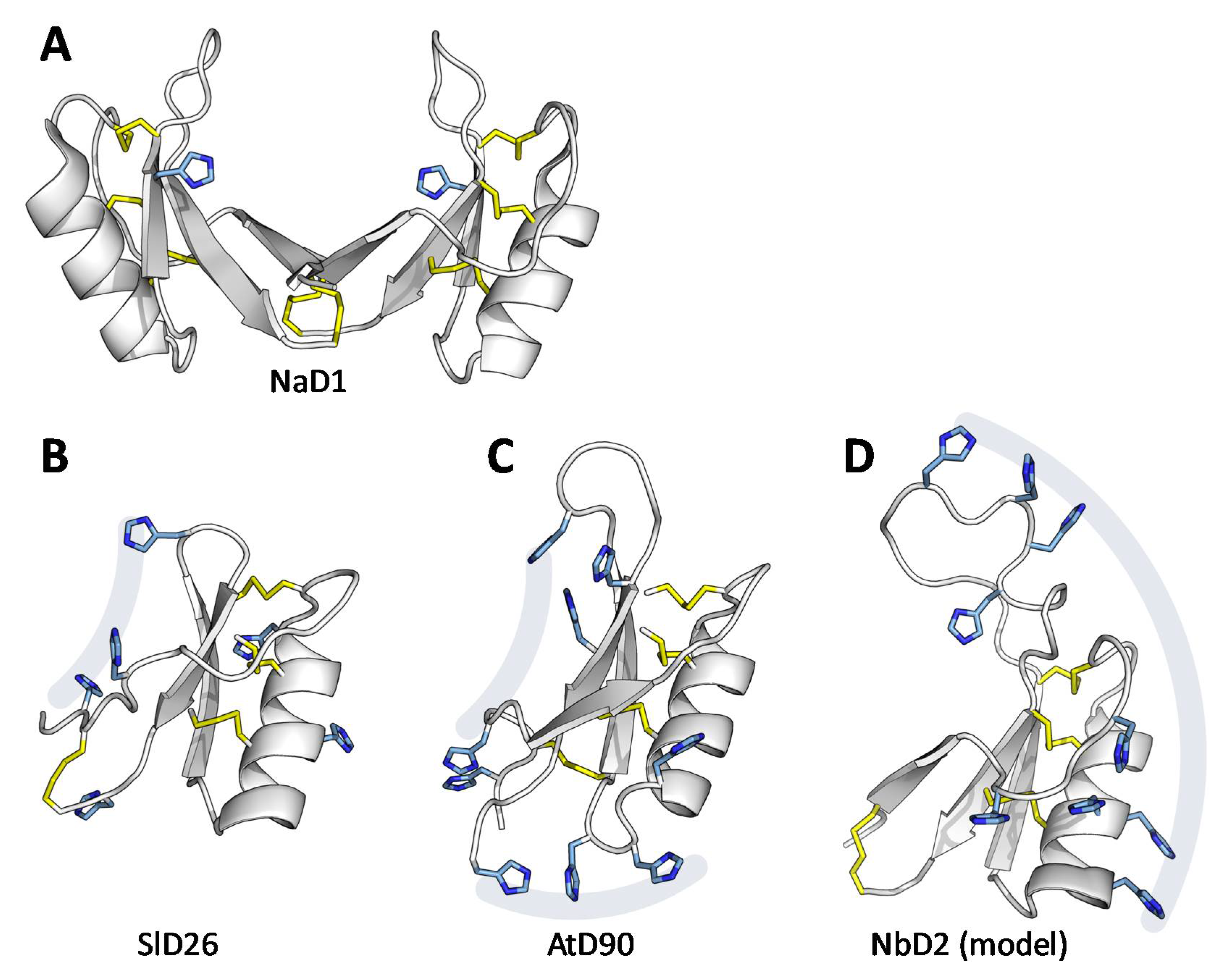
3.6. Structural Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Parisi, K.; Shafee, T.M.A.; Quimbar, P.; van der Weerden, N.L.; Bleackley, M.R.; Anderson, M.A. The Evolution, Function and Mechanisms of Action for Plant Defensins. Semin. Cell Dev. Biol. 2019, 88, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Shafee, T.M.; Lay, F.T.; Phan, T.K.; Anderson, M.A.; Hulett, M.D. Convergent Evolution of Defensin Sequence, Structure and Function. Cell. Mol. Life Sci. 2017, 4, 663–682. [Google Scholar] [CrossRef] [PubMed]
- Ishaq, N.; Bilal, M.; Iqbal, H. Medicinal Potentialities of Plant Defensins: A Review with Applied Perspectives. Medicines 2019, 6, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Weerden, N.L.; Anderson, M.A. Plant Defensins: Common Fold, Multiple Functions. Fungal Biol. Rev. 2013, 26, 121–131. [Google Scholar] [CrossRef]
- Shafee, T.M.; Robinson, A.J.; van der Weerden, N.; Anderson, M.A. Structural Homology Guided Alignment of Cysteine Rich Proteins. SpringerPlus 2016, 5, 1–7. [Google Scholar]
- Shafee, T.; Anderson, M.A. A Quantitative Map of Protein Sequence Space for the Cis-Defensin Superfamily. Bioinformatics 2018, 35, 743–752. [Google Scholar] [CrossRef]
- Luo, J.-S.; Gu, T.; Yang, Y.; Zhang, Z. A Non-Secreted Plant Defensin Atpdf2. 6 Conferred Cadmium Tolerance Via Its Chelation in Arabidopsis. Plant Mol. Biol. 2019, 100, 561–569. [Google Scholar]
- Luo, J.-S.; Huang, J.; Zeng, D.-L.; Peng, J.-S.; Zhang, G.-B.; Ma, H.-L.; Guan, Y.; Yi, H.-Y.; Fu, Y.-L.; Han, B. A Defensin-Like Protein Drives Cadmium Efflux and Allocation in Rice. Nat. Commun. 2018, 9, 645. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Luo, J.-S.; Xiao, Y.; Yao, J.; Wu, Z.; Yang, Y.; Ismail, A.M. Overexpression of a Defensin-Like Gene Cal2 Enhances Cadmium Accumulation in Plants. Front. Plant Sci. 2020, 11, 217. [Google Scholar]
- Mirouze, M.; Sels, J.; Richard, O.; Czernic, P.; Loubet, S.; Jacquier, A.; François, I.E.; Cammue, B.; Lebrun, M.; Berthomieu, P. A Putative Novel Role for Plant Defensins: A Defensin from the Zinc Hyper-Accumulating Plant, Arabidopsis Halleri, Confers Zinc Tolerance. Plant J. 2006, 47, 329–342. [Google Scholar] [CrossRef]
- Hara, M.; Fujinaga, M.; Kuboi, T. Metal Binding by Citrus Dehydrin with Histidine-Rich Domains. J. Exp. Bot. 2005, 56, 2695–2703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, I.S.; Choi, D.S.; Kim, N.H.; Kim, D.S.; Hwang, B.K. The Pepper Cysteine/Histidine-Rich Dc1 Domain Protein Cadc1 Binds Both Rna and DNA and Is Required for Plant Cell Death and Defense Response. New Phytol. 2014, 201, 518–530. [Google Scholar] [CrossRef] [PubMed]
- Park, C.J.; Park, C.B.; Hong, S.-S.; Lee, H.-S.; Lee, S.Y.; Kim, S.C. Characterization and Cdna Cloning of Two Glycine-and Histidine-Rich Antimicrobial Peptides from the Roots of Shepherd’s Purse, Capsella Bursa-Pastoris. Plant Mol. Biol. 2000, 44, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Tay, W.M.; Hanafy, A.I.; Angerhofer, A.; Ming, L.-J. A Plausible Role of Salivary Copper in Antimicrobial Activity of Histatin-5—Metal Binding and Oxidative Activity of Its Copper Complex. Bioorg. Med. Chem. Lett. 2009, 19, 6709–6712. [Google Scholar] [CrossRef]
- Grogan, J.; McKnight, C.J.; Troxler, R.F.; Oppenheim, F.G. Zinc and Copper Bind to Unique Sites of Histatin 5. FEBS Lett. 2001, 491, 76–80. [Google Scholar] [CrossRef] [Green Version]
- McCaslin, T.G.; Pagba, C.V.; Yohannan, J.; Barry, B.A. Specific Metallo-Protein Interactions and Antimicrobial Activity in Histatin-5, an Intrinsically Disordered Salivary Peptide. Sci. Rep. 2019, 9, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Sievers, F.; Higgins, D.G. Clustal Omega for Making Accurate Alignments of Many Protein Sequences. Protein Sci. 2018, 27, 135–145. [Google Scholar] [CrossRef] [Green Version]
- Schliep, K.P. Phangorn: Phylogenetic Analysis in R. Bioinformatics 2010, 27, 592–593. [Google Scholar] [CrossRef] [Green Version]
- Paradis, E.; Schliep, K. Ape 5.0: An Environment for Modern Phylogenetics and Evolutionary Analyses in R. Bioinformatics 2018, 35, 526–528. [Google Scholar] [CrossRef]
- Klepikova, A.V.; Kasianov, A.S.; Gerasimov, E.S.; Logacheva, M.D.; Penin, A.A. A High Resolution Map of the Arabidopsis Thaliana Developmental Transcriptome Based on Rna-Seq Profiling. Plant J. 2016, 88, 1058–1070. [Google Scholar] [CrossRef]
- Hayes, B.M.; Bleackley, M.R.; Anderson, M.A.; van der Weerden, N.L. The Plant Defensin Nad1 Enters the Cytoplasm of Candida Albicans Via Endocytosis. J. Fungi 2018, 4, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lay, F.T.; Mills, G.D.; Poon, I.K.; Cowieson, N.P.; Kirby, N.; Baxter, A.A.; van der Weerden, N.L.; Dogovski, C.; Perugini, M.A.; Anderson, M.A.; et al. Dimerization of Plant Defensin Nad1 Enhances Its Antifungal Activity. J. Biol. Chem. 2012, 287, 19961–19972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerenga, B.K.; McKenna, J.A.; Harvey, P.J.; Quimbar, P.; Garcia, D.; Lay, F.T.; Phan, T.K.; Veneer, P.K.; Vasa, S.; Parisi, K. Salt-Tolerant Antifungal and Antibacterial Activities of the Corn Defensin Zmd32. Front. Microbiol. 2019, 10, 795. [Google Scholar] [CrossRef] [PubMed]
- Da Costa, T.P.S.; Desbois, S.; Dogovski, C.; Gorman, M.A.; Ketaren, N.E.; Paxman, J.J.; Siddiqui, T.; Zammit, L.M.; Abbott, B.M.; Robins-Browne, R.M. Structural Determinants Defining the Allosteric Inhibition of an Essential Antibiotic Target. Structure 2016, 24, 1282–1291. [Google Scholar]
- Christensen, J.B.; Da Costa, T.S.; Faou, P.; Pearce, F.G.; Panjikar, S.; Perugini, M.A. Structure and Function of Cyanobacterial Dhdps and Dhdpr. Sci. Rep. 2016, 6, 37111. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Bax, A. Protein Backbone and Sidechain Torsion Angles Predicted from Nmr Chemical Shifts Using Artificial Neural Networks. J. Biomol. NMR 2013, 56, 227–241. [Google Scholar] [CrossRef] [Green Version]
- Brunger, A.T. Version 1.2 of the Crystallography and Nmr System. Nat. Protoc. 2007, 2, 2728. [Google Scholar] [CrossRef] [Green Version]
- Chen, V.B.; Arendall, W.B.; Headd, J.J.; Keedy, D.A.; Immormino, R.M.; Kapral, G.J.; Murray, L.W.; Richardson, J.S.; Richardson, D.C. Molprobity: All-Atom Structure Validation for Macromolecular Crystallography. Acta Crystallogr. Sect. D Biol. Crystallogr. 2010, 66, 12–21. [Google Scholar] [CrossRef] [Green Version]
- Armold, K. The Swissmodel Workspace: A Web-Based Environment for Protein Structure Homology Modilling. Bioinformatics 2006, 22, 195–201. [Google Scholar] [CrossRef] [Green Version]
- Penin, A.A.; Klepikova, A.V.; Kasianov, A.S.; Gerasimov, E.S.; Logacheva, M.D. Comparative Analysis of Developmental Transcriptome Maps of Arabidopsis Thaliana and Solanum Lycopersicum. Genes 2019, 10, 50. [Google Scholar] [CrossRef] [Green Version]
- Klepikova, A.V.; Kulakovskiy, I.V.; Kasianov, A.S.; Logacheva, M.D.; Penin, A.A. An Update to Database Trava: Organ-Specific Cold Stress Response in Arabidopsis Thaliana. BMC Plant Biol. 2019, 19, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Nakasugi, K.; Crowhurst, R.N.; Bally, J.; Wood, C.C.; Hellens, R.P.; Waterhouse, P.M. De Novo Transcriptome Sequence Assembly and Analysis of Rna Silencing Genes of Nicotiana Benthamiana. PLoS ONE 2013, 8, e59534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakasugi, K.; Crowhurst, R.; Bally, J.; Waterhouse, P. Combining Transcriptome Assemblies from Multiple De Novo Assemblers in the Allo-Tetraploid Plant Nicotiana Benthamiana. PLoS ONE 2014, 9, e91776. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, M.L.; Shafee, T.; Papenfuss, A.T.; Norton, R.S. Evolution of Cnidarian Trans-Defensins: Sequence, Structure and Exploration of Chemical Space. Proteins Struct. Funct. Bioinform. 2019, 87, 551–560. [Google Scholar] [CrossRef]
- Shafee, T.; Mitchell, M.L.; Norton, R.S. Mapping the Chemical and Sequence Space of the Shkt Superfamily. Toxicon 2019, 165, 95–102. [Google Scholar] [CrossRef]
- Lay, F.T.; Schirra, H.J.; Scanlon, M.J.; Anderson, M.A.; Craik, D.J. The Three-Dimensional Solution Structure of Nad1, a New Floral Defensin from Nicotiana Alata and Its Application to a Homology Model of the Crop Defense Protein Alfafp. J. Mol. Biol. 2003, 325, 175–188. [Google Scholar] [CrossRef]
- Develey-Rivière, M.P.; Galiana, E. Resistance to Pathogens and Host Developmental Stage: A Multifaceted Relationship within the Plant Kingdom. New Phytol. 2007, 175, 405–416. [Google Scholar] [CrossRef]
- Arif, N.; Yadav, V.; Singh, S.; Singh, S.; Ahmad, P.; Mishra, R.K.; Sharma, S.; Tripathi, D.K.; Dubey, N.; Chauhan, D.K. Influence of High and Low Levels of Plant-Beneficial Heavy Metal Ions on Plant Growth and Development. Front. Environ. Sci. 2016, 4, 69. [Google Scholar] [CrossRef]
- Jing, Y.; Shi, L.; Li, X.; Zheng, H.; Gao, J.; Wang, M.; He, L.; Zhang, W. Oxs2 Is Required for Salt Tolerance Mainly through Associating with Salt Inducible Genes, Ca1 and Araport11, in Arabidopsis. Sci. Rep. 2019, 9, 20341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Hoewyk, D.; Garifullina, G.F.; Ackley, A.R.; Abdel-Ghany, S.E.; Marcus, M.A.; Fakra, S.; Ishiyama, K.; Inoue, E.; Pilon, M.; Takahashi, H.; et al. Overexpression of Atcpnifs Enhances Selenium Tolerance and Accumulation in Arabidopsis. Plant Physiol. 2005, 139, 1518–1528. [Google Scholar] [CrossRef] [Green Version]
- Bally, J.; Nakasugi, K.; Jia, F.; Jung, H.; Ho, S.Y.; Wong, M.; Paul, C.M.; Naim, F.; Wood, C.C.; Crowhurst, R.N. The Extremophile Nicotiana Benthamiana Has Traded Viral Defence for Early Vigour. Nat. Plants 2015, 1, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Bleackley, M.R.; Payne, J.A.; Hayes, B.M.; Durek, T.; Craik, D.J.; Shafee, T.M.; Poon, I.K.; Hulett, M.D.; van der Weerden, N.L.; Anderson, M.A. Nicotiana Alata Defensin Chimeras Reveal Differences in the Mechanism of Fungal and Tumor Cell Killing and an Enhanced Antifungal Variant. Antimicrob. Agents Chemother. 2016, 60, 6302–6312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Weerden, N.L.; Bleackley, M.R.; Anderson, M.A. Properties and Mechanisms of Action of Naturally Occurring Antifungal Peptides. Cell Mol. Life Sci. 2013, 70, 3545–3570. [Google Scholar] [CrossRef] [PubMed]
- Vatamaniuk, O.K.; Mari, S.; Lu, Y.-P.; Rea, P.A. Atpcs1, a Phytochelatin Synthase from Arabidopsis: Isolation and in Vitro Reconstitution. Proc. Natl. Acad. Sci. USA 1999, 96, 7110–7115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cobbett, C.S. Phytochelatins and Their Roles in Heavy Metal Detoxification. Plant Physiol. 2000, 123, 825–832. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Cougnon, F.B.; Wanniarachchi, Y.A.; Hayden, J.A.; Nolan, E.M. Reduction of Human Defensin 5 Affords a High-Affinity Zinc-Chelating Peptide. ACS Chem. Biol. 2013, 8, 1907–1911. [Google Scholar] [CrossRef] [Green Version]
- Janssen, B.J.C.; Schirra, H.J.; Lay, F.T.; Anderson, M.A.; Craik, D.J. Structure of Petunia Hybrida Defensin 1, a Novel Plant Defensin with Five Disulfide Bonds. Biochemistry 2003, 42, 8214–8222. [Google Scholar] [CrossRef]
- Dash, T.S.; Shafee, T.; Harvey, P.J.; Zhang, C.; Peigneur, S.; Deuis, J.R.; Vetter, I.; Tytgat, J.; Anderson, M.A.; Craik, D.J. A Centipede Toxin Family Defines an Ancient Class of Csαβ Defensins. Structure 2019, 27, 315–326.e7. [Google Scholar] [CrossRef] [Green Version]
- Silverstein, K.A. Genome Organization of More Than 300 Defensin-Like Genes in Arabidopsis. Plant Physiol. 2005, 138, 600–610. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Amino Acid Sequence of Defensin Domain * | Total AA | His AA | Mass (Da) | pI |
|---|---|---|---|---|---|
| AtD90 | AHCDHFLGEAPVYPCKEKACKSVCKEHYHHACKGECEYHGREVHCHCYGDYH | 52 | 9 | 6038.8 | 6.5 |
| AtD212 | ANCDTYLGEVTVYYPCRERDCEAQCYEHYPHSCKGECEHHDHVVHHDNEEEHCHC | 55 | 9 | 6548.0 | 5.1 |
| CrD26 | ANDCDRFLGEATVSYPCRERECEAQCHEHYEHSCKGECEDHDHDHGHHDHDDHHDHEEHCHCYGH | 65 | 15 | 7721.0 | 5.2 |
| SlD26 | AKHCGKHSKSWNGKCFHKKCNHWCMEKEDAKYGSCSHGDCYCYYHC | 46 | 6 | 5422.2 | 8.7 |
| Protein | Purification Buffer System for IEX | |||||
|---|---|---|---|---|---|---|
| Citrate pH 3.0 | Acetate pH 4.0 | Phosphate pH 6.0 | Bis-Tris pH 7.0 | Buffer Selected for Purification | Yield (mg/L) | |
| AtD90 | - | Good | No Binding | NA | Acetate pH 4 | 6.3 |
| AtD212 | Good | No binding | NA | Precipitation | Citrate pH 3 | 5.0 |
| CrD26 | Good | Partial binding | NA | Precipitation | Citrate pH 3 | 5.0 |
| SlD26 | - | - | Good | - | Phosphate pH 6 | 16.0 |
| Protein | MIC F. graminearum (µg/mL) | MIC C. albicans (µg/mL) |
|---|---|---|
| AtD90 | 50 | 25 |
| AtD212 | ND | ND |
| CrD26 | ND | ND |
| SlD26 | 25 | 12.5 |
| NaD1 | 6.25 | 12.5 |
| Metal Salts | AtD90 | SlD26 | ||
|---|---|---|---|---|
| Metal Binding Affinity (Kd (µM)) | Std Error | Metal Binding Affinity (Kd (µM)) | Std Error | |
| NiCl2 | 495 | 4.3 | 44 | 3.9 |
| NaCl | no binding | - | no binding | - |
| ZnCl2 | 293 | 11.2 | 442 | 5.9 |
| MnCl2 | 230 | 3.5 | no binding | - |
| MgCl2 | no binding | - | no binding | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bleackley, M.R.; Vasa, S.; Harvey, P.J.; Shafee, T.M.A.; Kerenga, B.K.; Soares da Costa, T.P.; Craik, D.J.; Lowe, R.G.T.; Anderson, M.A. Histidine-Rich Defensins from the Solanaceae and Brasicaceae Are Antifungal and Metal Binding Proteins. J. Fungi 2020, 6, 145. https://doi.org/10.3390/jof6030145
Bleackley MR, Vasa S, Harvey PJ, Shafee TMA, Kerenga BK, Soares da Costa TP, Craik DJ, Lowe RGT, Anderson MA. Histidine-Rich Defensins from the Solanaceae and Brasicaceae Are Antifungal and Metal Binding Proteins. Journal of Fungi. 2020; 6(3):145. https://doi.org/10.3390/jof6030145
Chicago/Turabian StyleBleackley, Mark R., Shaily Vasa, Peta J. Harvey, Thomas M. A. Shafee, Bomai K. Kerenga, Tatiana P. Soares da Costa, David J. Craik, Rohan G. T. Lowe, and Marilyn A. Anderson. 2020. "Histidine-Rich Defensins from the Solanaceae and Brasicaceae Are Antifungal and Metal Binding Proteins" Journal of Fungi 6, no. 3: 145. https://doi.org/10.3390/jof6030145
APA StyleBleackley, M. R., Vasa, S., Harvey, P. J., Shafee, T. M. A., Kerenga, B. K., Soares da Costa, T. P., Craik, D. J., Lowe, R. G. T., & Anderson, M. A. (2020). Histidine-Rich Defensins from the Solanaceae and Brasicaceae Are Antifungal and Metal Binding Proteins. Journal of Fungi, 6(3), 145. https://doi.org/10.3390/jof6030145