The Penicillium chrysogenum Q176 Antimicrobial Protein PAFC Effectively Inhibits the Growth of the Opportunistic Human Pathogen Candida albicans

 , , , ,
, , , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
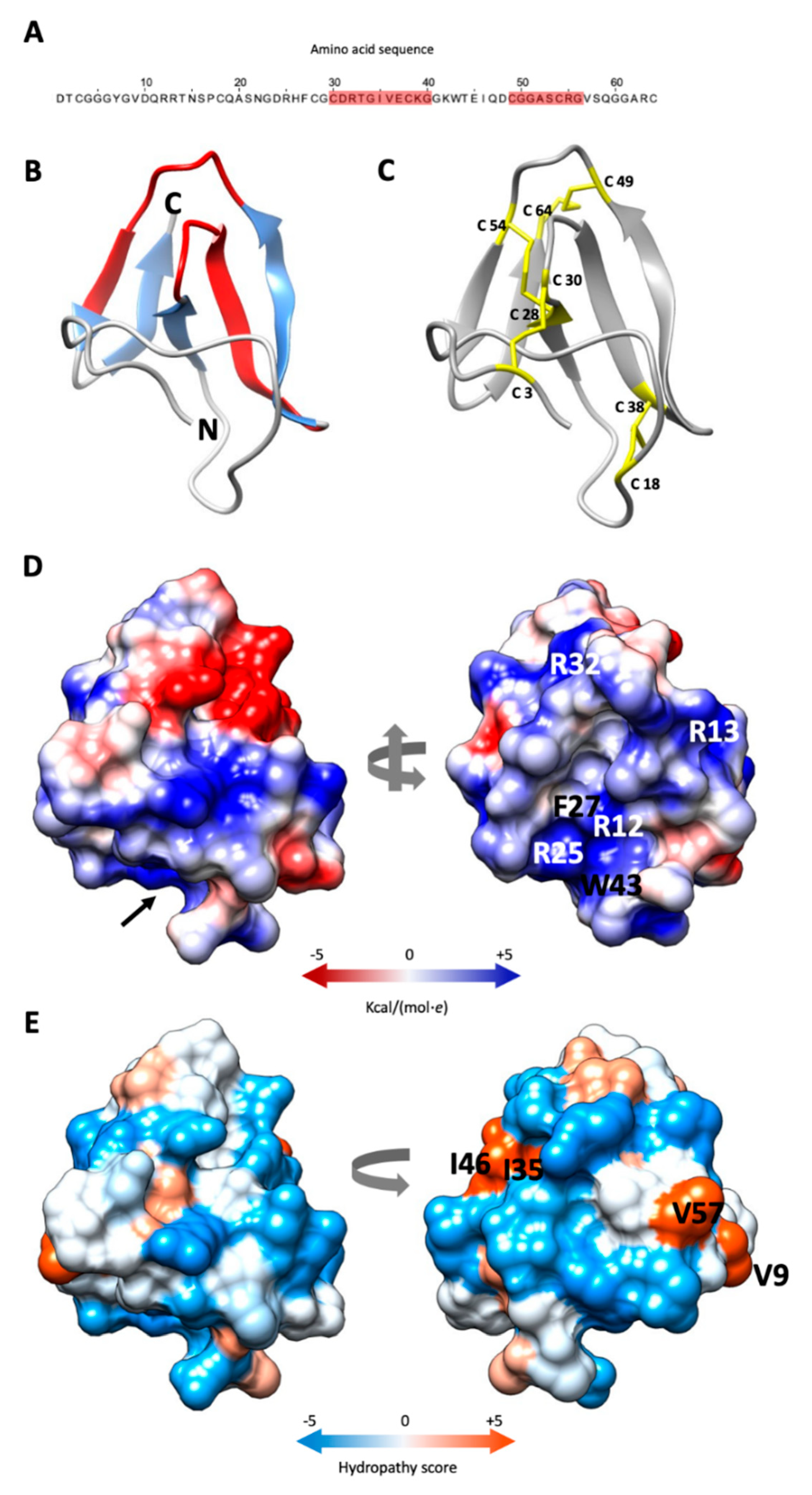
2.1. In Silico Structural Prediction of PAFC
2.2. Strains and Growth Conditions
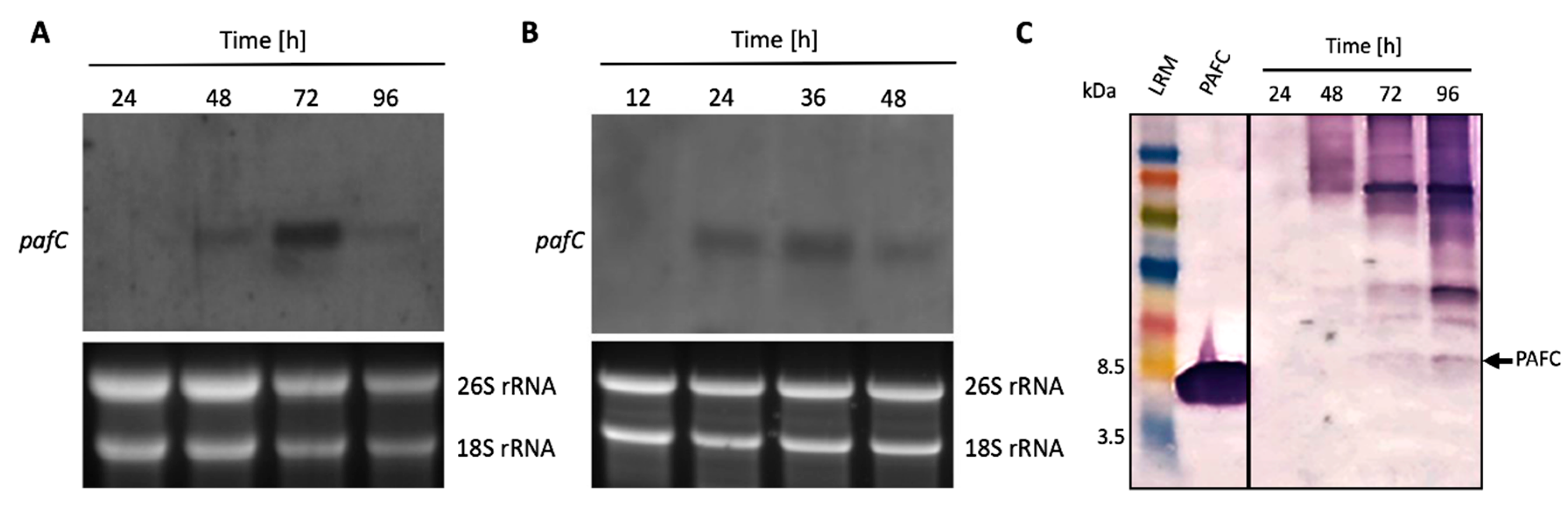
2.3. Detection of PafC mRNA
2.4. Detection of Native PAFC
2.5. Generation of Recombinant PAFC
2.6. Determination of Anti-Candida Activity
2.6.1. Broth Microdilution Assays
2.6.2. Determination of Colony Forming Units (CFU)
2.7. Scanning Electron Microscopy (SEM)
2.8. Fluorophore-Labeling of PAFC
2.9. Confocal Laser Scanning Microscopy (CLSM)
2.10. Fluorescence Activated Cell Sorting (FACS)
2.11. Hemolytic Activity
2.12. Statistical Analysis
2.13. Image Processing
3. Results
3.1. In Silico Prediction of the PAFC Structure
3.2. Expression of PAFC in P. chrysogenum
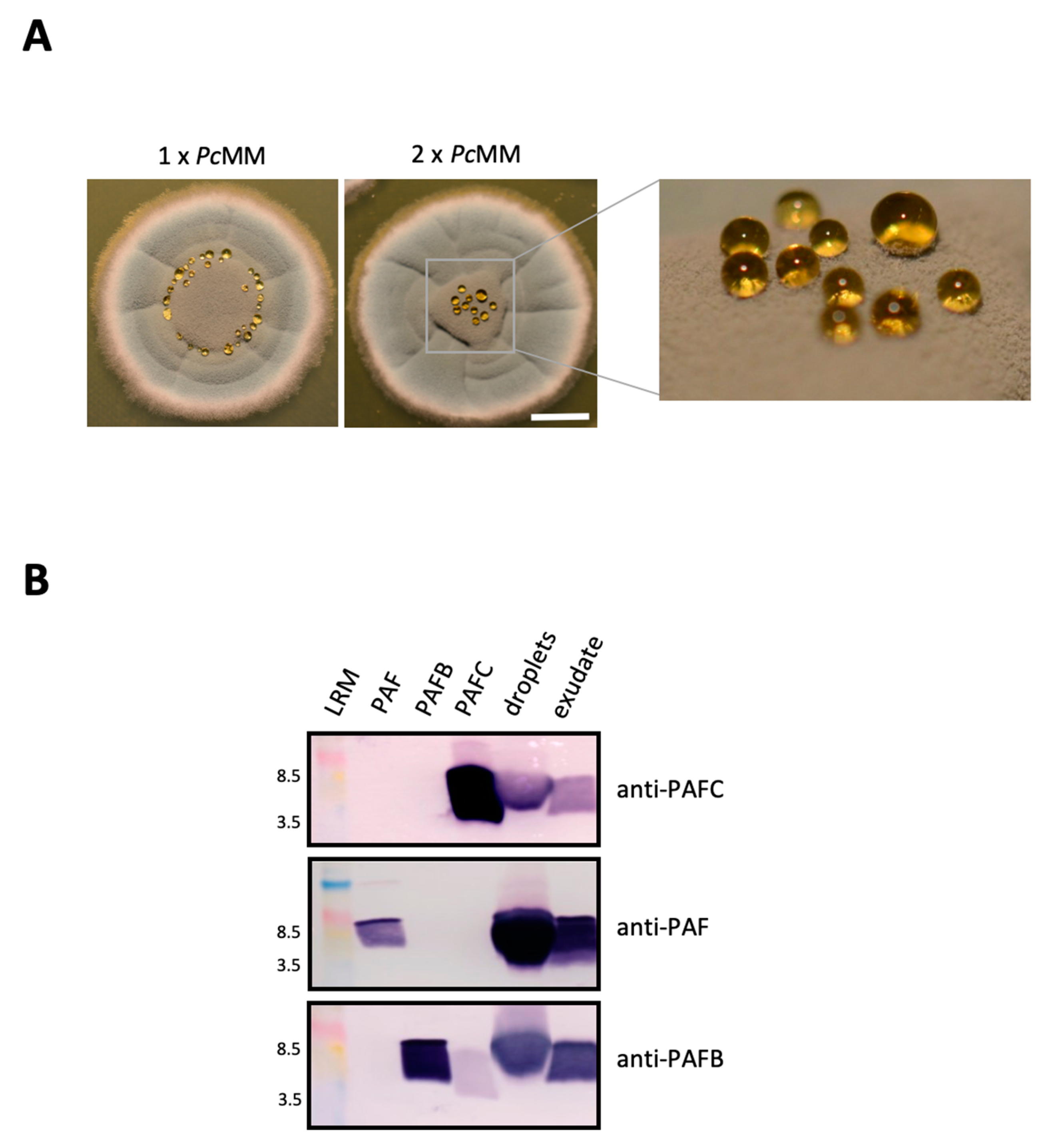
3.3. Expression of PAFC, PAF and PAFB in Fungal Exudates
3.4. Bulk Production and Purification of Recombinant PAFC
3.5. Anti-Candida Activity of PAFC in Broth Microdilution Assays
3.6. Impact of PAFC on C. albicans Biofilm
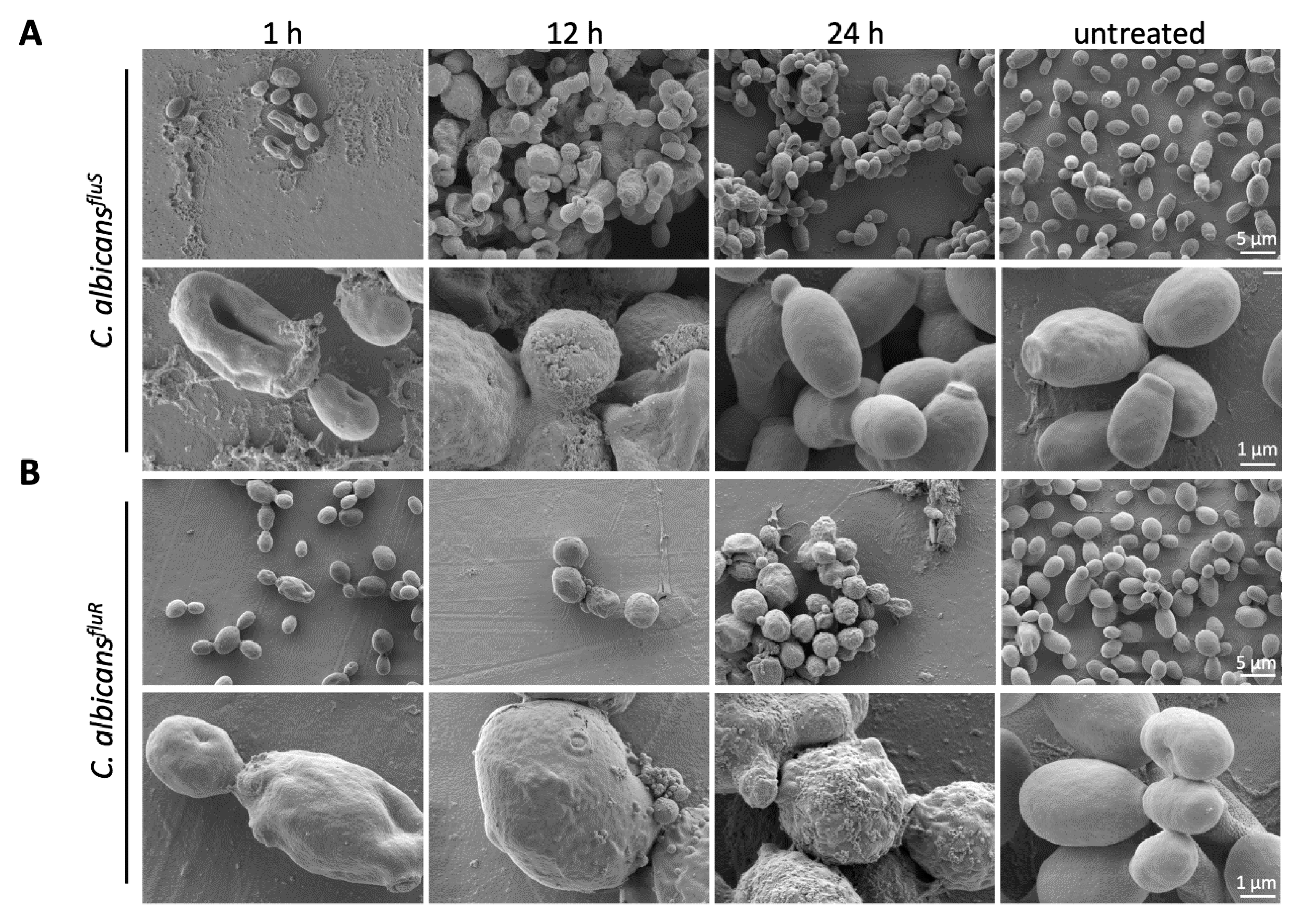
3.7. Effect of PAFC on the C. albicans Cell Morphology
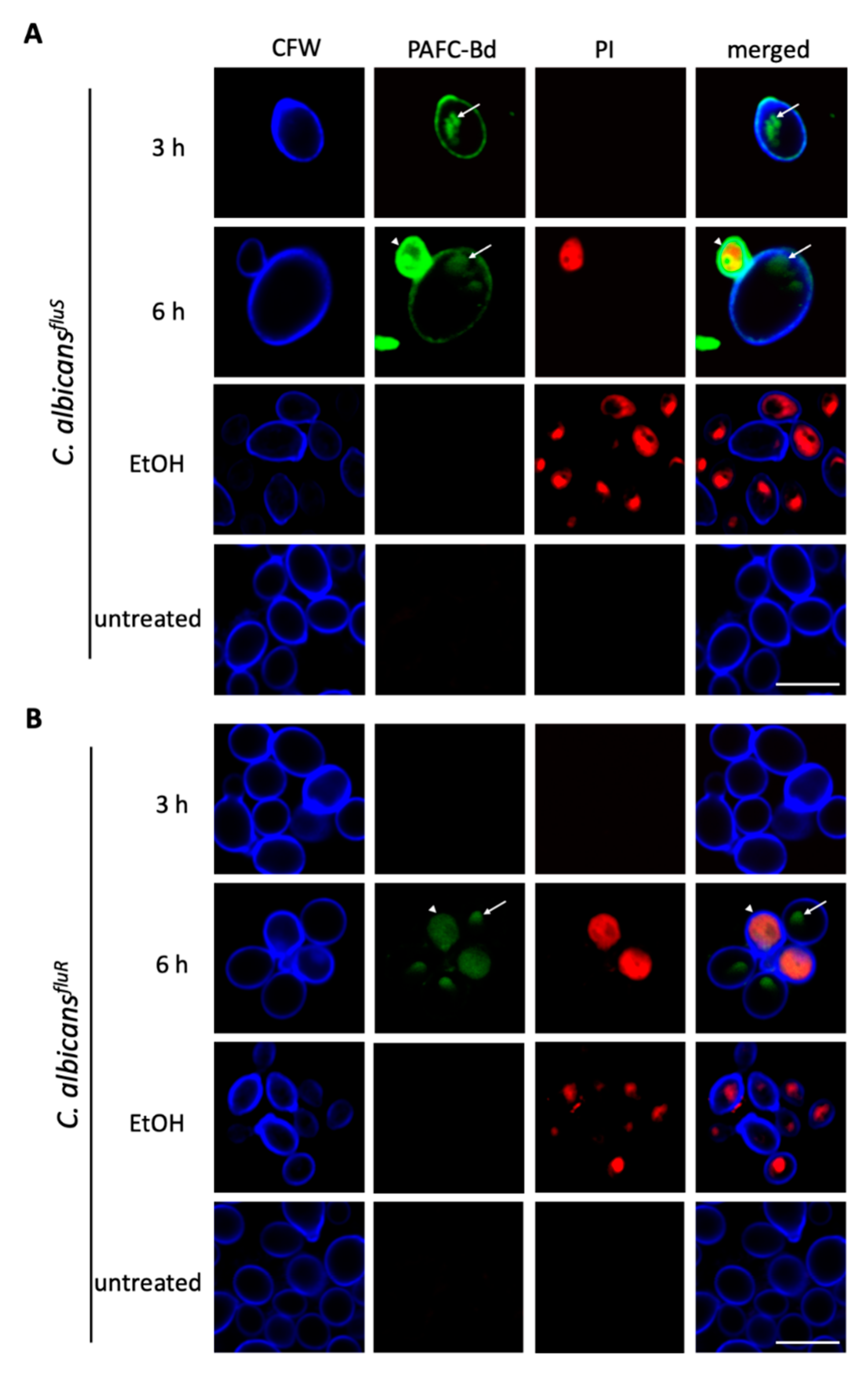
3.8. Cellular Localization of PAFC and Cell Death Induction
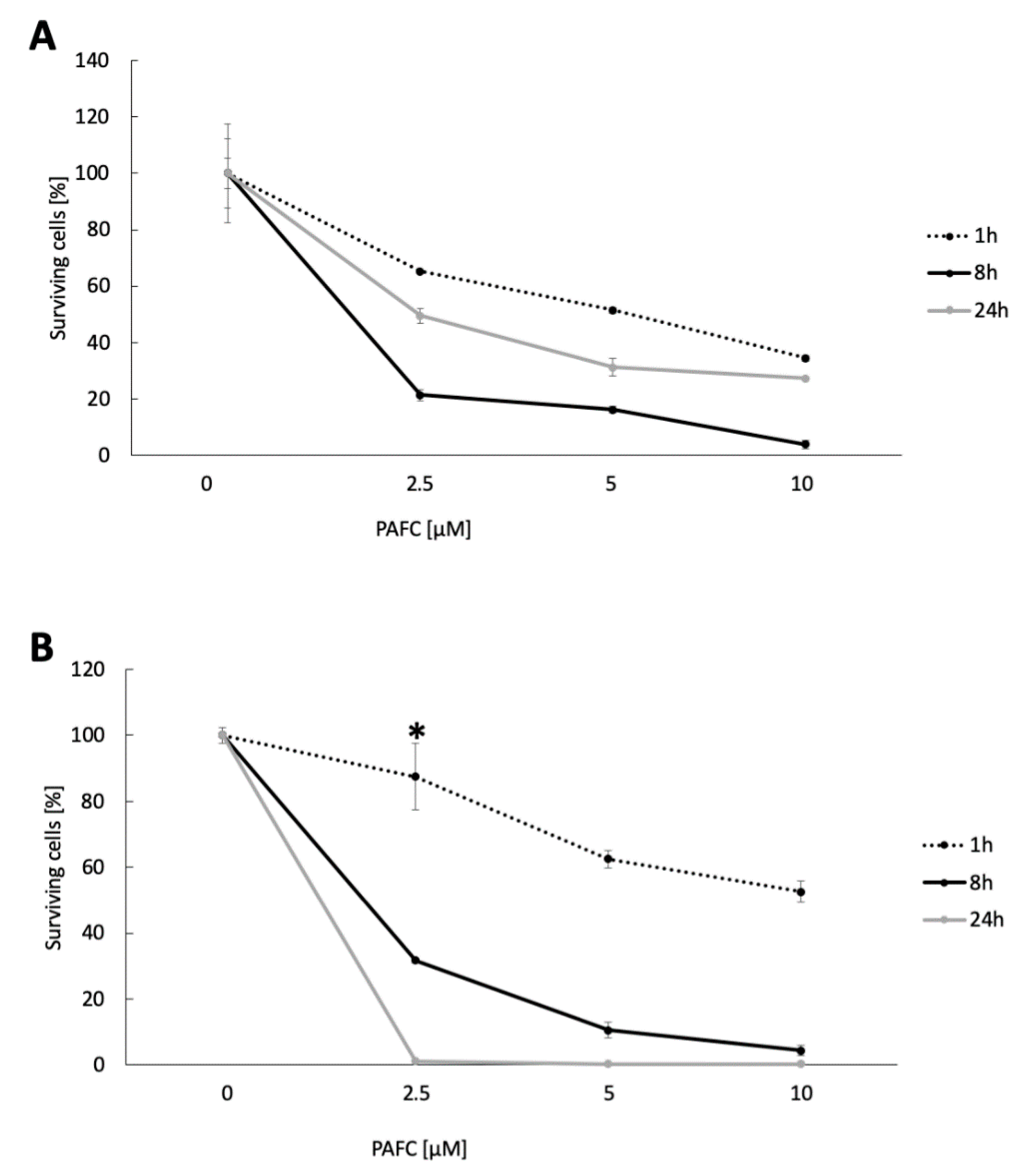
3.9. Candidacidal Efficacy of PAFC
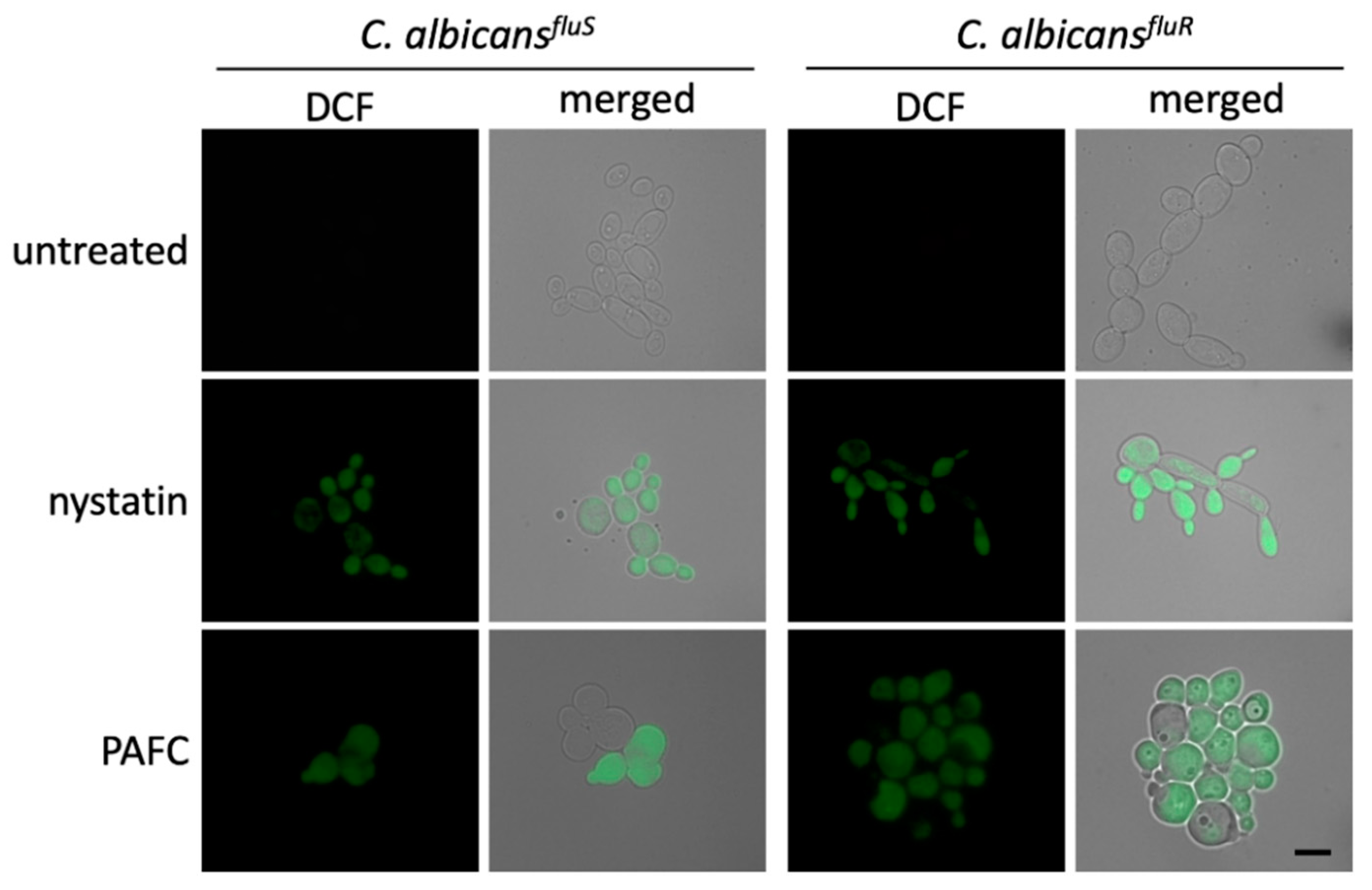
3.10. Intracellular ROS Induction by PAFC
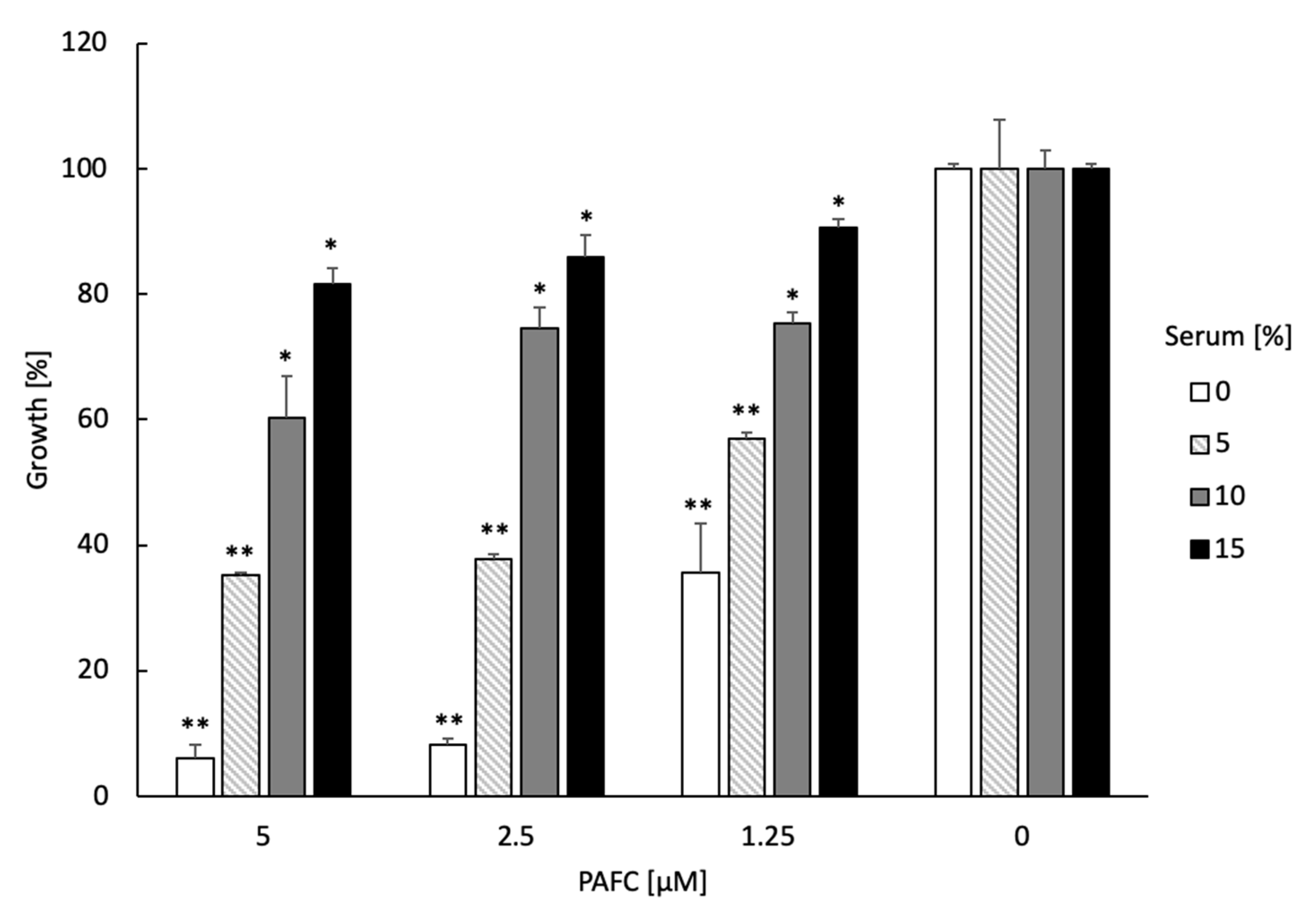
3.11. Testing of Serum–, Ion–, Thermotolerance and Hemolytic Activity of PAFC
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vallabhaneni, S.; Mody, R.K.; Walker, T.; Chiller, T. The global burden of fungal diseases. Infect. Dis. Clin. N. Am. 2016, 30, 1–11. [Google Scholar] [CrossRef]
- Perlin, D.S.; Rautemaa-Richardson, R.; Alastruey-Izquierdo, A. The global problem of antifungal resistance: Prevalence, mechanisms, and management. Lancet Infect. Dis. 2017, 17, e383–e392. [Google Scholar] [CrossRef]
- Berkow, E.L. Fluconazole resistance in Candida species: A current perspective. Infect. Drug Resist. 2017, 10, 237–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nobile, C.J.; Johnson, A.D. Candida albicans biofilms and human disease. Annu. Rev. Microbiol. 2005, 69, 71–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McManus, D. Side effects of drugs annual: A worldwide yearly survey of new data in adverse drug reactions. In Antifungal Drugs; Sidhartha, D.R., Ed.; Elsevier: Amsterdam, The Netherlands, 2017; Volume 38, pp. 243–253. [Google Scholar]
- Pristov, K.E.; Ghannoum, M.A. Resistance of Candida to azoles and echinocandins worldwide. Clin. Microbiol. Infect. 2019, 25, 792–798. [Google Scholar] [CrossRef]
- Galgóczy, L.; Yap, A.; Marx, F. Cysteine-rich antifungal proteins from filamentous fungi are promising bioactive natural compounds in anti-Candida therapy. Isr. J. Chem. 2019, 59, 360–370. [Google Scholar] [CrossRef] [Green Version]
- Huber, A.; Galgóczy, L.; Váradi, G.; Holzknecht, J.; Kakar, A.; Malanovic, N.; Leber, R.; Koch, J.; Keller, M.A.; Batta, G.; et al. Two small, cysteine-rich and cationic antifungal proteins from Penicillium chrysogenum: A comparative study of PAF and PAFB. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183246. [Google Scholar] [CrossRef]
- Huber, A.; Hajdu, D.; Bratschun-Khan, D.; Gáspári, Z.; Varbanov, M.; Philippot, S.; Fizil, A.; Czajlik, A.; Kele, Z.; Sonderegger, C.; et al. New antimicrobial potential and structural properties of PAFB: A cationic, cysteine-rich protein from Penicillium chrysogenum Q176. Sci. Rep. 2018, 8, 1751. [Google Scholar] [CrossRef]
- Chen, Z.; Ao, J.; Yang, W.; Jiao, L.; Zheng, T.; Chen, X. Purification and characterization of a novel antifungal protein secreted by Penicillium chrysogenum from an arctic sediment. Appl. Microbiol. Biotechnol. 2013, 97, 10381–10390. [Google Scholar] [CrossRef]
- Seibold, M.; Wolschann, P.; Bodevin, S.; Olsen, O. Properties of the bubble protein, a defensin and an abundant component of a fungal exudate. Peptides 2011, 32, 1989–1995. [Google Scholar] [CrossRef]
- Garrigues, S.; Gandía, M.; Castillo, L.; Coca, M.; Marx, F.; Manzanares, P.; Marcos, J.F.; Manzanares, P. Three antifungal proteins from Penicillium expansum: Different patterns of production and antifungal activity. Front Microbiol. 2018, 9, 2370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonderegger, C.; Váradi, G.; Galgóczy, L.; Kocsubé, S.; Posch, W.; Borics, A.; Dubrac, S.; Tóth, G.K.; Wilfingseder, D.; Marx, F. The evolutionary conserved γ-core motif influences the anti-Candida activity of the Penicillium chrysogenum antifungal protein PAF. Front Microbiol. 2018, 9, 1655. [Google Scholar] [CrossRef] [PubMed]
- Hoff, B.; Pöggeler, S.; Kück, U. Eighty years after its discovery, Fleming’s Penicillium strain discloses the secret of its sex. Eukaryot. Cell 2008, 7, 465–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martín, J.F. Insight into the genome of diverse Pencillium chrysogenum strains: Specific genes, cluster duplications and DNA fragment translocations. Int. J. Mol. Sci. 2020, 21, 3936. [Google Scholar] [CrossRef] [PubMed]
- Kovács, R.; Holzknecht, J.; Hargitai, Z.; Papp, C.; Farkas, A.; Borics, A.; Tóth, L.; Váradi, G.; Tóth, G.; Kovács, I.; et al. In vivo applicability of Neosartorya fischeri antifungal protein 2 (NFAP2) in treatment of vulvovaginal candidiasis. Antimicrob. Agents Chemother. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Yan, R.; Roy, A.; Xu, D.; Poisson, J.; Zhang, Y. The I-TASSER suite: Protein structure and function prediction. Nat. Methods 2015, 12, 7–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lovell, S.C.; Davis, I.W.; Arendall, W.B., 3rd; de Bakker, P.I.; Word, J.M.; Prisant, M.G.; Richardson, J.S.; Richardson, D.C. Structure validation by Calpha geometry: Phi, psi and Cbeta deviation. Proteins 2003, 50, 437–450. [Google Scholar] [CrossRef] [PubMed]
- Ceroni, A.; Passerini, A.; Vullo, A.; Frasconi, P. DISULFIND: A disulfide bonding state and cysteine connectivity prediction server. Nucleic. Acids Res. 2006, 34, W177–W181. [Google Scholar] [CrossRef] [Green Version]
- Leitner, A.; Walzthoeni, T.; Kahraman, A.; Herzog, F.; Rinner, O.; Beck, M.; Aebersold, R. Probing native protein structures by chemical cross-linking, mass spectrometry, and bioinformatics. Mol. Cell. Proteom. 2010, 9, 1634–1649. [Google Scholar] [CrossRef] [Green Version]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 13, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.; Clamp, M.; Barton, G.J. Jalview Version 2-a multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 5, 1189–1191. [Google Scholar] [CrossRef] [Green Version]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Hegedüs, N.; Sigl, C.; Zadra, I.; Pócsi, I.; Marx, F. The paf gene product modulates asexual development in Penicillium chrysogenum. J. Basic Microbiol. 2011, 51, 253–262. [Google Scholar] [CrossRef] [Green Version]
- Oberparleiter, C.; Kaiserer, L.; Haas, H.; Ladurner, P.; Andratsch, M.; Marx, F. Active internalization of the Penicillium chrysogenum antifungal protein PAF in sensitive Aspergilli. Antimicrob. Agents Chemother. 2003, 47, 3598–3601. [Google Scholar] [CrossRef] [Green Version]
- Sonderegger, C.; Galgóczy, L.; Garrigues, S.; Fizil, Á.; Borics, A.; Manzanares, P.; Hegedüs, N.; Huber, A.; Marcos, J.F.; Batta, G.; et al. A Penicillium chrysogenum-based expression system for the production of small, cysteine-rich antifungal proteins for structural and functional analyses. Microb. Cell Fact. 2016, 15, 192. [Google Scholar] [CrossRef] [Green Version]
- Sonderegger, C.; Fizil, Á.; Burtscher, L.; Hajdu, D.; Muñoz, A.; Gáspári, Z.; Read., N.D.; Batta, G.; Marx, F. D19S mutation of the cationic, cysteine-rich protein PAF: Novel insights into its structural dynamics, thermal unfolding and antifungal function. PLoS ONE 2017, 12, e0169920. [Google Scholar] [CrossRef] [Green Version]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Yount, N.Y.; Yeaman, M.R. Multidimensional signatures in antimicrobial peptides. Proc. Natl. Acad. Sci. USA 2004, 101, 7363–7368. [Google Scholar] [CrossRef] [Green Version]
- Olsen, J.G.; Flensburg, C.; Olsen, O.; Bricogne, G.; Henriksen, A. Solving the structure of the bubble protein using the anomalous sulfur signal from single-crystal in-house Cu Kalpha diffraction data only. Acta Cryst. D Biol Cryst. 2004, 60, 250–255. [Google Scholar] [CrossRef]
- Kyte, J.; Doolittle, R.F. A simple method for displaying the hydropathic character of a protein. J. Mol. Biol. 1982, 157, 105–132. [Google Scholar] [CrossRef] [Green Version]
- Cantoral, J.M.; Díez, B.; Barredo, J.; Alvarez, E.; Martín, J.F. High–frequency transformation of Penicillium chrysogenum. Nat. Biotechnol. 1987, 5, 494–497. [Google Scholar] [CrossRef]
- Kolar, M.; Punt, P.J.; van den Hondel, C.A.; Schwab, H. Transformation of Penicillium chrysogenum using dominant selection markers and expression of an Escherichia coli lacZ fusion gene. Gene 1988, 62, 127–134. [Google Scholar] [CrossRef]
- Polke, M.; Hube, B.; Jacobsen, I.D. Candida survival strategies. Adv. Appl. Microbiol. 2015, 91, 139–235. [Google Scholar] [CrossRef] [PubMed]
- Aerts, A.M.; François, I.E.; Meert, E.M.; Li, Q.T.; Cammue, B.P.; Thevissen, K. The antifungal activity of RsAFP2, a plant defensin from Raphanus sativus, involves the induction of reactive oxygen species in Candida albicans. J. Mol. Microbiol. Biotechnol. 2007, 13, 243–247. [Google Scholar] [CrossRef]
- Muñoz, A.; Marcos, J.F.; Read, N.D. Concentration-dependent mechanisms of cell penetration and killing by the de novo designed antifungal hexapeptide PAF26. Mol. Microbiol. 2012, 85, 89–106. [Google Scholar] [CrossRef]
- Marx, F.; Binder, U.; Leiter, É.; Pócsi, I. The Penicillium chrysogenum antifungal protein PAF, a promising tool for the development of new antifungal therapies and fungal cell biology studies. Cell. Mol. Life Sci. 2008, 65, 445–454. [Google Scholar] [CrossRef]
- Batta, G.; Barna, T.; Gáspári, Z.; Sándor, S.; Kövér, K.E.; Binder, U.; Sarg, B.; Kaiserer, L.; Chillar, A.K.; Eigentler, A.; et al. Functional aspects of the solution structure and dynamics of PAF—A highly-stable antifungal protein from Penicillium chrysogenum. FEBS J. 2009, 276, 2875–4658. [Google Scholar] [CrossRef] [Green Version]
- Laskowskie, R.A.; Rullmann, J.A.C.; MacArthur, M.W.; Kaptein, R.; Thornton, J.M. AQUA and PROCHECK-NMR: Programs for checking the quality of protein structures solved by NMR. J. Biomol. NMR 1996, 8, 477–486. [Google Scholar] [CrossRef]
- Breustedt, D.A.; Korndo, I.P.; Redl, B.; Skerra, A. The 1.8-Å crystal structure of human tear lipocalin reveals an extended branched cavity with capacity for multiple ligands. J. Biol. Chem. 2005, 280, 484–493. [Google Scholar] [CrossRef] [Green Version]
- Hegedüs, N.; Marx, F. Antifungal proteins: More than antimicrobials? Fungal. Biol. Rev. 2013, 26, 132–145. [Google Scholar] [CrossRef]
- Paege, N.; Jung, S.; Schäpe, P.; Müller-Hagen, D.; Ouedraogo, J.P.; Heiderich, C.; Jedamzick, J.; Nitsche, B.M.; van den Hondel, C.A.; Ram, A.F.; et al. A transcriptome meta-analysis proposes novel biological roles for the antifungal protein AnAFP in Aspergillus niger. PLoS ONE 2016, 11, e0165755. [Google Scholar] [CrossRef] [PubMed]
- McPhee, W.J.; Cologelo, N. Fungal exudates. I. Characteristics of hyphal exudates in Fursarium culmorum. Can. J. Bot. 1977, 55, 358–365. [Google Scholar] [CrossRef]
- Garrigues, S.; Gandía, M.; Popa, C.; Borics, A.; Marx, F.; Coca, M.; Marcos, J.F.; Manzanares, P. Efficient production and characterization of the novel and highly active antifungal protein AfpB from Penicillium digitatum. Sci. Rep. 2017, 7, 14663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fizil, Á.; Sonderegger, C.; Czajlik, A.; Fekete, A.; Komáromi, I.; Hajdu, D.; Marx, F.; Batta, G. Calcium binding of the antifungal protein PAF: Structure, dynamics and function aspects by NMR and MD simulations. PLoS ONE 2018, 13, e0204825. [Google Scholar] [CrossRef]
- Heredero, M.; Garrigues, S.; Gandía, M.; Marcos, J.F.; Manzanares, P. Rational design and biotechnological production of novel AfpB-PAF26 chimeric antifungal proteins. Microorganisms 2018, 6, 106. [Google Scholar] [CrossRef] [Green Version]
- Tóth, L.; Váradi, G.; Borics, A.; Batta, G.; Kele, Z.; Vendrinszky, Á.; Tóth, R.; Ficze, H.; Vágvölgyi, C.; Marx, F.; et al. Anti-candidal activity and functional mapping of recombinant and synthetic Neosartorya fischeri antifungal protein 2 ( NFAP2 ). Front Microbiol. 2018, 9, 393. [Google Scholar] [CrossRef] [Green Version]
- Hajdu, D.; Huber, A.; Czajlik, A.; Tóth, L.; Kele, Z.; Kocsubé, S.; Fizil, A.; Marx, F.; Galgóczy, L.; Batta, G. Solution structure and novel insights into phylogeny and mode of action of the Neosartorya (Aspergillus) fischeri antifungal protein (NFAP). Int. J. Biol. Macromol. 2019, 129, 511–522. [Google Scholar] [CrossRef]
- Ouedraogo, J.P.; Hagen, S.; Spielvogel, A.; Engelhardt, S.; Meyer, V. Survival strategies of yeast and filamentous fungi against the antifungal protein AFP. J. Biol. Chem. 2011, 286, 13859–13868. [Google Scholar] [CrossRef] [Green Version]
- Robbins, N.; Caplan, T.; Cowen, L.E. Molecular evolution of antifungal drug resistance. Annu. Rev. Microbiol. 2017, 71, 753–775. [Google Scholar] [CrossRef] [Green Version]
- Muñoz, A.; Harries, E.; Contreras-Valenzuela, A.; Carmona, L.; Read, N.D.; Marcos, J.F. Two functional motifs define the interaction, internalization and toxicity of the cell-penetrating antifungal peptide PAF26 on fungal cells. PLoS ONE 2013, 8, e54813. [Google Scholar] [CrossRef]
- Phillips, A.J.; Sudbery, I.; Ramsdale, M. Apoptosis induced by environmental stresses and amphotericin B in Candida albicans. Proc. Natl. Acad. Sci. USA 2003, 100, 14327–14332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaiserer, L.; Oberparleiter, C.; Weiler-Görz, R.; Burgstaller, W.; Leiter, E.; Marx, F. Characterization of the Penicillium chrysogenum antifungal protein PAF. Arch. Microbiol. 2003, 180, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Vriens, K.; Cammue, B.P.A.; Thevissen, K. Antifungal plant defensins: Mechanisms of action and production. Molecules 2014, 19, 12280–12303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parisi, K.; Shafee, T.M.A.; Quimbar, P.; van der Weerden, N.L.; Bleackley, M.R.; Anderson, M.A. The evolution, function and mechanisms of action for plant defensins. Semin. Cell Dev. Biol. 2019, 88, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Chen, C.; Zhang, B.; Jing, H.; Wang, Z.; Wu, C.; Hao, P.; Kuang, Y.; Yang, M. The chromogranin A-derived antifungal peptide CGA-N9 induces apoptosis in Candida tropicalis. Biochem. J. 2019, 476, 3069–3080. [Google Scholar] [CrossRef] [Green Version]
- Bourdichon, F.; Casaregola, S.; Farrokh, C.; Frisvad, J.C.; Gerds, M.L.; Hammes, W.P.; Harnett, J.; Huys, G.; Laulund, S.; Ouwehand, A.; et al. Food fermentations: Microoganisms with technological benefical use. Int. J. Food Microbiol. 2012, 154, 87–97. [Google Scholar] [CrossRef]
- Galgóczy, L.; Borics, A.; Virágh, M.; Ficze, H.; Váradi, G.; Kele, Z.; Marx, F. Structural Determinants of Neosartorya fischeri antifungal protein (NFAP) for folding, stability and antifungal activity. Sci. Rep. 2017, 7, 1963. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microorganisms | PAFC | Fluconazole | Amphotericin B | Nystatin |
|---|---|---|---|---|
| Candida albicansfluS | 2.50 (16.50) | 6.37 (1.95) | 0.02 (0.02) | 2.11 (1.95) |
| Candida albicansfluR | 2.50 (16.50) | 408.50 (>125) | 1.05 (0.97) | 0.53 (0.49) |
| Candida glabrata | 0.15 (0.99) | 12.78 (3.91) | 0.03 (0.03) | 0.53 (0.49) |
| Candida guilliermondii | 3.12 (20.59) | 1.60 (0.49) | 0.06 (0.06) | 1.06 (0.98) |
| Candida krusei | 2.50 (16.50) | 51.08 (15.63) | 0.06 (0.06) | 1.06 (0.98) |
| Candida parapsilosis | 0.04 (0.26) | 12.78 (3.91) | 8.45 (7.81) | 1.06 (0.98) |
| Strain | Untreated | PAFC | Amphotericin B 10 µg mL−1 | |
|---|---|---|---|---|
| 1 × IC90 | 10 × IC90 | |||
| Candida albicansfluS | 100 ± 10.8 | 83.4 ± 3.0 | 47.0 ± 3.8 * | 0 ± 0 ** |
| Candida albicansfluR | 100 ± 5.8 | 48.1 ± 2.2 ** | 28.1 ± 4.5 ** | 0 ± 0 ** |
| Strain | Untreated | 1 h | 8 h | 24 h | EtOH |
|---|---|---|---|---|---|
| C. albicansfluS | 0.10 ± 0.1 | 38.10 ± 0.8 ** | 60.95 ± 12.0 ** | 0.89 ± 0.3 | 99.17 ± 3.1 ** |
| C. albicansfluR | 0.09 ± 0.04 | 17.37 ± 2.3 ** | 53.30 ± 5.6 ** | 77.95 ± 11.7 ** | 99.17 ± 5.4 ** |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Holzknecht, J.; Kühbacher, A.; Papp, C.; Farkas, A.; Váradi, G.; Marcos, J.F.; Manzanares, P.; Tóth, G.K.; Galgóczy, L.; Marx, F. The Penicillium chrysogenum Q176 Antimicrobial Protein PAFC Effectively Inhibits the Growth of the Opportunistic Human Pathogen Candida albicans. J. Fungi 2020, 6, 141. https://doi.org/10.3390/jof6030141
Holzknecht J, Kühbacher A, Papp C, Farkas A, Váradi G, Marcos JF, Manzanares P, Tóth GK, Galgóczy L, Marx F. The Penicillium chrysogenum Q176 Antimicrobial Protein PAFC Effectively Inhibits the Growth of the Opportunistic Human Pathogen Candida albicans. Journal of Fungi. 2020; 6(3):141. https://doi.org/10.3390/jof6030141
Chicago/Turabian StyleHolzknecht, Jeanett, Alexander Kühbacher, Csaba Papp, Attila Farkas, Györgyi Váradi, Jose F. Marcos, Paloma Manzanares, Gábor K. Tóth, László Galgóczy, and Florentine Marx. 2020. "The Penicillium chrysogenum Q176 Antimicrobial Protein PAFC Effectively Inhibits the Growth of the Opportunistic Human Pathogen Candida albicans" Journal of Fungi 6, no. 3: 141. https://doi.org/10.3390/jof6030141
APA StyleHolzknecht, J., Kühbacher, A., Papp, C., Farkas, A., Váradi, G., Marcos, J. F., Manzanares, P., Tóth, G. K., Galgóczy, L., & Marx, F. (2020). The Penicillium chrysogenum Q176 Antimicrobial Protein PAFC Effectively Inhibits the Growth of the Opportunistic Human Pathogen Candida albicans. Journal of Fungi, 6(3), 141. https://doi.org/10.3390/jof6030141