The Therapy of Pulmonary Fibrosis in Paracoccidioidomycosis: What Are the New Experimental Approaches?


Abstract
:1. Introduction
2. Paracoccidioidomycosis and Development of Pulmonary Fibrosis
3. Therapeutic Approaches for Pulmonary Fibrosis in Paracoccidioidomycosis
3.1. Pharmacological Therapy
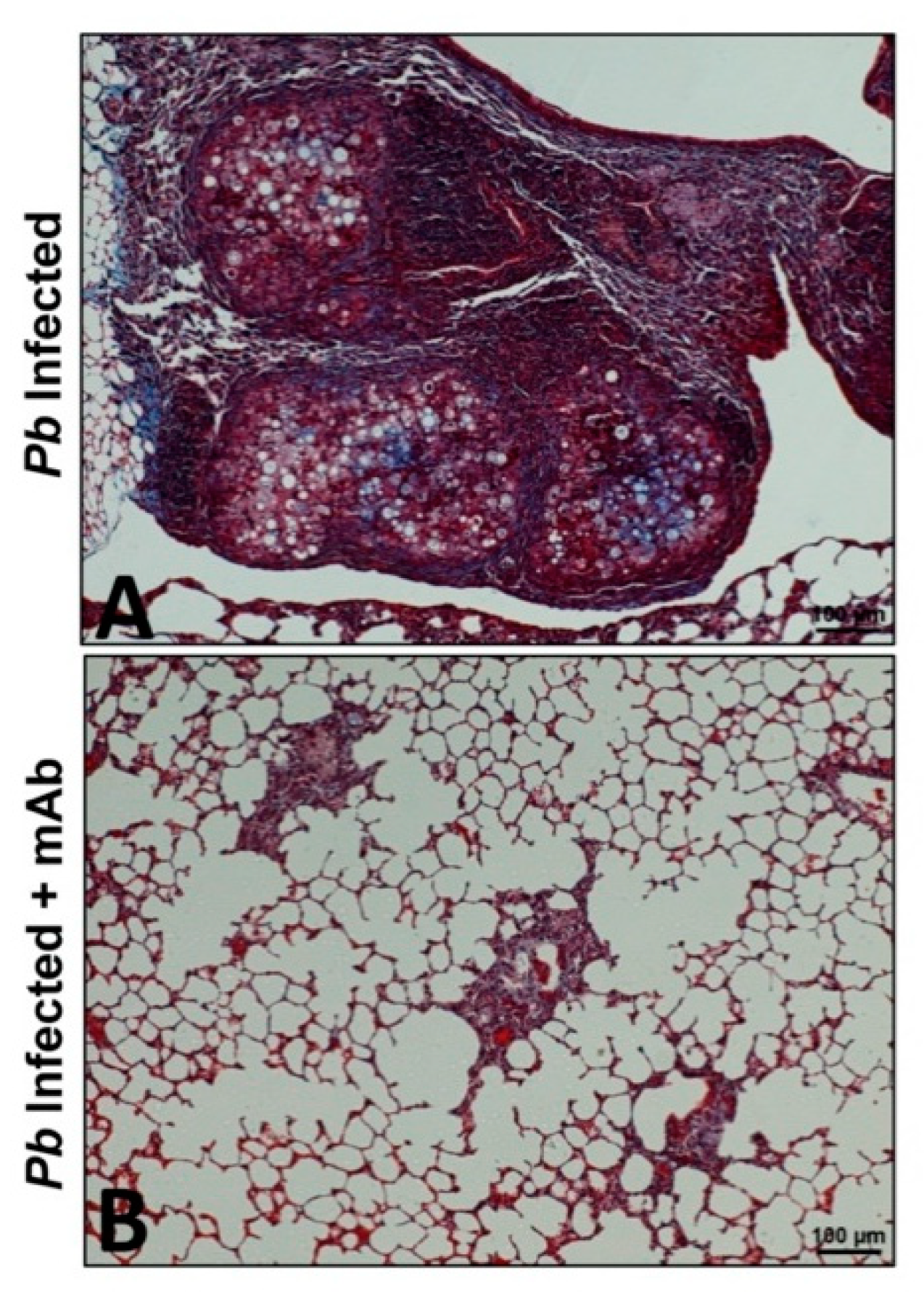
3.2. Immunotherapy or Antibody-Based Therapy
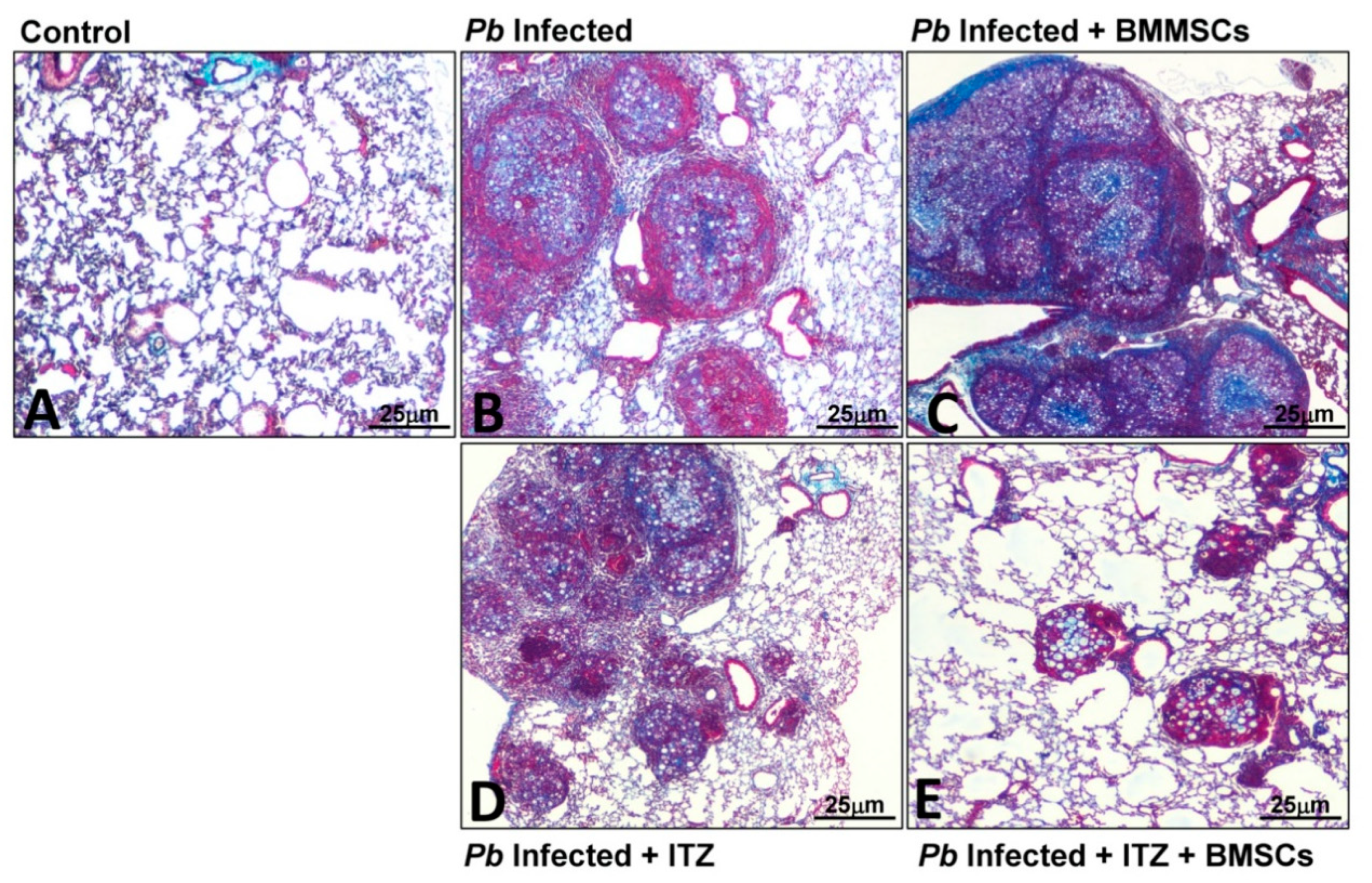
3.3. Cellular Therapy
3.4. Vaccination
4. Conclusions and Future Directions
Funding
Conflicts of Interest
References
- Wynn, T.A.; Ramalingam, T.R. Mechanisms of fibrosis: Therapeutic translation for fibrotic disease. Nat. Med. 2012, 18, 1028–1040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nalysnyk, L.; Cid-Ruzafa, J.; Rotella, P.; Esser, D. Incidence and prevalence of idiopathic pulmonary fibrosis: A review of the literature. Eur. Respir. Rev. 2012, 21, 355–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, T.E., Jr.; Pardo, A.; Selman, M. Idiopathic pulmonary fibrosis. Lancet 2011, 378, 1949–1961. [Google Scholar] [CrossRef]
- Chioma, O.S.; Drake, W.P. Role of Microbial Agents in Pulmonary Fibrosis. Yale J. Biol. Med. 2017, 90, 219–227. [Google Scholar]
- Restrepo, A.; Tobón, A.M.; González, A. Paracoccidioidomycosis, Chapter 267. In Principles and Practice of Infectious Diseases, 9th ed.; Mandell, G.L., Douglas, Bennett’s, J.E., Eds.; Elsevier: Philadelphia, PA, USA, 2019; pp. 3211–3221. [Google Scholar]
- Mendes, R.P.; de Cavalcante, R.S.; Marques, S.A.; Marques, M.E.A.; Venturini, J.; Sylvestre, T.F.; Paniago, A.M.M.; Pereira, A.C.; da Silva, J.F.; Fabro, A.T.; et al. Paracoccidioidomycosis: Current perspectives from Brazil. Open Microbiol. J. 2017, 11, 224–282. [Google Scholar] [CrossRef] [Green Version]
- Tobón, A.M.; Agudelo, C.A.; Osorio, M.L.; Álvarez, D.L.; Arango, M.; Cano, L.E.; Restrepo, A. Residual pulmonary abnormalities in adult patients with chronic paracoccidioidomycosis: Prolonged follow-up after itraconazole therapy. Clin. Infect. Dis. 2003, 37, 898–904. [Google Scholar] [CrossRef] [Green Version]
- Naranjo, T.W.; Lopera, D.E.; Diaz-Granados, L.R.; Duque, J.J.; Restrepo, A.M.; Cano, L.E. Combined itraconazole-pentoxifylline treatment promptly reduces lung fibrosis induced by chronic pulmonary paracoccidioidomycosis in mice. Pulm. Pharmacol. Ther. 2011, 24, 81–91. [Google Scholar] [CrossRef]
- Lopera, D.E.; Naranjo, T.W.; Hidalgo, J.M.; Echeverri, L.; Patiño, J.H.; Moreno, Á.R.; Lenzi, H.L.; Cano, L.E. Pentoxifylline immunomodulation in the treatment of experimental chronic pulmonary paracoccidioidomycosis. Fibrogenes. Tissue Repair 2015, 8, 10. [Google Scholar] [CrossRef] [Green Version]
- Puerta-Arias, J.D.; Pino-Tamayo, P.A.; Arango, J.C.; González, Á. Depletion of Neutrophils Promotes the Resolution of Pulmonary Inflammation and Fibrosis in Mice Infected with Paracoccidioides brasiliensis. PLoS ONE 2016, 11, e0163985. [Google Scholar] [CrossRef] [Green Version]
- Puerta-Arias, J.D.; Pino-Tamayo, P.A.; Arango, J.C.; Salazar-Peláez, L.M.; González, A. Itraconazole in combination with neutrophil depletion reduces the expression of genes related to pulmonary fibrosis in an experimental model of paracoccidioidomycosis. Med. Mycol. 2018, 56, 579–590. [Google Scholar] [CrossRef] [Green Version]
- Finato, A.C.; Almeida, D.F.; Dos Santos, A.R.; Nascimento, D.C.; Cavalcante, R.S.; Mendes, R.P.; Soares, C.T.; Paniago, A.M.M.; Venturini, J. Evaluation of antifibrotic and antifungal combined therapies in experimental pulmonary paracoccidioidomycosis. Med. Mycol. 2020, 58, 667–678. [Google Scholar] [CrossRef] [PubMed]
- Martinez, R. Epidemiology of paracoccidioidomycosis. Rev. Inst. Med. Trop. Sao Paulo 2015, 57, 11–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brummer, E.; Castaneda, E.; Restrepo, A. Paracoccidioidomycosis: An update. Clin. Microbiol. Rev. 1993, 6, 89–117. [Google Scholar] [CrossRef] [PubMed]
- Marques, S.A. Paracoccidioidomycosis: Epidemiological, clinical, diagnostic, and treatment updating. An. Bras. Dermatol. 2013, 88, 700–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, R. New trends in paracoccidioidomycosis epidemiology. J. Fungi. 2017, 3, 1. [Google Scholar] [CrossRef] [Green Version]
- Bellissimo-Rodrigues, F.; Machado, A.A.; Martinez, R. Paracoccidioidomycosis epidemiological features of a 1000-cases series from a hyperendemic area on the southeast of Brazil. Am. J. Trop. Med. Hyg. 2011, 85, 546–550. [Google Scholar] [CrossRef]
- Shikanai-Yasuda, M.A.; de Telles Filho, F.Q.; Mendes, R.P.; Colombo, A.L.; Moretti, M.L. Guidelines in paracoccidioidomycosis. Rev. Soc. Bras. Med. Trop. 2006, 39, 297–310. [Google Scholar] [CrossRef] [Green Version]
- De Castro, L.F.; Ferreira, M.C.; da Silva, R.M.; Blotta, M.H.; Longhi, L.N.; Mamoni, R.L. Characterization of the immune response in human paracoccidioidomycosis. J. Infect. 2013, 67, 470–485. [Google Scholar] [CrossRef]
- Cano, L.; González, A.; Lopera, D.; Naranjo, T.W.; Restrepo, A. Pulmonary paracoccidioidomycosis: Clinical, immunological and histopathological aspects. In Lung Diseases: Selected State of the Art Reviews; Irusen, E.M., Ed.; InTech: Rijeka, Croatia, 2012; pp. 359–392. [Google Scholar]
- Restrepo, A.; Benard, G.; de Castro, C.C.; Agudelo, C.A.; Tobón, A.M. Pulmonary paracoccidioidomycosis. Semin. Resp. Crit. Care Med. 2008, 29, 182–197. [Google Scholar] [CrossRef]
- González, A.; Lenzi, H.L.; Motta, E.M.; Caputo, L.; Restrepo, A.; Cano, L.E. Expression and arrangement of extracellular matrix proteins in the lungs of mice infected with Paracoccidioides brasiliensis conidia. Int. J. Exp. Pathol. 2008, 89, 106–116. [Google Scholar] [CrossRef]
- Naranjo, T.W.; Lopera, D.E.; Diaz-Granados, L.R.; Duque, J.J.; Restrepo, A.; Cano, L.E. Histopathologic and immunologic effects of the itraconazole treatment in a murine model of chronic pulmonary paracoccidioidomycosis. Microbes Infect. 2010, 12, 1153–1162. [Google Scholar] [CrossRef] [PubMed]
- Arango, J.C.; Puerta-Arias, J.D.; Pino-Tamayo, P.A.; Salazar-Peláez, L.M.; Rojas, M.; González, Á. Impaired anti-fibrotic effect of bone marrow-derived mesenchymal stem cell in a mouse model of pulmonary paracoccidioidomycosis. PLoS Negl. Trop. Dis. 2017, 11, e0006006. [Google Scholar] [CrossRef] [PubMed]
- Restrepo, A.; Cano, L.E.; Gonzalez, A. The power of the small: The example of Paracoccidioides brasiliensis conidia. Rev. Inst. Med. Trop. Sao Paulo 2015, 57, 5–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopera, D.; Naranjo, T.; Hidalgo, J.M.; de Oliveira Pascarelli, B.M.; Patiño, J.H.; Lenzi, H.L.; Restrepo, A.; Cano, L.E. Pulmonary abnormalities in mice with paracoccidioidomycosis: A sequential study comparing high resolution computed tomography and pathologic findings. PLoS Negl. Trop. Dis. 2010, 4, e726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, A.N.; Benard, G.; Albuquerque, A.L.; Fujita, C.L.; Magri, A.S.; Salge, J.M.; Restrepo, A.; Shikanai-Yasuda, M.A.; Carvalho, C.R. The lung in paracoccidioidomycosis: New insights into old problems. Clinics 2013, 68, 441–448. [Google Scholar] [CrossRef]
- de Pina, D.R.; Alvarez, M.; Giacomini, G.; Pavan, A.L.; Guedes, C.I.; de Souza Cavalcante, R.; Mendes, R.P.; Paniago, A.M. Paracoccidioidomycosis: Level of pulmonary sequelae in high resolution computed tomography images from patients of two endemic regions of Brazil. Quant. Imaging Med. Surg. 2017, 7, 318–325. [Google Scholar] [CrossRef] [Green Version]
- Neuner, P.; Klosner, G.; Schauer, E.; Pourmojib, M.; Macheiner, W.; Grünwald, C.; Knobler, R.; Schwarz, A.; Luger, T.A.; Schwarz, T. Pentoxifylline in vivo down-regulates the release of IL-1 beta, IL-6, IL-8 and tumor necrosis factor-alpha by human peripheral blood mononuclear cells. Immunology 1994, 83, 262–267. [Google Scholar] [PubMed]
- Berman, B.; Duncan, M.R. Pentoxifylline inhibits normal human dermal fibroblast in vitro proliferation, collagen, glycosaminoglycan, and fibronectin production, and increases collagenase activity. J. Investig. Dermatol. 1989, 92, 605–610. [Google Scholar] [CrossRef] [Green Version]
- Gilhar, A.; Grossman, N.; Kahanovicz, S.; Reuveni, H.; Cohen, S.; Eitan, A. Antiproliferative effect of pentoxifylline on psoriatic and normal epidermis. In vitro and in vivo studies. Acta Derm. Venereol. 1996, 76, 437–441. [Google Scholar]
- Wuyts, W.A.; Willems, S.; Vos, R.; Vanaudenaerde, B.M.; de Vleeschauwer, S.I.; Rinaldi, M.; Vanhooren, H.M.; Geudens, N.; Verleden, S.E.; Demedts, M.G.; et al. Azithromycin reduces pulmonary fibrosis in a bleomycin mouse model. Exp. Lung Res. 2010, 36, 602–614. [Google Scholar] [CrossRef]
- Koch, H.P. Thalidomide and congeners as anti-inflammatory agents. Prog. Med. Chem. 1985, 22, 165–242. [Google Scholar]
- Sampaio, E.P.; Sarno, E.N.; Galilly, R.; Cohn, Z.A.; Kaplan, G. Thalidomide selectively inhibits tumor necrosis factor-alpha production by stimulated human monocytes. J. Exp. Med. 1991, 173, 699–703. [Google Scholar] [CrossRef] [Green Version]
- Haslett, P.A.; Corral, L.G.; Albert, M.; Kaplan, G. Thalidomide costimulates primary human T lymphocytes, preferentially inducing proliferation, cytokine production, and cytotoxic responses in the CD8+ subset. J. Exp. Med. 1998, 187, 1885–1892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corral, L.G.; Haslett, P.A.; Muller, G.W.; Chen, R.; Wong, L.M.; Ocampo, C.J.; Patterson, R.T.; Stirling, D.I.; Kaplan, G. Differential cytokine modulation and T cell activation by two distinct classes of thalidomide analogues that are potent inhibitors of TNF-α. J. Immunol. 1999, 163, 380–386. [Google Scholar] [PubMed]
- D’Amato, R.J.; Loughnan, M.S.; Flynn, E.; Folkman, J. Thalidomide is an inhibitor of angiogenesis. Proc. Natl. Acad. Sci. USA 1994, 91, 4082–4085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choe, J.Y.; Jung, H.J.; Park, K.Y.; Kum, Y.S.; Song, G.G.; Hyun, D.S.; Park, S.H.; Kim, S.K. Anti-fibrotic effect of thalidomide through inhibiting TGF-beta-induced ERK1/2 pathways in bleomycin-induced lung fibrosis in mice. Inflamm. Res. 2010, 59, 177–188. [Google Scholar] [CrossRef] [PubMed]
- Tabata, C.; Tabata, R.; Kadokawa, Y.; Hisamori, S.; Takahashi, M.; Mishima, M.; Nakano, T.; Kubo, H. Thalidomide prevents bleomycininduced pulmonary fibrosis in mice. J. Immunol. 2007, 179, 708–714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sgalla, G.; Cocconcelli, E.; Tonelli, R.; Richeldi, L. Novel drug targets for idiopathic pulmonary fibrosis. Expert Rev. Respir. Med. 2016, 10, 393–405. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues-Vendramini, F.A.V.; Marschalk, C.; Toplak, M.; Macheroux, P.; Bonfim-Mendonça, P.D.S.; Svidzinski, T.I.E.; Seixas, F.A.V.; Kioshima, E.S. Promising new antifungal treatment targeting chorismate synthase from Paracoccidioides brasiliensis. Antimicrob. Agents Chemother. 2019, 63, e01097-18. [Google Scholar] [CrossRef] [Green Version]
- Arango, J.C.; Puerta-Arias, J.D.; Pino-Tamayo, P.A.; Arboleda-Toro, D.; González, Á. Bone marrow-derived mesenchymal stem cells transplantation alters the course of experimental paracoccidioidomycosis by exacerbating the chronic pulmonary inflammatory response. Med. Mycol. 2018, 56, 884–895. [Google Scholar] [CrossRef]
- Morais, E.A.; do Nascimento-Martins, E.M.; Boelone, J.N.; Gomes, D.A.; Goes, A.M. Immunization with recombinant Pb27 protein reduces the levels of pulmonary fibrosis caused by the inflammatory response against Paracoccidioides brasiliensis. Mycopathologia 2015, 179, 31–43. [Google Scholar] [CrossRef]
- Lewis, R.E. Current concepts in antifungal pharmacology. Mayo Clin. Proc. 2011, 86, 805–817. [Google Scholar] [CrossRef] [Green Version]
- Colombo, A.L.; Tobón, A.; Restrepo, A.; Queiroz-Telles, F.; Nucci, M. Epidemiology of endemic systemic fungal infections in Latin America. Med. Mycol. 2011, 49, 785–898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shikanai-Yasuda, M.A.; Mendes, R.P.; Colombo, A.L.; Telles, F.Q.; Kono, A.; Paniago, A.M.M.; Nathan, A.; Valle, A.C.F.D.; Bagagli, E.; Benard, G.; et al. Brazilian guidelines for the clinical management of paracoccidioidomycosis. Epidemiol. Serv. Saude 2018, 27, e0500001. [Google Scholar] [CrossRef] [PubMed]
- De Cavalcante, R.S.; Sylvestre, T.F.; Levorato, A.D.; de Caravalho, L.R.; Mendes, R.P. Comparison between itraconazole and cotrimoxazole in the treatment of paracoccidiodomycosis. PLoS Negl. Trop. Dis. 2014, 8, e2793. [Google Scholar] [CrossRef] [PubMed]
- Muenster, S.; Bode, C.; Diedrich, B.; Jahnert, S.; Weisheit, C.; Steinhagen, F.; Frede, S.; Hoeft, A.; Meyer, R.; Boehm, O.; et al. Antifungal antibiotics modulate the pro-inflammatory cytokine production and phagocytic activity of human monocytes in an in vitro sepsis model. Life Sci. 2015, 141, 128–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macheroux, P.; Schmid, J.; Amrhein, N.; Schaller, A. A unique reaction in a common pathway: Mechanism and function of chorismate synthase in the shikimate pathway. Planta 1999, 207, 325–334. [Google Scholar] [CrossRef]
- Sgalla, G.; Flore, M.; Siciliano, M.; Richeldi, L. Antibody-based therapies for idiopathic pulmonary fibrosis. Expert Opin. Biol. Ther. 2020, 20, 779–786. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Jiang, T.; Feng, Y.; Zhou, M.; Han, X. Monoclonal antibody against laminin receptor 1 inhibits the pulmonary fibrosis induced by bleomycin in rats. Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi 2013, 29, 574–576. [Google Scholar]
- Xu, L.; Yang, D.; Zhu, S.; Gu, J.; Ding, F.; Bian, W.; Rong, Z.; Shen, C. Bleomycin-induced pulmonary fibrosis is attenuated by an antibody against KL-6. Exp. Lung Res. 2013, 39, 241–248. [Google Scholar] [CrossRef]
- Richeldi, L.; Fernández-Pérez, E.R.; Costabel, U.; Albera, C.; Lederer, D.J.; Flaherty, K.R.; Ettinger, N.; Perez, R.; Scholand, M.B.; Goldin, J.; et al. Pamrevlumab, an anti-connective tissue growth factor therapy, for idiopathic pulmonary fibrosis (PRAISE): A phase 2, randomised, double-blind, placebo-controlled trial. Lancet Respir. Med. 2020, 8, 25–33. [Google Scholar] [CrossRef]
- Duguid, M.; Resnick, R.; Sebalusky, B.; Huang, R.; Oliver, J.; Yabkowitz, R.; Subramanian, A.; Genzyme, S.; Framingham, M.A. IL-13 is a driver of pulmonary fibrosis in the Fra-2 transgenic mouse model. Am. J. Respir. Crit. Care Med. 2015, 191, A3441. [Google Scholar]
- Sui, J.N.; Guo, J.; Wang, Z.; Gao, L.; Zhang, H. Effects of tumor necrosis factor-α monoclonal antibody on nuclear factor-κB activation and inducible nitric oxide synthase expression in rats with silicotic fibrosis. Zhonghua Lao Dong Wei Sheng Zhi Ye Bing Za Zhi 2017, 35, 332–336. [Google Scholar] [PubMed]
- Li, J.; He, B.; Weng, B. The therapeutic effect of TGF-beta monoclonal antibody to bleomycin-induced pulmonary fibrosis in rats. Zhonghua Jie He He Hu Xi Za Zhi 1997, 20, 347–349. [Google Scholar]
- Kishi, M.; Aono, Y.; Sato, S.; Koyama, K.; Azuma, M.; Abe, S.; Kawano, H.; Kishi, J.; Toyoda, Y.; Okasaki, H.; et al. Blockade of plateletderived growth factor receptor-β, not receptor-α ameliorates bleomycin-induced pulmonary fibrosis in mice. PLoS ONE 2018, 13, e0209786. [Google Scholar] [CrossRef]
- Faress, J.A.; Nethery, D.E.; Kern, E.F.O.; Eisenberg, R.; Jacono, F.J.; Allen, C.L.; Kern, J.A. Bleomycin-induced pulmonary fibrosis is attenuated by a monoclonal antibody targeting HER2. J. Appl. Physiol. (1985) 2007, 103, 2077–2083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Y.; Geng, Y.; Li, L.; Li, X.; Yan, X.; Fang, Y.; Li, X.; Dong, S.; Liu, X.; Li, X.; et al. Blocking follistatin-like 1 attenuates bleomycin-induced pulmonary fibrosis in mice. J. Exp. Med. 2015, 212, 235–252. [Google Scholar] [CrossRef] [PubMed]
- Mor, A.; Salto, M.S.; Katav, A.; Barashi, N.; Edelshtein, V.; Manetti, M.; Levi, Y.; George, J.; Matucci-Cerinic, M. Blockade of CCL24 with a monoclonal antibody ameliorates experimental dermal and pulmonary fibrosis. Ann. Rheum. Dis. 2019, 78, 1260–1268. [Google Scholar] [CrossRef] [PubMed]
- Besnard, A.G.; Struyf, S.; Guabiraba, R.; Fauconnier, L.; Rouxel, N.; Proost, P.; Uyttenhove, C.; Van Snick, J.; Couillin, I.; Ryffel, B. CXCL6 antibody neutralization prevents lung inflammation and fibrosis in mice in the bleomycin model. J. Leukoc. Biol. 2013, 94, 1317–1323. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, K.; Habiel, D.M.; Jaffar, J.; Binder, U.; Darby, W.G.; Hosking, C.G.; Skerra, A.; Westall, G.P.; Hogaboam, C.M.; Foley, M. Anti-fibrotic Effects of CXCR4-Targeting i-body AD-114 in Preclinical Models of Pulmonary Fibrosis. Sci. Rep. 2018, 8, 3212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elhai, M.; Avouac, J.; Hoffmann-Vold, A.M.; Ruzehaji, N.; Amiar, O.; Ruiz, B.; Brahiti, H.; Ponsoye, M.; Fréchet, M.; Burgevin, A.; et al. OX40L blockade protects against inflammation-driven fibrosis. Proc. Natl. Acad. Sci. USA 2016, 113, E3901–E3910. [Google Scholar] [CrossRef] [Green Version]
- Xiong, S.; Guo, R.; Yang, Z.; Xu, L.; Du, L.; Li, R.; Xiao, F.; Wang, Q.; Zhu, M.; Pan, X. Treg depletion attenuates irradiation-induced pulmonary fibrosis by reducing fibrocyte accumulation, inducing Th17 response, and shifting IFN-γ, IL-12/IL-4, IL-5 balance. Immunobiology 2015, 220, 1284–1291. [Google Scholar] [CrossRef]
- Parker, J.M.; Glaspole, I.N.; Lancaster, L.H.; Haddad, T.J.; She, D.; Roseti, S.L.; Fiening, J.P.; Grant, E.P.; Kell, C.M.; Flaherty, K.R. A Phase 2 Randomized Controlled Study of Tralokinumab in Subjects with Idiopathic Pulmonary Fibrosis. Am. J. Respir. Crit. Care Med. 2018, 197, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Pino-Tamayo, P.A.; Puerta-Arias, J.D.; Lopera, D.; Urán-Jiménez, M.E.; González, Á. Depletion of neutrophils exacerbates the early inflammatory immune response in lungs of mice infected with Paracoccidioides brasiliensis. Med. Inflamm. 2016, 2016, 3183285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez, A.; Cano, L.E. Participation of the polymorphonuclear neutrophil in the immune response against Paracoccidioides brasiliensis [Participación del polimorfonuclear neutrófilo en la respuesta inmune contra Paracoccidioides brasiliensis]. Biomedica 2001, 21, 264–274. [Google Scholar] [CrossRef] [Green Version]
- Ghadiri, M.; Young, P.M.; Traini, D. Cell-based therapies for the treatment of idiopathic pulmonary fibrosis (IPF) disease. Rev. Expert Opin. Biol. Ther. 2016, 16, 375–387. [Google Scholar] [CrossRef] [PubMed]
- Srour, N.; Thébaud, B. Mesenchymal stromal cells in animal bleomycin pulmonary fibrosis models: A systematic review. Stem Cells Transl. Med. 2015, 4, 1500–1510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glassberg, M.K.; Toonkel, R.L. Moving stem cell therapy to patients with idiopathic pulmonary fibrosis. Respirology 2014, 9, 950–951. [Google Scholar] [CrossRef]
- Yang, J.; Jia, Z. Cell-based therapy in lung regenerative medicine. Regen. Med. Res. 2014, 2, 7–13. [Google Scholar] [CrossRef] [Green Version]
- Zhang, E.; Yang, Y.; Zhang, J.; Ding, G.; Chen, S.; Peng, C.; Lavin, M.F.; Yeo, A.J.; Du, Z.; Shao, H. Efficacy of bone marrow mesenchymal stem cell transplantation in animal models of pulmonary fibrosis after exposure to bleomycin: A meta-analysis. Exp. Ther. Med. 2019, 17, 2247–2255. [Google Scholar] [CrossRef]
- Cores, J.; Hensley, M.T.; Kinlaw, K.; Rikard, S.M.; Dinh, P.U.; Paudel, D.; Tang, J.; Vandergriff, A.C.; Allen, T.A.; Li, Y.; et al. Safety and efficacy of allogeneic lung spheroid cells in a mismatched rat model of pulmonary fibrosis. Stem Cells Transl. Med. 2017, 6, 1905–1916. [Google Scholar] [CrossRef] [Green Version]
- Chu, K.A.; Wang, S.Y.; Yeh, C.C.; Fu, T.W.; Fu, Y.Y.; Ko, T.L.; Chiu, M.M.; Chen, T.H.; Tsai, P.J.; Fu, Y.S. Reversal of bleomycin-induced rat pulmonary fibrosis by a xenograft of human umbilical mesenchymal stem cells from Wharton’s jelly. Theranostics 2019, 9, 6646–6664. [Google Scholar] [CrossRef]
- Lan, Y.W.; Theng, S.M.; Huang, T.T.; Choo, K.B.; Chen, C.M.; Kuo, H.P.; Chong, K.Y. Oncostatin M-Preconditioned mesenchymal stem cells alleviate bleomycin-induced pulmonary fibrosis through paracrine effects of the hepatocyte growth factor. Stem Cells Transl. Med. 2017, 6, 1006–1017. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; He, Z.; Gao, Y.; Zheng, R.; Zhang, X.; Zhao, L.; Tan, M. Induced pluripotent stem cells inhibit bleomycin-induced pulmonary fibrosis in mice through suppressing TGF- b1/Smad-mediated epithelial to mesenchymal transition. Front. Pharmacol. 2016, 7, 430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Echeverri, C.; Puerta-Arias, J.D.; González, Á. Paracoccidioides brasiliensis activates mesenchymal stem cells through TLR2, TLR4, and Dectin-1. Med. Mycol. 2020, myaa039. [Google Scholar] [CrossRef] [PubMed]
- Reis, B.S.; Fernandes, V.C.; Martins, E.M.; Serakides, R.; Goes, A.M. Protective immunity induced by rPb27 of Paracoccidioides brasiliensis. Vaccine 2008, 26, 5461–5469. [Google Scholar] [CrossRef]
- Morais, E.A.; Martins, E.M.N.; Fernandes, V.C.; Santos, I.G.D.; Silva, G.A.B.; Gomes, D.A.; Miranda, A. Immunization with rPb27 protects mice from the disruption of VEGF signaling in Paracoccidioides brasiliensis infection. J. Life Sci. 2013, 7, 913–927. [Google Scholar]



{kind=link}
{kind=link}
| Therapy Approach | Findings | References |
|---|---|---|
| Drugs | ||
| ITC |  Fungal burden, inflammatory response; and PF if treatment was started an early time post-infection (4 wk.) Fungal burden, inflammatory response; and PF if treatment was started an early time post-infection (4 wk.) IL-1β, IL-13, TNF-α and TGF-β IL-1β, IL-13, TNF-α and TGF-β | [23] |
| PTX |  Fungal burden, inflammatory response; and PF if treatment was started an early time post-infection (4 wk.) Fungal burden, inflammatory response; and PF if treatment was started an early time post-infection (4 wk.) GM-CSF, IL-12p70, IL-10, IL-13 GM-CSF, IL-12p70, IL-10, IL-13 RANTES RANTES | [9] |
| ITC + PTX |  Fungal burden, inflammatory response; and PF even if treatment was started an advanced time post-infection (8 wk.) Fungal burden, inflammatory response; and PF even if treatment was started an advanced time post-infection (8 wk.) Hydroxyproline Hydroxyproline IL-1β; IL-6, IL-17, TGF-β1 IL-1β; IL-6, IL-17, TGF-β1 IL-10 IL-10 | [8,12] |
| ITC + AZT |  PF, CCL3, IL-17, IFN-γ, VEGF PF, CCL3, IL-17, IFN-γ, VEGF IL-1β, IL-6, IL-10, TGF-β1 IL-1β, IL-6, IL-10, TGF-β1 | [12] |
| ITC + Thal |  PF PF IL-1β, IL-6, IL-10, IL-12, IL-17, TGF-β1, VEGF, IFN-γ, CCL3 IL-1β, IL-6, IL-10, IL-12, IL-17, TGF-β1, VEGF, IFN-γ, CCL3 | [12] |
| CMX + PTX |  PF, CCL3 PF, CCL3 IL-17, TGF-β1 IL-17, TGF-β1 | [12] |
| CMX + AZT |  Inflammatory response, hydroxyproline Inflammatory response, hydroxyproline TGF-β1 TGF-β1 TNF-α, IL-10 TNF-α, IL-10 | [12] |
| CMX + Thal |  PF PF IL-1β, IL-6, IL-17, TGF-β1, VEGF, IFN-γ, CCL3 IL-1β, IL-6, IL-17, TGF-β1, VEGF, IFN-γ, CCL3 | [12] |
| CP1 |  Fungal burden, Inflammatory response, PF Fungal burden, Inflammatory response, PF | [41] |
| Biological | ||
| mAbs | ||
| anti-neutrophils |  Fungal burden, Inflammatory response, PF Fungal burden, Inflammatory response, PF IL-17, TNF-α, TGF-β1, TGF-β3, MMP-12, MMP-14 IL-17, TNF-α, TGF-β1, TGF-β3, MMP-12, MMP-14 MMP8, TIMP-2 MMP8, TIMP-2 | [10] |
| anti-neutrophil + ITC |  Fungal burden, Inflammatory response, PF Fungal burden, Inflammatory response, PF IL-1β, IL-6, IL-17, IL-10, TNF-α, TGF-β1, MMP-1α, IL-1β, IL-6, IL-17, IL-10, TNF-α, TGF-β1, MMP-1α, GATA-3, RORc, Ahr, GATA-3, RORc, Ahr, MMP-15, TIMP-1, TIMP-2 MMP-15, TIMP-1, TIMP-2 | [11] |
| MSCs | ||
| BMMSCs |  Fungal burden, inflammatory response, PF Fungal burden, inflammatory response, PF IL-6, IL-9, GM-CSF, CXCL1, CXCL9, CCL5 IL-6, IL-9, GM-CSF, CXCL1, CXCL9, CCL5 Collagen-3α1, TGF-β3, MMP-15 Collagen-3α1, TGF-β3, MMP-15 | [24,42] |
| BMMSCs + ITC |  Fungal burden, inflammatory response, PF Fungal burden, inflammatory response, PF Col3α1, TGF-β3, MMP-8, MMP-12, TIMP-1 Col3α1, TGF-β3, MMP-8, MMP-12, TIMP-1 TIMP-2 TIMP-2 | [24] |
| Vaccines | ||
| rPb27 |  PF, collagen PF, collagen IFN-γ, TGF-β, IL-10 IFN-γ, TGF-β, IL-10 CCR7 CCR7 Activity of caspase 3 Activity of caspase 3 | [43] |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
González, Á. The Therapy of Pulmonary Fibrosis in Paracoccidioidomycosis: What Are the New Experimental Approaches? J. Fungi 2020, 6, 217. https://doi.org/10.3390/jof6040217
González Á. The Therapy of Pulmonary Fibrosis in Paracoccidioidomycosis: What Are the New Experimental Approaches? Journal of Fungi. 2020; 6(4):217. https://doi.org/10.3390/jof6040217
Chicago/Turabian StyleGonzález, Ángel. 2020. "The Therapy of Pulmonary Fibrosis in Paracoccidioidomycosis: What Are the New Experimental Approaches?" Journal of Fungi 6, no. 4: 217. https://doi.org/10.3390/jof6040217
APA StyleGonzález, Á. (2020). The Therapy of Pulmonary Fibrosis in Paracoccidioidomycosis: What Are the New Experimental Approaches? Journal of Fungi, 6(4), 217. https://doi.org/10.3390/jof6040217