Oral Administration of Lactobacillus helveticus LA401 and Lactobacillus gasseri LA806 Combination Attenuates Oesophageal and Gastrointestinal Candidiasis and Consequent Gut Inflammation in Mice

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. C. albicans and Bacterial Strains
2.2. Murine Model of Gastrointestinal Candidiasis
2.3. Quantification of the Number of Viable C. albicans in the Stools
2.4. Quantification of C. albicans in the Gastrointestinal Tract and Microbiota Analysis Using Real-Time PCR
2.5. Gene Expression Analysis by Reverse Transcription and Real-Time PCR
2.6. Preparation of Mouse Peritoneal Macrophages
2.7. Killing Assay
2.8. Binding and Phagocytosis Assays
2.9. ROS Quantification
2.10. Measurement of Nitrites (NO2−)
2.11. Cytokine Measurement by ELISA
2.12. Statistical Analysis
3. Results
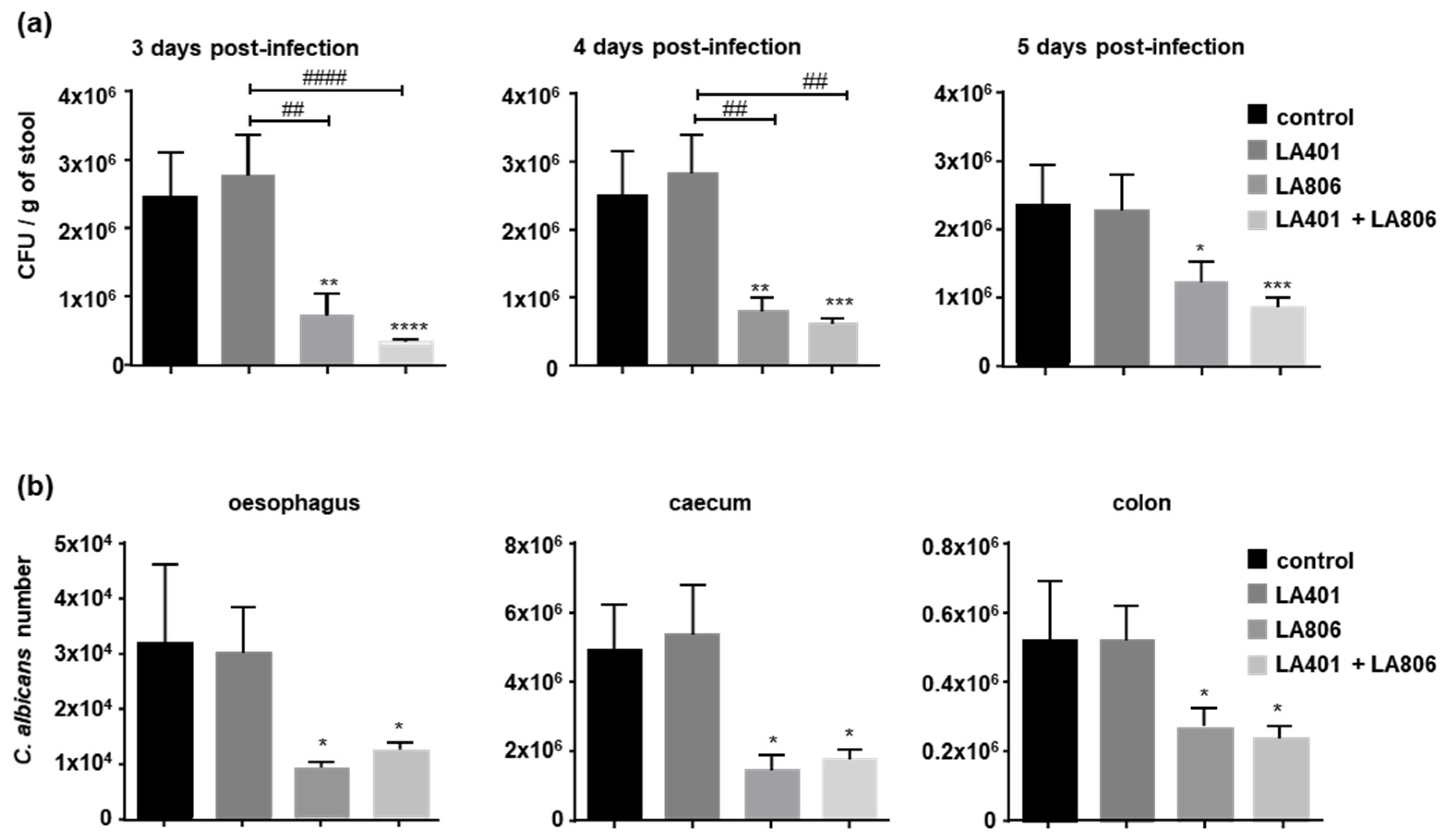
3.1. Lactobacillus gasseri LA806 Alone or Combined with Lactobacillus helveticus LA401 Effectively Reduces C. albicans Burden in Mice Gastrointestinal Tract
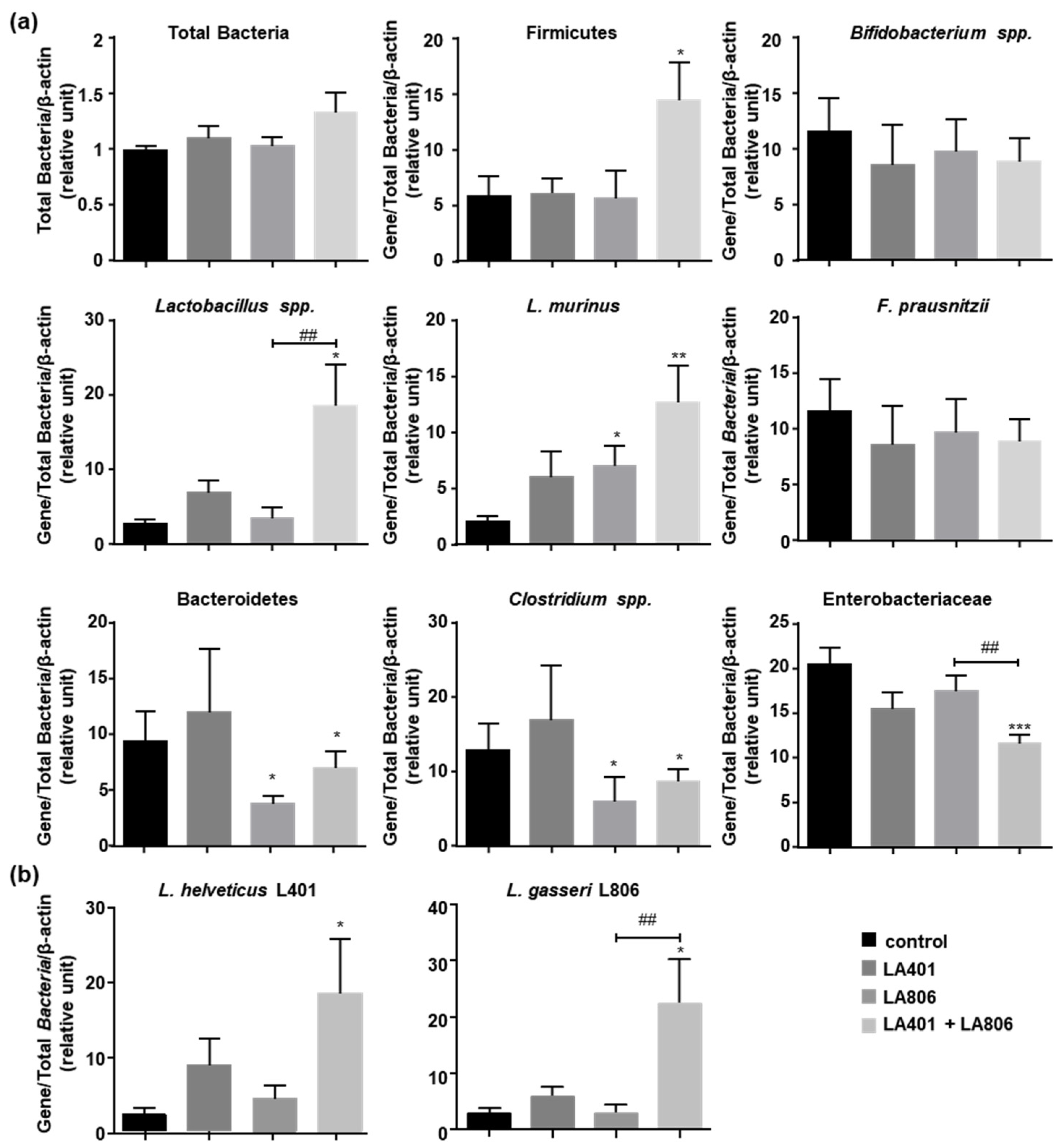
3.2. Lactobacillus gasseri LA806 Alone or Combined with Lactobacillus helveticus LA401 Modulates Gut Microbiota
3.3. Lactobacillus helveticus LA401 and Lactobacillus gasseri LA806 Co-Administration Improves Their Respective Intestinal Colonization
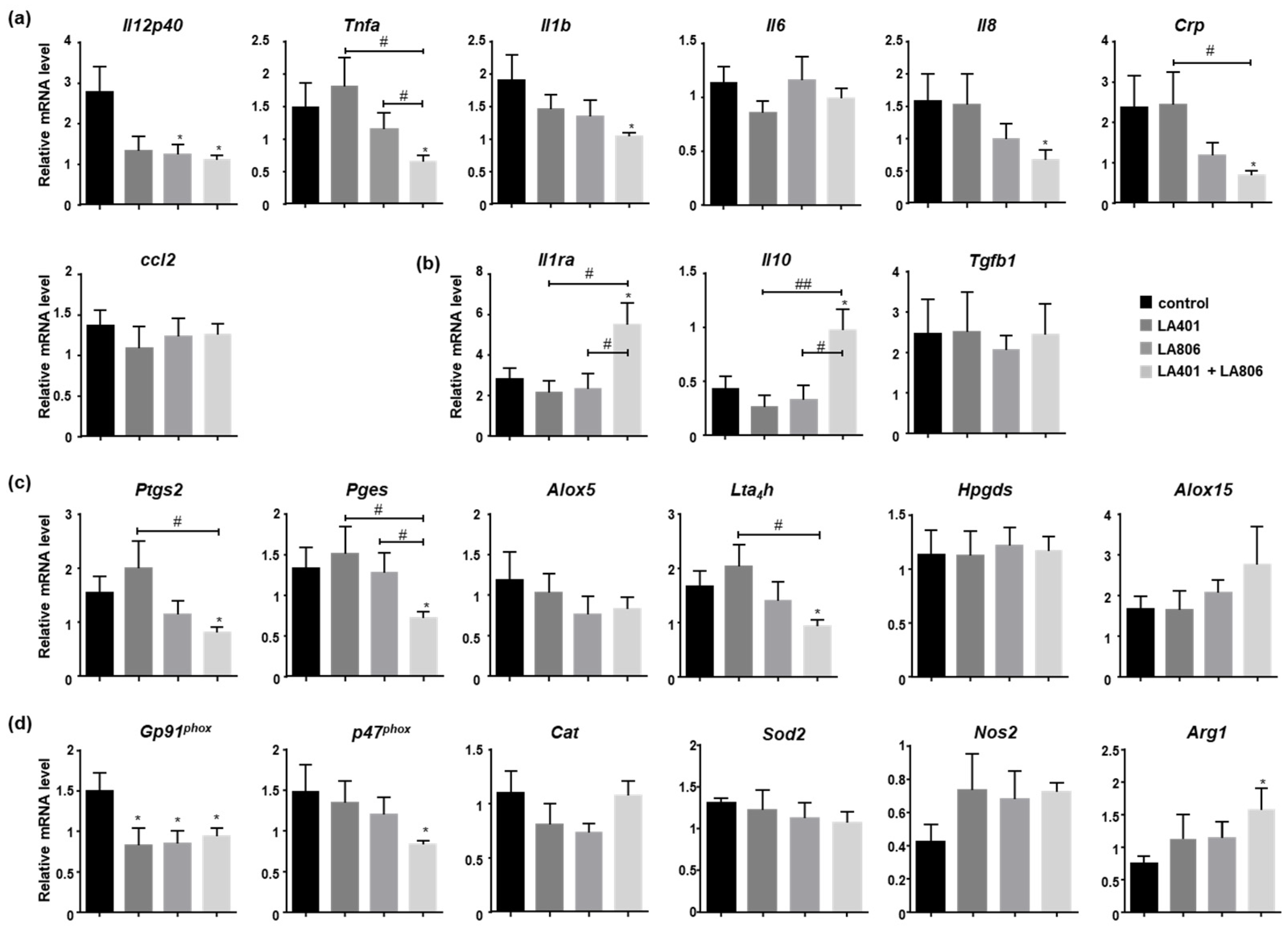
3.4. Lactobacillus helveticus LA401 and Lactobacillus gasseri LA806 Combination Dampens Inflammation and Oxidative Stress in the Colon of Mice with Gastrointestinal Candidiasis
3.5. Lactobacillus helveticus LA401 and/or Lactobacillus gasseri LA806 Improve the Fungicidal Properties of Macrophages While Controlling Their Inflammatory Status
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gulati, M.; Nobile, C.J. Candida Albicans Biofilms: Development, Regulation, and Molecular Mechanisms. Microbes Infect. 2016, 18, 310–321. [Google Scholar] [CrossRef] [Green Version]
- Poulain, D. Candida Albicans, Plasticity and Pathogenesis. Crit. Rev. Microbiol. 2015, 41, 208–217. [Google Scholar] [CrossRef]
- Ribeiro, F.C.; Rossoni, R.D.; de Barros, P.P.; Santos, J.D.; Fugisaki, L.R.O.; Leão, M.P.V.; Junqueira, J.C. Action Mechanisms of Probiotics on Candida Spp. and Candidiasis Prevention: An Update. J. Appl. Microbiol. 2019, 129, 175–185. [Google Scholar] [CrossRef] [Green Version]
- Duplaga, K.K.; Krawczyk, A.; Oleksiak, A.S.; Salamon, D.; Wędrychowicz, A.; Fyderek, K.; Gosiewski, T. Dependence of Colonization of the Large Intestine by Candida on the Treatment of Crohn’s Disease. Pol. J. Microbiol. 2019, 68, 121–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumamoto, C.A. Inflammation and Gastrointestinal Candida Colonization. Curr. Opin. Microbiol. 2011, 14, 386–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Chen, D.; Yu, B.; He, J.; Zheng, P.; Mao, X.; Yu, J.; Luo, J.; Tian, G.; Huang, Z.; et al. Fungi in Gastrointestinal Tracts of Human and Mice: From Community to Functions. Microb. Ecol. 2018, 75, 821–829. [Google Scholar] [CrossRef]
- Sokol, H.; Leducq, V.; Aschard, H.; Pham, H.-P.; Jegou, S.; Landman, C.; Cohen, D.; Liguori, G.; Bourrier, A.; Larmurier, I.N.; et al. Fungal Microbiota Dysbiosis in IBD. Gut 2017, 66, 1039–1048. [Google Scholar] [CrossRef] [Green Version]
- Pérez, J.C. Candida Albicans Dwelling in the Mammalian Gut. Curr. Opin. Microbiol. 2019, 52, 41–46. [Google Scholar] [CrossRef]
- Kühl, A.A.; Erben, U.; Kredel, L.I.; Siegmund, B. Diversity of Intestinal Macrophages in Inflammatory Bowel Diseases. Front. Immunol. 2015, 6, 613. [Google Scholar] [CrossRef] [Green Version]
- Coste, A.; Lagane, C.; Filipe, C.; Authier, H.; Galès, A.; Bernad, J.; Echinard, V.D.; Lepert, J.-C.; Balard, P.; Linas, M.-D.; et al. IL-13 Attenuates Gastrointestinal Candidiasis in Normal and Immunodeficient RAG-2(-/-) Mice via Peroxisome Proliferator-Activated Receptor-Gamma Activation. J. Immunol. 2008, 180, 4939–4947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mantovani, A.; Sica, A.; Sozzani, S.; Allavena, P.; Vecchi, A.; Locati, M. The Chemokine System in Diverse Forms of Macrophage Activation and Polarization. Trends Immunol. 2004, 25, 677–686. [Google Scholar] [CrossRef] [PubMed]
- Mosser, D.M.; Edwards, J.P. Exploring the Full Spectrum of Macrophage Activation. Nat. Rev. Immunol. 2008, 8, 958–969. [Google Scholar] [CrossRef] [PubMed]
- Coste, A.; Dubourdeau, M.; Linas, M.D.; Cassaing, S.; Lepert, J.-C.; Balard, P.; Chalmeton, S.; Bernad, J.; Orfila, C.; Séguéla, J.-P.; et al. PPARgamma Promotes Mannose Receptor Gene Expression in Murine Macrophages and Contributes to the Induction of This Receptor by IL. Immunity 2003, 19, 329–339. [Google Scholar] [CrossRef] [Green Version]
- Galès, A.; Conduché, A.; Bernad, J.; Lefevre, L.; Olagnier, D.; Béraud, M.; Blondel, G.M.; Linas, M.-D.; Auwerx, J.; Coste, A.; et al. PPARγ Controls Dectin-1 Expression Required for Host Antifungal Defense against Candida Albicans. PLoS Pathog. 2010, 6. [Google Scholar] [CrossRef] [Green Version]
- Lefèvre, L.; Galès, A.; Olagnier, D.; Bernad, J.; Perez, L.; Burcelin, R.; Valentin, A.; Auwerx, J.; Pipy, B.; Coste, A. PPARγ Ligands Switched High Fat Diet-Induced Macrophage M2b Polarization toward M2a Thereby Improving Intestinal Candida Elimination. PLoS ONE 2010, 5, e12828. [Google Scholar] [CrossRef]
- Netea, M.G.; Gow, N.A.R.; Munro, C.A.; Bates, S.; Collins, C.; Ferwerda, G.; Hobson, R.P.; Bertram, G.; Hughes, H.B.; Jansen, T.; et al. Immune Sensing of Candida Albicans Requires Cooperative Recognition of Mannans and Glucans by Lectin and Toll-like Receptors. J. Clin. Investig. 2006, 116, 1642–1650. [Google Scholar] [CrossRef]
- Matsubara, V.H.; Bandara, H.M.H.N.; Mayer, M.P.A.; Samaranayake, L.P. Probiotics as Antifungals in Mucosal Candidiasis. Clin. Infect. Dis. 2016, 62, 1143–1153. [Google Scholar] [CrossRef] [Green Version]
- Hayama, K.; Ishijima, S.; Ono, Y.; Izumo, T.; Ida, M.; Shibata, H.; Abe, S. Protective activity of S-PT84, a heat-killed preparation of Lactobacillus pentosus, against oral and gastric candidiasis in an experimental murine model. Med. Mycol. J. 2014, 55, J123–J129. [Google Scholar] [CrossRef] [Green Version]
- Hu, L.; Zhou, M.; Young, A.; Zhao, W.; Yan, Z. In Vivo Effectiveness and Safety of Probiotics on Prophylaxis and Treatment of Oral Candidiasis: A Systematic Review and Meta-Analysis. BMC Oral Health 2019, 19, 140. [Google Scholar] [CrossRef]
- Shenoy, A.; Gottlieb, A. Probiotics for Oral and Vulvovaginal Candidiasis: A Review. Dermatol. Ther. 2019, 32, e12970. [Google Scholar] [CrossRef]
- Wcislo, M.Z.; Brzozowski, T.; Budak, A.; Kwiecien, S.; Sliwowski, Z.; Drozdowicz, D.; Trojanowska, D.; Rudnicka-Sosin, L.; Mach, T.; Konturek, S.J.; et al. Effect of Candida Colonization on Human Ulcerative Colitis and the Healing of Inflammatory Changes of the Colon in the Experimental Model of Colitis Ulcerosa. J. Physiol. Pharmacol. Off. J. Pol. Physiol. Soc. 2009, 60, 107–118. [Google Scholar]
- Lam, S.; Zuo, T.; Ho, M.; Chan, F.K.L.; Chan, P.K.S.; Ng, S.C. Review Article: Fungal Alterations in Inflammatory Bowel Diseases. Aliment. Pharmacol. Ther. 2019, 50, 1159–1171. [Google Scholar] [CrossRef] [PubMed]
- Wagner, R.D.; Warner, T.; Roberts, L.; Farmer, J.; Dohnalek, M.; Hilty, M.; Balish, E. Variable Biotherapeutic Effects of Lactobacillus Acidophilus Isolates on Orogastric and Systemic Candidiasis in Immunodeficient Mice. Rev. Iberoam. Micol. 1998, 15, 271–276. [Google Scholar] [PubMed]
- Azad, M.A.K.; Sarker, M.; Wan, D. Immunomodulatory Effects of Probiotics on Cytokine Profiles. Available online: https://www.hindawi.com/journals/bmri/2018/8063647/ (accessed on 9 July 2020).
- Olivares, M.; Ropero, M.P.D.; Gómez, N.; Villoslada, F.L.; Sierra, S.; Maldonado, J.A.; Martín, R.; Rodríguez, J.M.; Xaus, J. The Consumption of Two New Probiotic Strains, Lactobacillus Gasseri CECT 5714 and Lactobacillus Coryniformis CECT 5711, Boosts the Immune System of Healthy Humans. Int. Microbiol. Off. J. Span. Soc. Microbiol. 2006, 9, 47–52. [Google Scholar]
- Xiao, L.; Ding, G.; Ding, Y.; Deng, C.; Ze, X.; Chen, L.; Zhang, Y.; Song, L.; Yan, H.; Liu, F.; et al. Effect of Probiotics on Digestibility and Immunity in Infants: A Study Protocol for a Randomized Controlled Trial. Medicine (Baltimore) 2017, 96, e5953. [Google Scholar] [CrossRef]
- Galdeano, C.M.; de LeBlanc, A.d.M.; Carmuega, E.; Weill, R.; Perdigón, G. Mechanisms Involved in the Immunostimulation by Probiotic Fermented Milk. J. Dairy Res. 2009, 76, 446–454. [Google Scholar] [CrossRef]
- Chiba, Y.; Shida, K.; Nagata, S.; Wada, M.; Bian, L.; Wang, C.; Shimizu, T.; Yamashiro, Y.; Shibata, J.K.; Nanno, M.; et al. Well-Controlled Proinflammatory Cytokine Responses of Peyer’s Patch Cells to Probiotic Lactobacillus Casei. Immunology 2010, 130, 352–362. [Google Scholar] [CrossRef]
- Benmoussa, K.; Authier, H.; Prat, M.; AlaEddine, M.; Lefèvre, L.; Rahabi, M.C.; Bernad, J.; Aubouy, A.; Bonnafé, E.; Leprince, J.; et al. P17, an Original Host Defense Peptide from Ant Venom, Promotes Antifungal Activities of Macrophages through the Induction of C-Type Lectin Receptors Dependent on LTB4-Mediated PPARγ Activation. Front. Immunol. 2017, 8, 1650. [Google Scholar] [CrossRef]
- Lefèvre, L.; Authier, H.; Stein, S.; Majorel, C.; Couderc, B.; Dardenne, C.; Eddine, M.A.; Meunier, E.; Bernad, J.; Valentin, A.; et al. LRH-1 Mediates Anti-Inflammatory and Antifungal Phenotype of IL-13-Activated Macrophages through the PPARγ Ligand Synthesis. Nat. Commun. 2015, 6, 6801. [Google Scholar] [CrossRef] [Green Version]
- Khan, Z.; Mustafa, A.S.; Alam, F.F. Real-Time LightCycler Polymerase Chain Reaction and Melting Temperature Analysis for Identification of Clinically Important Candida spp. J. Microbiol. Immunol. Infect. 2009, 42, 290–295. [Google Scholar]
- Carroll, I.M.; Chang, Y.-H.; Park, J.; Sartor, R.B.; Ringel, Y. Luminal and Mucosal-Associated Intestinal Microbiota in Patients with Diarrhea-Predominant Irritable Bowel Syndrome. Gut Pathog. 2010, 2, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, X.; Xia, X.; Tang, R.; Zhou, J.; Zhao, H.; Wang, K. Development of a Real-Time PCR Method for Firmicutes and Bacteroidetes in Faeces and Its Application to Quantify Intestinal Population of Obese and Lean Pigs. Lett. Appl. Microbiol. 2008, 47, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Barman, M.; Unold, D.; Shifley, K.; Amir, E.; Hung, K.; Bos, N.; Salzman, N. Enteric Salmonellosis Disrupts the Microbial Ecology of the Murine Gastrointestinal Tract. Infect. Immun. 2008, 76, 907–915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehman, A.; Sina, C.; Gavrilova, O.; Häsler, R.; Ott, S.; Baines, J.F.; Schreiber, S.; Rosenstiel, P. Nod2 Is Essential for Temporal Development of Intestinal Microbial Communities. Gut 2011, 60, 1354–1362. [Google Scholar] [CrossRef]
- Bindels, L.B.; Beck, R.; Schakman, O.; Martin, J.C.; De Backer, F.; Sohet, F.M.; Dewulf, E.M.; Pachikian, B.D.; Neyrinck, A.M.; Thissen, J.-P.; et al. Restoring Specific Lactobacilli Levels Decreases Inflammation and Muscle Atrophy Markers in an Acute Leukemia Mouse Model. PLoS ONE 2012, 7, e37971. [Google Scholar] [CrossRef] [Green Version]
- Iliev, I.D.; Funari, V.A.; Taylor, K.D.; Nguyen, Q.; Reyes, C.N.; Strom, S.P.; Brown, J.; Becker, C.A.; Fleshner, P.R.; Dubinsky, M.; et al. Interactions between Commensal Fungi and the C-Type Lectin Receptor Dectin-1 Influence Colitis. Science 2012, 336, 1314–1317. [Google Scholar] [CrossRef] [Green Version]
- Tang, C.; Kamiya, T.; Liu, Y.; Kadoki, M.; Kakuta, S.; Oshima, K.; Hattori, M.; Takeshita, K.; Kanai, T.; Saijo, S.; et al. Inhibition of Dectin-1 Signaling Ameliorates Colitis by Inducing Lactobacillus-Mediated Regulatory T Cell Expansion in the Intestine. Cell Host Microbe 2015, 18, 183–197. [Google Scholar] [CrossRef] [Green Version]
- Taverniti, V.; Guglielmetti, S. Health-Promoting Properties of Lactobacillus Helveticus. Front. Microbiol. 2012, 3, 392. [Google Scholar] [CrossRef] [Green Version]
- Heung, L.J. Monocytes and the Host Response to Fungal Pathogens. Front. Cell. Infect. Microbiol. 2020, 10, 34. [Google Scholar] [CrossRef]
- Silva, M.P.; Rossoni, R.D.; CamposJunqueira, J.; Jorge, A.O.C. Probiotics for Prevention and Treatment of Candidiasis and Other Infectious Diseases: Lactobacillus Spp. and Other Potential Bacterial Species. Probiot. Prebiot. Hum. Nutr. Health 2016. [Google Scholar] [CrossRef] [Green Version]
- Ramírez, L.M.R.; Solano, R.A.P.; Alonso, S.L.C.; Moreno Guerrero, S.S.; Pacheco, A.R.; García Garibay, M.; Eslava, C. Probiotic Lactobacillus Strains Stimulate the Inflammatory Response and Activate Human Macrophages. J. Immunol. Res. 2017, 2017, 4607491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joo, H.-M.; Kim, K.-A.; Myoung, K.-S.; Ahn, Y.-T.; Lee, J.-H.; Huh, C.-S.; Han, M.J.; Kim, D.-H. Lactobacillus Helveticus HY7801 Ameliorates Vulvovaginal Candidiasis in Mice by Inhibiting Fungal Growth and NF-ΚB Activation. Int. Immunopharmacol. 2012, 14, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Parolin, C.; Marangoni, A.; Laghi, L.; Foschi, C.; Ñahui Palomino, R.A.; Calonghi, N.; Cevenini, R.; Vitali, B. Isolation of Vaginal Lactobacilli and Characterization of Anti-Candida Activity. PLoS ONE 2015, 10, e0131220. [Google Scholar] [CrossRef] [PubMed]
- Terayama, Y.; Matsuura, T.; Uchida, M.; Narama, I.; Ozaki, K. Probiotic (Yogurt) Containing Lactobacillus Gasseri OLL2716 Is Effective for Preventing Candida Albicans-Induced Mucosal Inflammation and Proliferation in the Forestomach of Diabetic Rats. Histol. Histopathol. 2016, 31, 689–697. [Google Scholar] [CrossRef]
- Azad, A.K.; Sarker, M.; Li, T.; Yin, J. Probiotic Species in the Modulation of Gut Microbiota: An Overview. BioMed Res. Int. 2018, 2018, 9478630. [Google Scholar] [CrossRef] [Green Version]
- Bertolini, M.; Ranjan, A.; Thompson, A.; Diaz, P.I.; Sobue, T.; Maas, K.; Bagtzoglou, A.D. Candida Albicans Induces Mucosal Bacterial Dysbiosis That Promotes Invasive Infection. PLoS Pathog. 2019, 15, e1007717. [Google Scholar] [CrossRef]
- Zeng, M.; Inohara, N.; Nuñez, G. Mechanisms of Inflammation-Driven Bacterial Dysbiosis in the Gut. Mucosal Immunol. 2017, 10, 18–26. [Google Scholar] [CrossRef] [Green Version]
- Nira, H.K.; Suzuki, C.; Kobayashi, M.; Sasaki, K.; Mizumachi, K. Inhibition of Leukotriene B4 Production in Murine Macrophages by Lactic Acid Bacteria. Int. J. Food Microbiol. 2009, 129, 321–324. [Google Scholar] [CrossRef]
- Alard, J.; Peucelle, V.; Boutillier, D.; Breton, J.; Kuylle, S.; Pot, B.; Holowacz, S.; Grangette, C. New Probiotic Strains for Inflammatory Bowel Disease Management Identified by Combining in Vitro and in Vivo Approaches. Benef. Microbes 2018, 9, 317–331. [Google Scholar] [CrossRef]
- Holowacz, S.; Guinobert, I.; Guilbot, A.; Hidalgo, S.; Bisson, J.F. A Mixture of Five Bacterial Strains Attenuates Skin Inflammation in Mice. Anti Inflamm. Anti Allergy Agents Med. Chem. 2018, 17, 125–137. [Google Scholar] [CrossRef]
- Kobatake, E.; Nakagawa, H.; Seki, T.; Miyazaki, T. Protective Effects and Functional Mechanisms of Lactobacillus Gasseri SBT2055 against Oxidative Stress. PLoS ONE 2017, 12, e0177106. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Evivie, S.E.; Lu, J.; Jiao, Y.; Wang, C.; Li, Z.; Liu, F.; Huo, G. Lactobacillus Helveticus KLDS1.8701 Alleviates d-Galactose-Induced Aging by Regulating Nrf-2 and Gut Microbiota in Mice. Food Funct. 2018, 9, 6586–6598. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.-O.; Kim, H.-J.; Kim, Y.-J.; Kang, M.-J.; Kwon, J.-W.; Seo, J.-H.; Kim, H.Y.; Kim, B.-J.; Yu, J.; Hong, S.-J. Asthma Prevention by Lactobacillus Rhamnosus in a Mouse Model Is Associated With CD4(+)CD25(+)Foxp3(+) T Cells. Allergy Asthma Immunol. Res. 2012, 4, 150–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, M.M.; Saio, M.; Yamashita, H.; Tanaka, H.; Takami, T.; Ezaki, T.; Inagaki, N. Lactobacillus Acidophilus Strain L-92 Induces CD4+CD25+Foxp3+ Regulatory T Cells and Suppresses Allergic Contact Dermatitis. Biol. Pharm. Bull. 2012, 35, 612–616. [Google Scholar] [CrossRef] [Green Version]
- Takahara, K.; Tokieda, S.; Nagaoka, K.; Inaba, K. Efficient Capture of Candida Albicans and Zymosan by SIGNR1 Augments TLR2-Dependent TNF-α Production. Int. Immunol. 2012, 24, 89–96. [Google Scholar] [CrossRef] [Green Version]
- Delcenserie, V.; Martel, D.; Lamoureux, M.; Amiot, J.; Boutin, Y.; Roy, D. Immunomodulatory Effects of Probiotics in the Intestinal Tract. Curr. Issues Mol. Biol. 2008, 10, 37–54. [Google Scholar]
- Galdeano, C.M.; Perdigón, G. The Probiotic Bacterium Lactobacillus Casei Induces Activation of the Gut Mucosal Immune System through Innate Immunity. Clin. Vaccine Immunol. 2006, 13, 219–226. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Universal Name | 5’-3’ Sequence | |
|---|---|---|---|
| Candida spp. [31] | sense | TCGCATCGATGAAGAACGCAGC | |
| antisense | TCTTTTCCTCCGCTTATTGATATGC | ||
| Clostridium spp. [32] | sense | CGGTACCTGACTAAGAAGC | |
| antisense | AGTTTYATTCTTGCGAACG | ||
| Bifidobacterium spp. [32] | sense | GGGTGGTAATGCCGGATG | |
| antisense | TAAGCGATGGACTTTCACACC | ||
| Lactobacillus spp. [32] | sense | AGCAGTAGGGAATCTTCCA | |
| antisense | CACCGCTACACATGGAG | ||
| Total bacteria [33] | sense | Eub338F | ACTCCTACGGGAGGCAGCAG |
| antisense | Eub518R | ATTACCGCGGCTGCTGG | |
| Bacteroidetes [33] | sense | Bact934F | GGARCATGTGGTTTAATTCGATGAT |
| antisense | Bact1060R | AGCTGACGACAACCATGCAG | |
| Firmicutes [33] | sense | Firm934F | GGAGYATGTGGTTTAATTCGAAGCA |
| antisense | Firm1060R | AGCTGACGACAACCATGCAC | |
| Enterobacteriaceae [34] | sense | Uni515F | GTGCCAGCMGCCGCGGTAA |
| antisense | Ent826R | GCCTCAAGGGCACAACCTCCAAG | |
| Faecalibacterium prausnitzii [35] | sense | Fprau223F | GATGGCCTCGCGTCCGATTAG |
| antisense | Fprau420R | CCGAAGACCTTCTTCCTCC | |
| Lactobacillus murinus/animalis [36] | sense | TCGAACGAAACTTCTTTATCACC | |
| antisense | ATGACCCAGATCATGTTTGA | ||
| Lactobacillus helveticus | sense | ACCTGCCCCATAGTCTAGGA | |
| antisense | ACGCCGCCTTTTATAAGCTG | ||
| Lactobacillus gasseri | sense | AGACATGCGTCTAGTGTTGTT | |
| antisense | TGGGTAACCTGCCCAAGAGA | ||
| Genomic actin [37] | sense | ATGACCCAGATCATGTTTGA | |
| antisense | TACGACCAGAGGCATACAG | ||
| Fungi [38] | sense | ITS1-2 F | CTTGGTCATTTAGAGGAAGTAA |
| antisense | ITS1-2 R | GCTGCGTTCTTCATCGATGC | |
| Gene | 5’-3’Sequence | Sequence | Function |
|---|---|---|---|
| Alox15 | sense | GTTCAGGAACCACAGGGAGG | 12/15-Lipoxygenase |
| antisense | GTCAGAGATACTGGTCGCCG | enzyme involved in the synthesis of anti-inflammatory eicosanoids | |
| Alox5 | sense | AGAGCGGCAGCTCAGTTTAG | 5-Lipoxygenase |
| antisense | GGAACTGGTGTGTACAGGGG | enzyme involved in the synthesis of pro-inflammatory eicosanoids | |
| Arg1 | sense | CGTGTACATTGGCTTGCGAG | Arginase-1/anti-inflammatory marker |
| antisense | TCGGCCTTTTCTTCCTTCCC | /by degrading arginine, deprives NOS2 of its substrate | |
| Cat | sense | ACATGGTCTGGGACTTCTGG | Catalase-1 |
| antisense | CAAGTTTTTGATGCCCTGGT | antioxidant enzyme | |
| Ccl2 | sense | AGGTCCCTGTCATGCTTCTG | pro-inflammatory chemokine |
| antisense | TCTGGACCCATTCCTTCTTG | recruit monocytes to the site of inflammation | |
| Cd209b | sense | GGCACGAAAGTGAGGCACAT | SIGNR1/C-type lectin receptor |
| antisense | AGCTCATCTCCGCTCCTACCT | macrophage surface receptor | |
| Clec7a | sense | CCTCCAAGGCATCCCAAACT | Dectin-1/C-type lectin receptor |
| antisense | TAGCTGGGAGCAGTGTCTCT | macrophage surface receptor | |
| Crp | sense | CGCAGCTTCAGTGTCTTCTC | C reactive protein |
| antisense | AGATGTGTGTTGGAGCCTCA | inflammatory marker | |
| Fcgr3 | sense | TGTTTGCTTTTGCAGACAGG | CD16 Fcγ receptors |
| antisense | TGCTCCATTTGACACCGATA | macrophage surface receptor | |
| Fcgr1 | sense | GTTATTGCCACCAAGGCTGT | CD64 Fcγ receptors |
| antisense | ACCTGTATTCGTCACTGTCC | macrophage surface receptor | |
| Gapdh | sense | ACACATTGGGGGTAGGAACA | housekeeping |
| antisense | AACTTTGGCATTGTGGAAGG | ||
| Gp91phox | sense | ACTGCGGAGAGTTTGGAAGA | cytosolic subunit of the NADPH oxidase complex/reactive oxygen species release |
| antisense | GGTGATGACCACCTTTTGCT | ||
| Hpgds | sense | GGACACGCTGGATGACTTCA | Prostaglandin D synthase |
| antisense | TCCCAGTAGAAGTCTGCCCA | enzyme involved in the synthesis of anti-inflammatory eicosanoids | |
| Il10 | sense | AGGCGCTGTCATCGATTTCT | anti-inflammatory cytokine |
| antisense | GCTCCACTGCCTTGCTCTTA | ||
| Il12p40 | sense | AGGTCACACTGGACCAAAGG | pro-inflammatory cytokine |
| antisense | TGGTTTGATGATGTCCCTGA | ||
| Il1ra | sense | GGCCTAGGTGTCTTCTGCTC | Interleukin-1 receptor antagonist |
| antisense | GTAAGGGAGTCACTTGGGGC | anti-inflammatory marker | |
| Il1b | sense | CAACCAACAAGTGATATTCTCGATG | pro-inflammatory cytokine |
| antisense | GATCCACACTCTCCAGCTGCA | ||
| Il6 | sense | GAGGATACCACTCCCAACAGACC | pro-inflammatory cytokine |
| antisense | AAGTGCATCATCGTTGTTCATACA | ||
| Il8 | sense | TCCCTTGTGGAGGCTAGAGA | pro-inflammatory cytokine |
| antisense | AGGCACAGGTAGGATCC | ||
| Lta4h | sense | GTTGACAGCTGAACCCCAGT | LTB4 hydrolase critical to produce the pro-inflammatory mediator LTB4 |
| antisense | CGTGCCCTTAGTTCCACATT | ||
| Mrc1 | sense | GGGTTCACCTGGAGTGATGG | Mannose receptor/C-type lectin receptor |
| antisense | ATGCCAGGGTCACCTTTCAG | macrophage surface receptor | |
| Nos2 | sense | TCCTGGACATTACGACCCCT | Inducible Nitric oxide synthase |
| antisense | ACAAGGCCTCCAATCTCTGC | pro-inflammatory marker | |
| Pges | sense | CCTAGGCTTCAGCCTCACAC | Prostaglandin E synthase |
| antisense | CAGCCTATTGTTCAGCGACA | enzyme involved in the synthesis of pro-inflammatory eicosanoids | |
| Ptgs2 | sense | AGAAGGAAATGGCTGCAGAA | Cyclooxygenase-2 |
| antisense | GCTCGGCTTCCAGTATTGAG | enzyme involved in the synthesis of pro/anti-inflammatory eicosanoids | |
| p47phox (Ncf1) | sense | AGTGATGCGGAGACTTTGCT | cytosolic subunit of the NADPH oxidase complex/reactive oxygen species release |
| antisense | ACCGGAGTTACAGGCAAATG | ||
| Sod2 | sense | GCCCCCTGAGTTGTTGAATA | Superoxide dismutase-2 |
| antisense | AGACAGGCAAGGCTCTACCA | antioxidant enzyme | |
| Tgfb1 | sense | AGGTTGGCATTCCACTTCAC | anti-inflammatory cytokine |
| antisense | AGGGGCCTCTAAGAGCAGTC | ||
| Tlr2 | sense | TGCTTTCCTGCTGGAGATTT | Toll like receptor-2 |
| antisense | TGTAACGCAACAGCTTCAGG | macrophage surface receptor | |
| Tnfa | sense | AGCCCCCAGTCTGTATCCTT | pro-inflammatory cytokine |
| antisense | CTCCCTTTGCAGAACTCAGG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Authier, H.; Salon, M.; Rahabi, M.; Bertrand, B.; Blondeau, C.; Kuylle, S.; Holowacz, S.; Coste, A. Oral Administration of Lactobacillus helveticus LA401 and Lactobacillus gasseri LA806 Combination Attenuates Oesophageal and Gastrointestinal Candidiasis and Consequent Gut Inflammation in Mice. J. Fungi 2021, 7, 57. https://doi.org/10.3390/jof7010057
Authier H, Salon M, Rahabi M, Bertrand B, Blondeau C, Kuylle S, Holowacz S, Coste A. Oral Administration of Lactobacillus helveticus LA401 and Lactobacillus gasseri LA806 Combination Attenuates Oesophageal and Gastrointestinal Candidiasis and Consequent Gut Inflammation in Mice. Journal of Fungi. 2021; 7(1):57. https://doi.org/10.3390/jof7010057
Chicago/Turabian StyleAuthier, Hélène, Marie Salon, Mouna Rahabi, Bénédicte Bertrand, Claude Blondeau, Sarah Kuylle, Sophie Holowacz, and Agnès Coste. 2021. "Oral Administration of Lactobacillus helveticus LA401 and Lactobacillus gasseri LA806 Combination Attenuates Oesophageal and Gastrointestinal Candidiasis and Consequent Gut Inflammation in Mice" Journal of Fungi 7, no. 1: 57. https://doi.org/10.3390/jof7010057
APA StyleAuthier, H., Salon, M., Rahabi, M., Bertrand, B., Blondeau, C., Kuylle, S., Holowacz, S., & Coste, A. (2021). Oral Administration of Lactobacillus helveticus LA401 and Lactobacillus gasseri LA806 Combination Attenuates Oesophageal and Gastrointestinal Candidiasis and Consequent Gut Inflammation in Mice. Journal of Fungi, 7(1), 57. https://doi.org/10.3390/jof7010057