
Identification of Fungal Pathogens to Control Postharvest Passion Fruit (Passiflora edulis) Decays and Multi-Omics Comparative Pathway Analysis Reveals Purple Is More Resistant to Pathogens than a Yellow Cultivar

 , ,
, ,  , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fruit Sample Collection
2.2. Fungal Pathogens Isolation
2.3. Morphological Identification of Fungal Isolates
2.4. Molecular Identification of Fungal Isolates
2.5. Phylogenetic Analysis
2.6. Pathogenicity Test
2.7. Phytohormones Analysis
2.8. RNA Extraction, cDNA Library Construction, and Transcriptome Sequencing
2.9. Gene Function Annotation, Expression, and Pathway Analysis
2.10. Non-Target Metabolite Analysis by LC-ESI-Q-ToF-MS
2.11. Statistical Analysis
3. Results
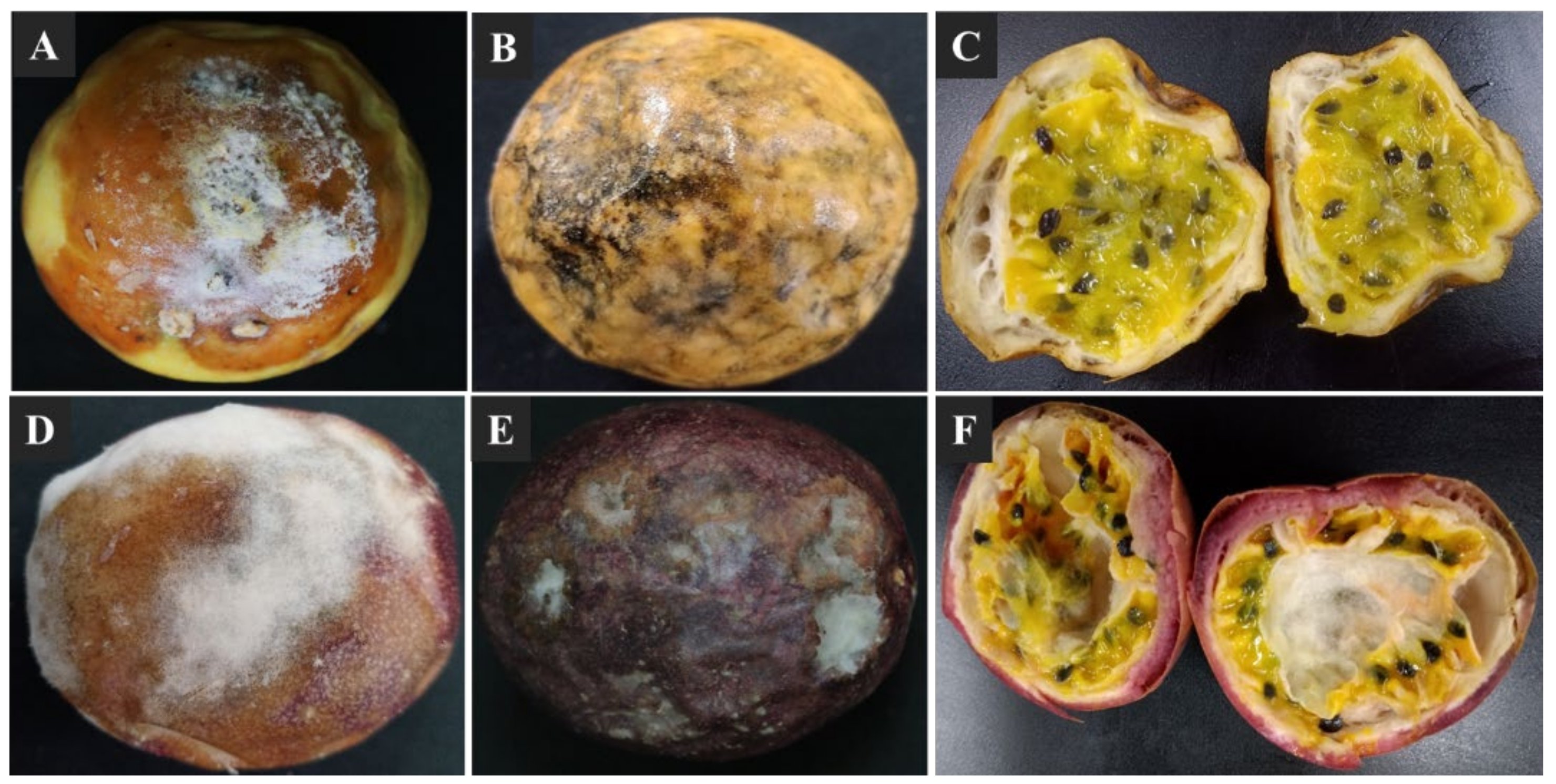
3.1. Postharvest Decay Symptoms in Passion Fruits
3.2. Fungus Isolates from Infected Passion Fruit
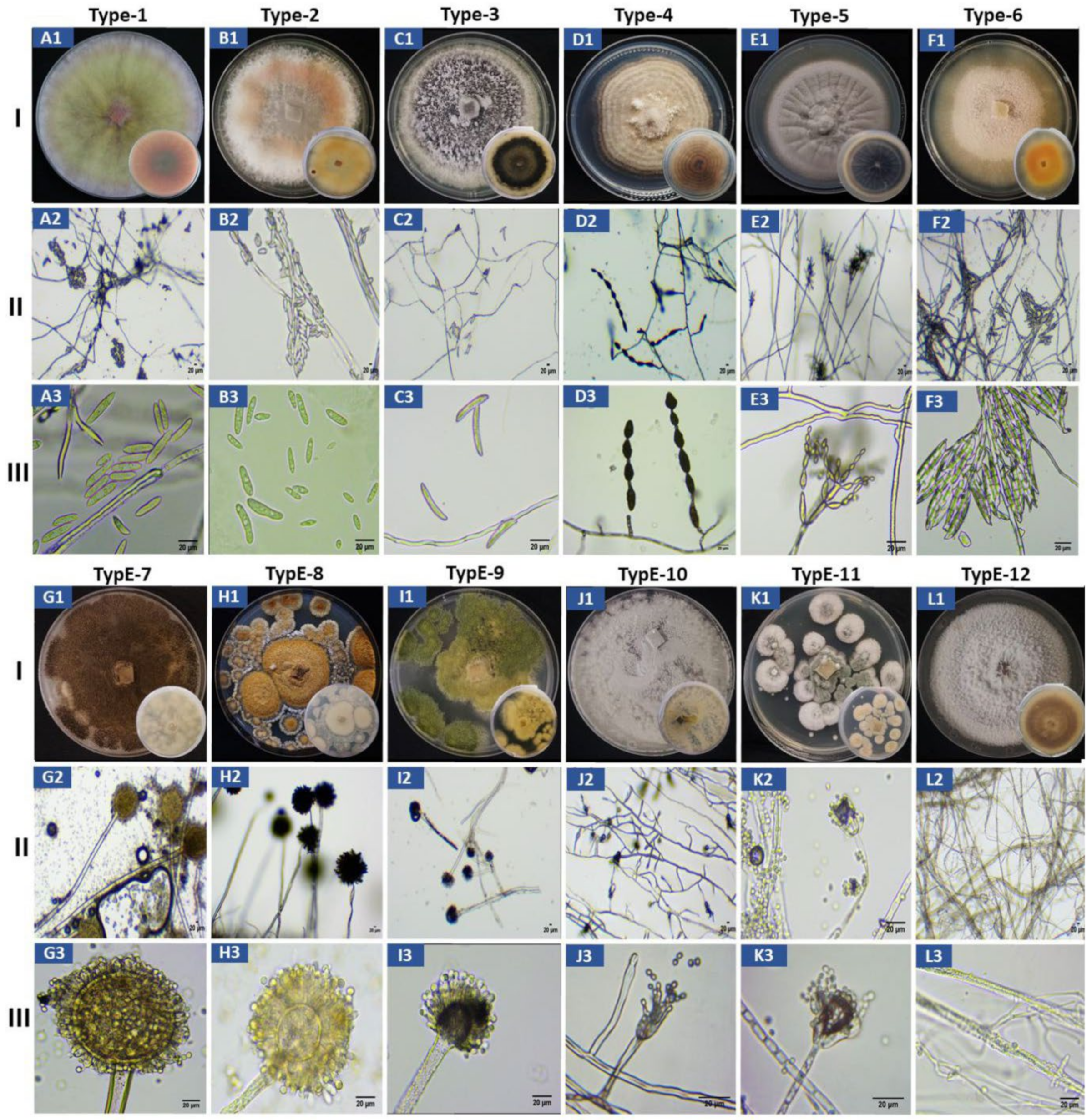
3.3. Morphological Characterization of the 12 Morphotypes
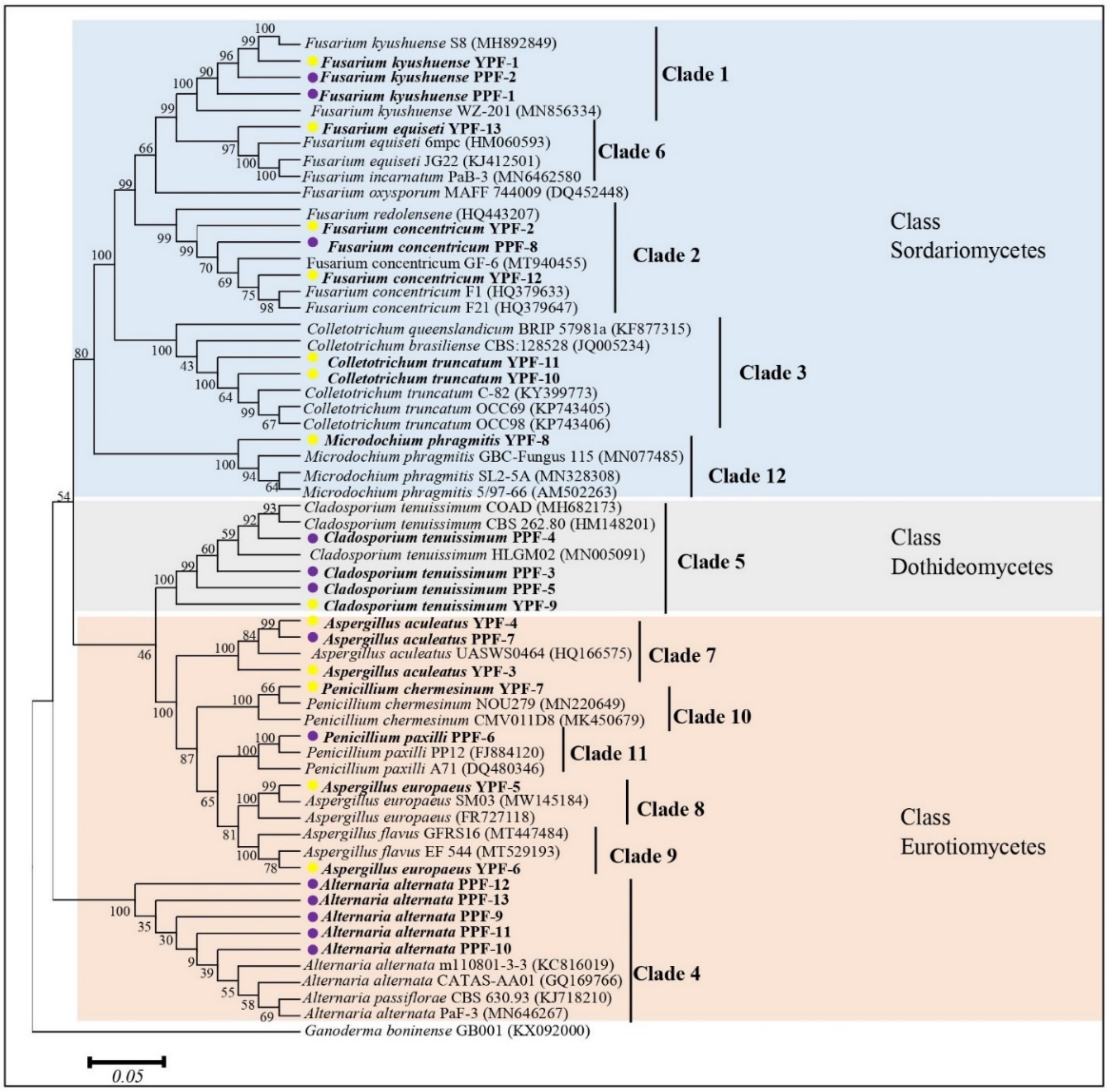
3.4. Molecular Characterization and Phylogenetic Analysis
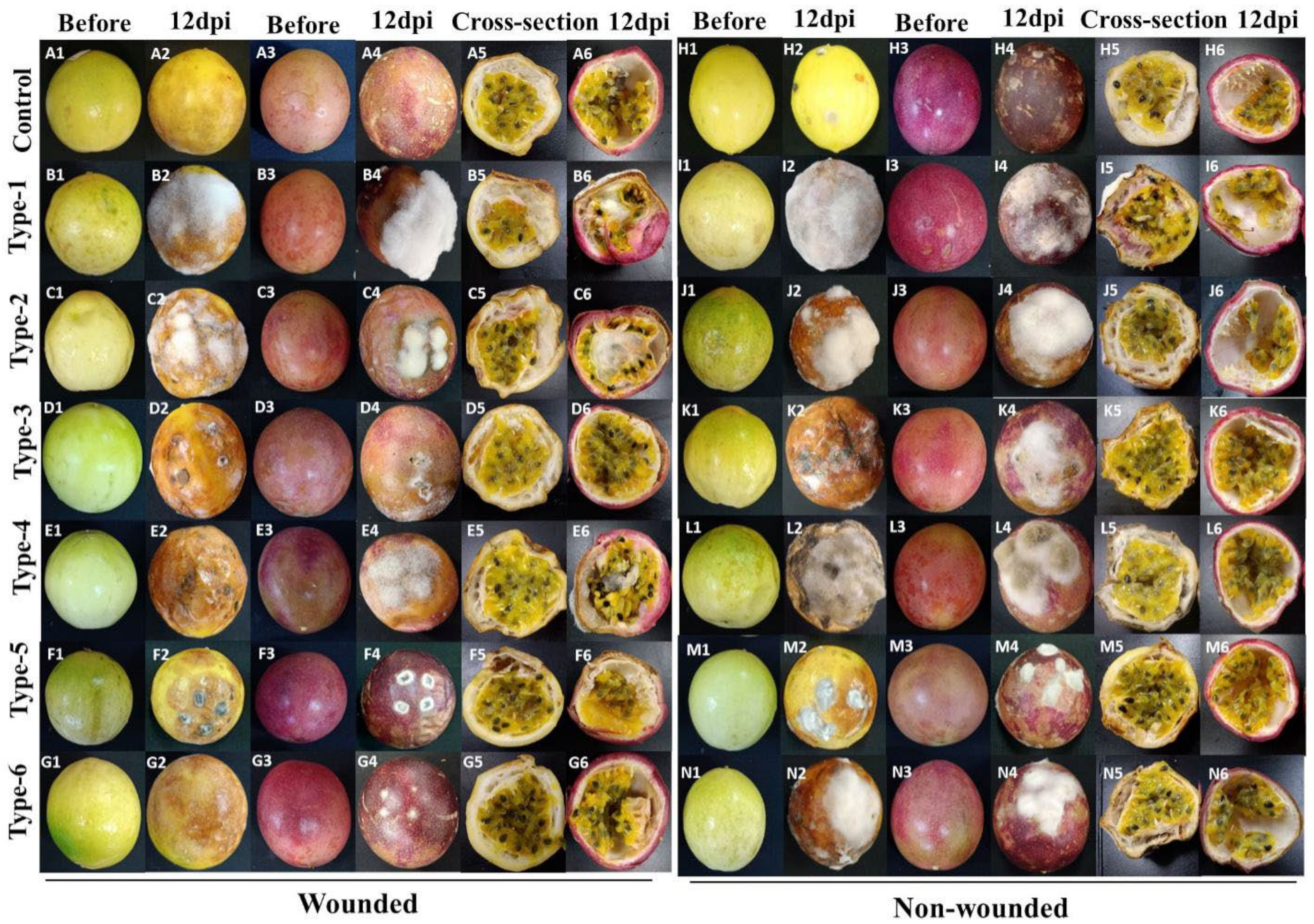
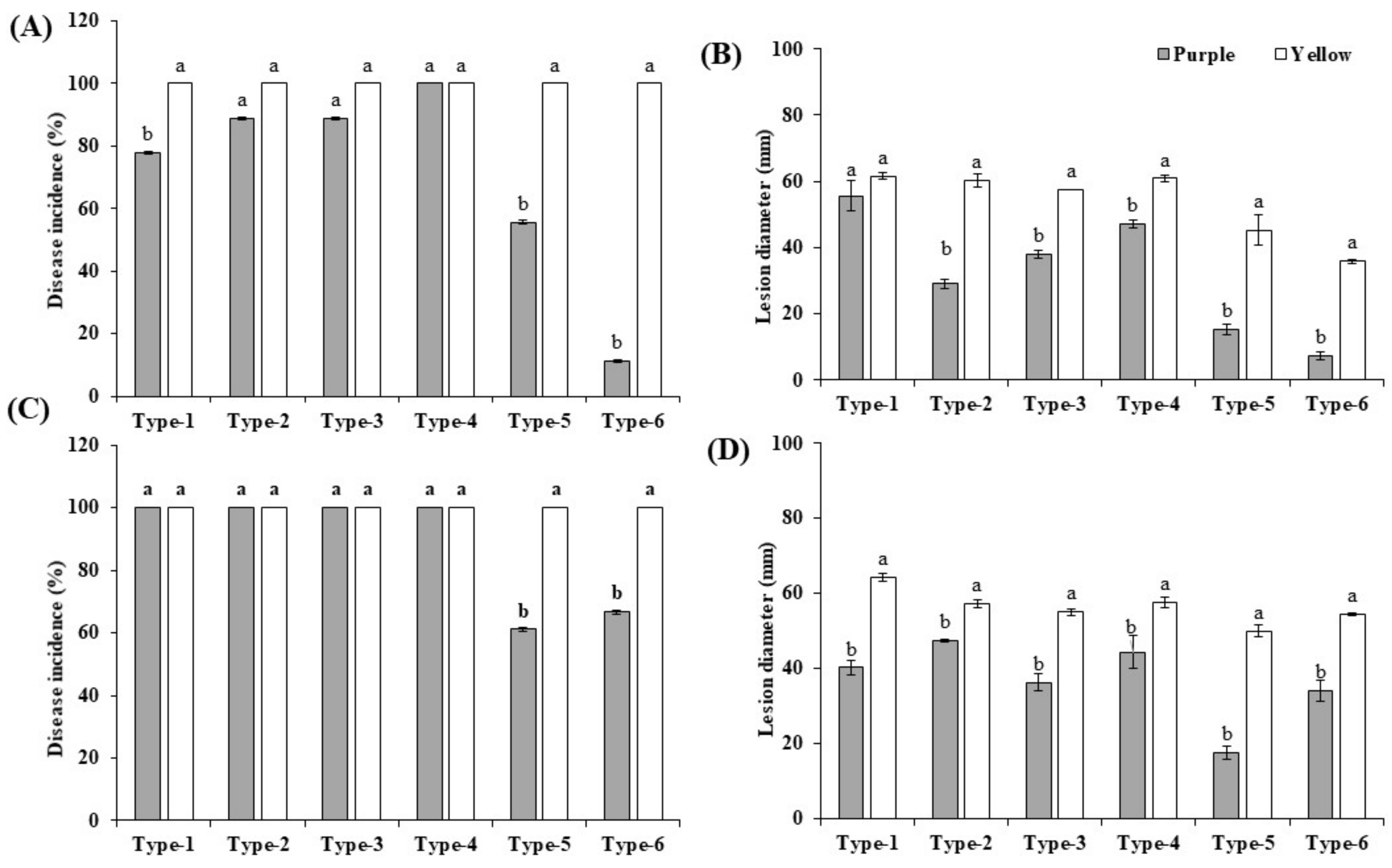
3.5. Pathogenicity Tests
3.6. Phytohormones Levels in the Peels of the Fruit of the Yellow and Purple Cultivars
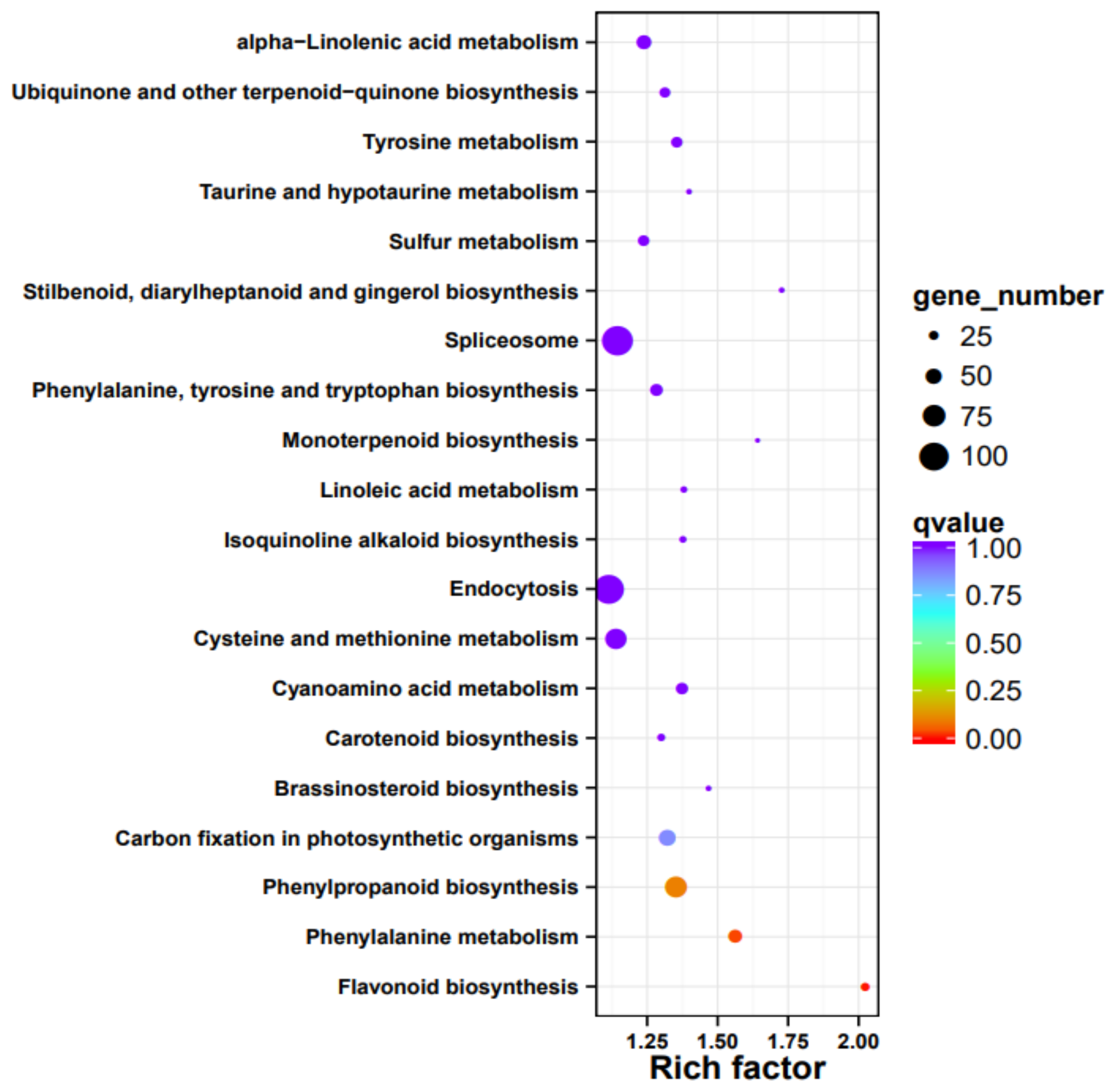
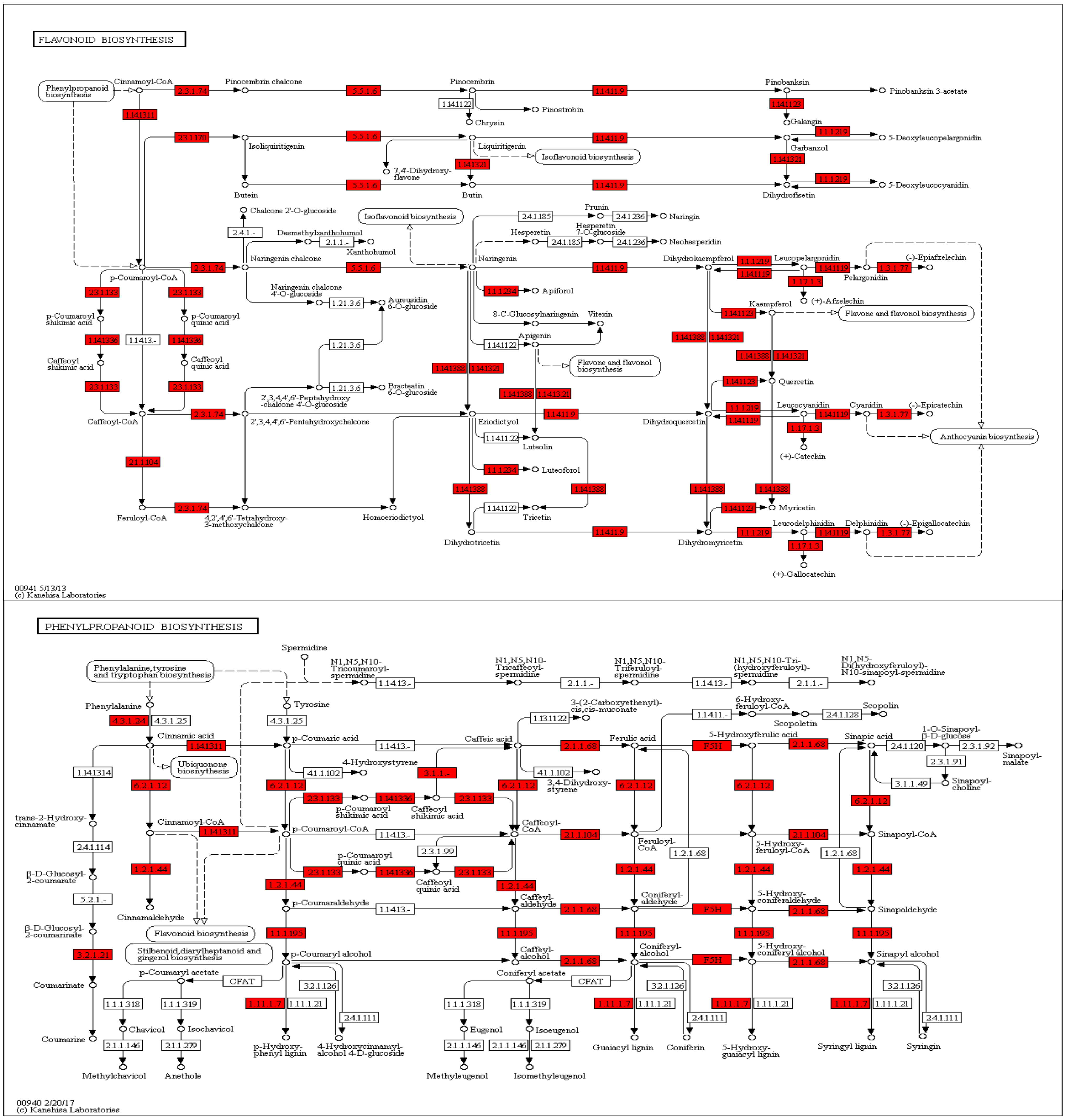
3.7. Gene Function Annotation, Expression, and Pathway Analysis
3.8. Differences in the Metabolite Profiles of Purple and Yellow Peels
4. Discussion
4.1. Comparison of the Fruit of the Yellow and Purple Cultivar
4.2. Hormones
4.3. Expression and Metabolite Profiles
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- De Toledo, N.M.V.; De Camargo, A.C.; Ramos, P.B.M.; Button, D.C.; Granato, D.; Canniatti-Brazaca, S.G. Potentials and Pitfalls on the Use of Passion Fruit By-Products in Drinkable Yogurt: Physicochemical, Technological, Microbiological, and Sensory Aspects. Beverages 2018, 4, 47. [Google Scholar] [CrossRef] [Green Version]
- FAO. Minor Tropical Fruits; Food and Agriculture Organization of United Nation: Rome, Italy, 2018; Available online: http://www.fao.org/fileadmin/templates/est/COMM_MARKETS_MONITORING/Tropical_Fruits/Documents/Minor_Tropical_Fruits_FoodOutlook_1_2018.pdf (accessed on 11 October 2020).
- Antognoni, F.; Zheng, S.; Pagnucco, C.; Baraldi, R.; Poli, F.; Biondi, S. Induction of flavonoid production by UV-B radiation in Passiflora quadrangularis callus cultures. Fitoterapia 2007, 78, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Zeraik, M.L.; Yariwake, J. Quantification of isoorientin and total flavonoids in Passiflora edulis fruit pulp by HPLC-UV/DAD. Microchem. J. 2010, 96, 86–91. [Google Scholar] [CrossRef]
- Liu, S.; Li, A.; Chen, C.; Cai, G.; Zhang, L.; Guo, C.; Xu, M. De Novo Transcriptome Sequencing in Passiflora edulis Sims to Identify Genes and Signaling Pathways Involved in Cold Tolerance. Forests 2017, 8, 435. [Google Scholar] [CrossRef] [Green Version]
- Brasil, I.M.; Siddiqui, M.W. Postharvest quality of fruits and vegetables: An overview. In Preharvest Modulation of Postharvest Fruit and Vegetable Quality; Elsevier: Cambridge, MA, USA, 2018; pp. 1–40. [Google Scholar]
- Aulakh, J.; Regmi, A.; Fulton, J.R.; Alexander, C.E. Estimating Post-Harvest Food Losses: Developing A Consistent Global Estimation Framework; Joint Annual Meeting: Washington, DC, USA, 2013. [Google Scholar]
- Dutra, J.B.; Blum, L.E.B.; Lopes, L.F.; Cruz, A.F.; Uesugi, C.H. Use of hot water, combination of hot water and phosphite, and 1-MCP as post-harvest treatments for passion fruit (Passiflora edulis f. flavicarpa) reduces anthracnose and does not alter fruit quality. Hortic. Environ. Biotechnol. 2018, 59, 847–856. [Google Scholar] [CrossRef]
- Gustavsson, J.; Cederberg, C.; Sonesson, U.; Van Otterdijk, R.; Meybeck, A. Global Food Losses and Food Waste; FAO: Rome, Italy, 2011. [Google Scholar]
- Alkan, N.; Fortes, A.M. Insights into molecular and metabolic events associated with fruit response to post-harvest fungal pathogens. Front. Plant Sci. 2015, 6, 889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez, M.F.; Contreras, L.; Garnica, N.M.; Fernández-Zenoff, M.V.; Farias, M.E.; Sepulveda, M.; Ramallo, J.; Dib, J.R. Native Killer Yeasts as Biocontrol Agents of Postharvest Fungal Diseases in Lemons. PLoS ONE 2016, 11, e0165590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palou, L.; Montesinos-Herrero, C.; Tarazona, I.; Besada, C.; Taberner, V. Incidence and Etiology of Postharvest Fungal Diseases of Persimmon (Diospyros kaki Thunb. cv. Rojo Brillante) in Spain. Plant Dis. 2015, 99, 1416–1425. [Google Scholar] [CrossRef] [PubMed]
- Kamle, M.; Kumar, P.; Gupta, V.K.; Tiwari, A.K.; Misra, A.K.; Pandey, B.K. Identification and phylogenetic correlation among Colletotrichum gloeosporioides pathogen of anthracnose for mango. Biocatal. Agric. Biotechnol. 2013, 2, 285–287. [Google Scholar] [CrossRef]
- Damm, U.; Sato, T.; Alizadeh, A.; Groenewald, J.Z.; Crous, P.W. The Colletotrichum dracaenophilum, C. ámagnum and C. áorchidearum species complexes. Stud. Mycol. 2019, 92, 1–46. [Google Scholar] [CrossRef]
- Qiu, F.; Li, X.; Xie, C.P.; Li, J.; Zheng, F.Q. Identification of Colletotrichum brevisporum causing fruit rot in yellow passion fruit (Passiflora edulis f. flavicarpa) in China. Australas. Plant Pathol. 2021, 50, 229–232. [Google Scholar] [CrossRef]
- Peres, N.A.R.; Kuramae, E.; Dias, M.S.C.; De Souza, N.L. Identification and Characterization of Colletotrichum spp. affecting Fruit after Harvest in Brazil. J. Phytopathol. 2002, 150, 128–134. [Google Scholar] [CrossRef]
- Zhang, W.; Niu, X.L.; Yang, J.Y. First Report of Postharvest Fruit Rot on Passion Fruit (Passiflora edulis) Caused by Lasiodiplodia theobromae in Mainland China. Plant Dis. 2021, 105, 1198. [Google Scholar] [CrossRef]
- Muniz, M.; Rocha, D.F.; Silveira, N.S.S.; Menezes, M. Identification of fungi causal agents of postharvest diseases on commercialized fruits in Alagoas, Brazil. Summa Phytopathol. 2003, 29, 38–42. [Google Scholar]
- Gil, J.G.R.; Tamayo, P.J.; Morales, J.G. Identification and pathogenicity of microorganisms affecting purple passion fruit in Colombia. Rev. Ceres 2017, 64, 250–257. [Google Scholar] [CrossRef] [Green Version]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W. Fungal Barcoding Consortium. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raja, H.A.; Miller, A.N.; Pearce, C.J.; Oberlies, N.H. Fungal Identification Using Molecular Tools: A Primer for the Natural Products Research Community. J. Nat. Prod. 2017, 80, 756–770. [Google Scholar] [CrossRef] [PubMed]
- Harsonowati, W.; Marian, M.; Surono Narisawa, K. The Effectiveness of a Dark Septate Endophytic Fungus, Cladophialophora chaetospira SK51, to Mitigate Strawberry Fusarium Wilt Disease and with Growth Promotion Activities. Front. Microbiol. 2020, 11, 585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manter, D.K.; Vivanco, J.M. Use of the ITS primers, ITS1F and ITS4, to characterize fungal abundance and diversity in mixed-template samples by qPCR and length heterogeneity analysis. J. Microbiol. Methods 2007, 71, 7–14. [Google Scholar] [CrossRef]
- Zarrin, M.; Ganj, F.; Faramarzi, S. Analysis of the rDNA internal transcribed spacer region of the Fusarium species by polymerase chain reaction-restriction fragment length polymorphism. Biomed. Rep. 2016, 4, 471–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M., Gelfand, D., Sninsky, J., White, T., Eds.; Academic Press Inc.: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Hu, Y.; Jiao, L.; Jiang, M.-H.; Yin, S.; Dong, P.; Zhao, Z.-M.; Yang, D.-P.; Ho, P.-T.; Wang, D.-M. A new C-glycosyl flavone and a new neolignan glycoside from Passiflora edulis Sims peel. Nat. Prod. Res. 2018, 32, 2312–2318. [Google Scholar] [CrossRef]
- Farag, M.A.; Otify, A.; Porzel, A.; Michel, C.G.; Elsayed, A.; Wessjohann, L.A. Comparative metabolite profiling and fingerprinting of genus Passiflora leaves using a multiplex approach of UPLC-MS and NMR analyzed by chemometric tools. Anal. Bioanal. Chem. 2016, 408, 3125–3143. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Luan, F.; Yang, Y.; Wang, Z.; Zhao, Z.; Fang, J.; Wang, M.; Zuo, M.; Li, Y. Passiflora edulis: An Insight Into Current Researches on Phytochemistry and Pharmacology. Front. Pharmacol. 2020, 11, 617. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Xin, M.; Li, L.; He, X.; Liu, G.; Li, J.; Sheng, J.; Sun, J. Transcriptome profiling helps to elucidate the mechanisms of ripening and epidermal senescence in passion fruit (Passiflora edulia Sims). PLoS ONE 2020, 15, e0236535. [Google Scholar] [CrossRef]
- Yan, C.; Rizwan, H.M.; Liang, D.; Reichelt, M.; Mithöfer, A.; Scholz, S.S.; Oelmüller, R.; Chen, F. The effect of the root-colonizing Piriformospora indica on passion fruit (Passiflora edulis) development: Initial defense shifts to fitness benefits and higher fruit quality. Food Chem. 2021, 359, 129671. [Google Scholar] [CrossRef] [PubMed]
- Tziros, G.; Tzavella-Klonari, K. Pomegranate fruit rot caused by Coniella granati confirmed in Greece. Plant Pathol. 2008, 57, 783. [Google Scholar]
- Li, X.; Lu, X.; He, Y.; Deng, M.; Lv, Y. Identification the Pathogens Causing Rot Disease in Pomegranate (Punica granatum L.) in China and the Antifungal Activity of Aqueous Garlic Extract. Forests 2019, 11, 34. [Google Scholar] [CrossRef] [Green Version]
- Thrane, U.; Adler, A.; Clasen, P.-E.; Galvano, F.; Langseth, W.; Lew, H.; Logrieco, A.F.; Nielsen, K.F.; Ritieni, A. Diversity in metabolite production by Fusarium langsethiae, Fusarium poae, and Fusarium sporotrichioides. Int. J. Food Microbiol. 2004, 95, 257–266. [Google Scholar] [CrossRef]
- Zurlo, D.; Assante, G.; Moricca, S.; Colantuoni, V.; Lupo, A. Cladosporol A, a new peroxisome proliferator-activated receptor γ (PPARγ) ligand, inhibits colorectal cancer cells proliferation through β-catenin/TCF pathway inactivation. Biochim. Biophys. Acta (BBA)—Gen. Subj. 2014, 1840, 2361–2372. [Google Scholar] [CrossRef]
- Shi, W.; Tan, Y.; Wang, S.; Gardiner, D.M.; De Saeger, S.; Liao, Y.; Wang, C.; Fan, Y.; Wang, Z.; Wu, A. Mycotoxigenic Potentials of Fusarium Species in Various Culture Matrices Revealed by Mycotoxin Profiling. Toxins 2016, 9, 6. [Google Scholar] [CrossRef] [Green Version]
- Mishra, R.; Mohanty, J.N.; Chand, S.K.; Joshi, R.K. Can-miRn37a mediated suppression of ethylene response factors enhances the resistance of chilli against anthracnose pathogen Colletotrichum truncatum L. Plant Sci. 2018, 267, 135–147. [Google Scholar] [CrossRef] [PubMed]
- Kojima, H.; Hossain, M.; Kubota, M.; Hyakumachi, M. Involvement of the salicylic acid signaling pathway in the systemic resistance induced in Arabidopsis by plant growth-promoting fungus Fusarium equiseti GF19-1. J. Oleo Sci. 2013, 62, 415–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Han, S.; Wang, G.; Liu, X.; Amombo, E.; Xie, Y.; Fu, J. The Fungus Aspergillus aculeatus Enhances Salt-Stress Tolerance, Metabolite Accumulation, and Improves Forage Quality in Perennial Ryegrass. Front. Microbiol. 2017, 8, 1664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Zhao, C.; Zhang, T.; Wang, G.; Amombo, E.; Xie, Y.; Fu, J. Exogenous Aspergillus aculeatus Enhances Drought and Heat Tolerance of Perennial Ryegrass. Front. Microbiol. 2021, 12, 307. [Google Scholar]
- Lohmar, J.M.; Puel, O.; Cary, J.W.; Calvo, A.M. The Aspergillus flavus rtfA Gene Regulates Plant and Animal Pathogenesis and Secondary Metabolism. Appl. Environ. Microbiol. 2019, 85, e02446-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toghueo, R.M.K.; Boyom, F.F. Endophytic Penicillium species and their agricultural, biotechnological, and pharmaceutical applications. 3 Biotech 2020, 10, 107. [Google Scholar] [CrossRef]
- Ernst, M.; Neubert, K.; Mendgen, K.W.; Wirsel, S.G. Niche differentiation of two sympatric species of Microdochium colonizing the roots of common reed. BMC Microbiol. 2011, 11, 242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerqueira-Silva, C.B.M.; Jesus, O.N.; Oliveira, E.; Santos, E.S.L.; Souza, A.P. Characterization and selection of passion fruit (yellow and purple) accessions based on molecular markers and disease reactions for use in breeding programs. Euphytica 2015, 202, 345–359. [Google Scholar] [CrossRef] [Green Version]
- Duthie, G.G.; Wood, A.D. Natural salicylates: Foods, functions and disease prevention. Food Funct. 2011, 2, 515–520. [Google Scholar] [CrossRef]
- Maynard, D.; Gröger, H.; Dierks, T.; Dietz, K.-J. The function of the oxylipin 12-oxophytodienoic acid in cell signaling, stress acclimation, and development. J. Exp. Bot. 2018, 69, 5341–5354. [Google Scholar] [CrossRef]
- Dave, A.; Graham, I.A. Oxylipin Signaling: A Distinct Role for the Jasmonic Acid Precursor cis-(+)-12-Oxo-Phytodienoic Acid (cis-OPDA). Front. Plant Sci. 2012, 3, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Yu, H.; Rao, X.; Li, L.; Dixon, R.A. Abscisic acid regulates secondary cell-wall formation and lignin deposition in Arabidopsis thaliana through phosphorylation of NST1. Proc. Natl. Acad. Sci. USA 2021, 118, e2010911118. [Google Scholar]
- Moya-León, M.A.; Mattus-Araya, E.; Herrera, R. Molecular Events Occurring During Softening of Strawberry Fruit. Front. Plant Sci. 2019, 10, 615. [Google Scholar] [CrossRef] [PubMed]
- Forlani, S.; Masiero, S.; Mizzotti, C. Fruit ripening: The role of hormones, cell wall modifications, and their relationship with pathogens. J. Exp. Bot. 2019, 70, 2993–3006. [Google Scholar] [CrossRef]
- Leng, P.; Yuan, B.; Guo, Y. The role of abscisic acid in fruit ripening and responses to abiotic stress. J. Exp. Bot. 2013, 65, 4577–4588. [Google Scholar] [CrossRef] [PubMed]
- Zaynab, M.; Fatima, M.; Abbas, S.; Sharif, Y.; Umair, M.; Zafar, M.H.; Bahadar, K. Role of secondary metabolites in plant defense against pathogens. Microb. Pathog. 2018, 124, 198–202. [Google Scholar] [CrossRef] [PubMed]
- Zozio, S.; Servent, A.; Cazal, G.; Mbéguié-A-Mbéguié, D.; Ravion, S.; Pallet, D.; Abel, H. Changes in antioxidant activity during the ripening of jujube (Ziziphus mauritiana Lamk). Food Chem. 2014, 150, 448–456. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Morphotype | Number of Isolates | Passion Fruit Cultivar | Identified Species |
|---|---|---|---|
| 1 | 3 | yellow and purple | Fusarium kyushuense |
| 2 | 3 | yellow and purple | Fusarium concentricum |
| 3 | 2 | yellow | Colletotrichum truncatum |
| 4 | 5 | purple | Alternaria alternata |
| 5 | 4 | yellow and purple | Cladosporium tenuissimum |
| 6 | 1 | yellow | Fusarium equiseti |
| 7 | 3 | yellow and purple | Aspergillus aculeatus |
| 8 | 1 | yellow | Aspergillus europaeus |
| 9 | 1 | yellow | Aspergillus flavus |
| 10 | 1 | yellow | Penicillium chermesinum |
| 11 | 1 | purple | Penicillium paxilli |
| 12 | 1 | yellow | Microdochium phragmitis |
| Morphotype | Colony on PDA Media | Conidia | ||
|---|---|---|---|---|
| Morphology | Growth Rate (mm/Week) | Length (µm) | Shape | |
| Type-1 | Reddish-white and floccose mycelia with deep red pigmentation on the agar side. | 60–65 | 20–25 | Obovate, ellipsoidal to clavate and 3–5 septate in macro-conidia. |
| Type-2 | Reddish to white with lavish cottony mycelia. White to pale yellow pigmentation on the agar side. | 50–55 | 26–45 | Oval, obovoid to allantoid and macro-conidia slender with 3–5 septate. |
| Type-3 | White grayish to dark grey, light to dark reverse pigmentation. | 55–60 | 21–27 | Non-septate, hyaline, falcate, and truncate. |
| Type-4 | White to gray at the edge and olivaceous buff in the center. | 55–60 | 25–56 | Chains, obclavate, ovoid or ellipsoid and three to seven transverse septa. |
| Type-5 | Olive-brown to dark green with gray-olivaceous to white edges, velvet-like texture with radially furrowed, dark pigmentation on the agar side. | 35–40 | 7–12 | Smooth, single-celled, olive-brown, elliptical to limonifor. |
| Type-6 | Lavish white and fluffy aerial mycelium with dark to pale brown from front and pigments on the agar side. | 27–35 | 13–34 | Macro-conidia with mostly three to five-septae, slightly curved to lunate at apex. |
| Type-7 | Dark brown to black colonies with rough texture, white mycelia underneath the colonies. Whitish yellow radial furrows at the backside. | 30–35 | 4–5 | Dark brown to black conidia, ellipsoidal, phialides, spinose, borne in radiate heads. |
| Type-8 | Plane colonies, floccose from center with strong sporulation, no soluble pigment, and light olive on the agar side. | 20–25 | 3–4.5 | Globose conidia, roughened and yellow-brown to brown at maturity. |
| Type-9 | Plain and flat at the edges, raised at the center and wrinkled cerebriform pattern, produce greenish conidia with a white border, cream color on the agar side. | 20–25 | 3–6 | Conidia with a thick mycelial mat, globose shape, thin walls and rough texture. Metulae obscured on the entire surface of the vesicles. |
| Type-10 | Fast growing, white green-gray shade, dense conidiophores, and non-circular growth. | 22–25 | 2.5–4 | Basocatenate, hyaline or greenish, globose, ellipsoidal, cylindrical or fusiform, and smooth or rough-walled. |
| Type-11 | Fast growing, white to light green shade, dense conidiophores with white edges and irregular growth. | 18–25 | 2.5–4 | Hyaline or greenish, globose, ellipsoidal, cylindrical, or fusiform. |
| Type-12 | Pinkish white flat colonies, entire margins, slightly raised to umbonate center and greyish orange on the agar side. | 35–40 | - | No conidia produced in lab condition. |
| Isolates | Morpho-Type | Accession Numbers a | Fungal Taxon | Query Cover (%) | Seq. Similarity (%) | Accession Numbers of ITS-rDNA b |
|---|---|---|---|---|---|---|
| YPF-1 | Type-1 | MW880893 | Fusarium kyushuense | 96 | 100 | KC466546 |
| PPF-1 | Type-1 | MW880906 | Fusarium kyushuense | 100 | 100 | KC466546 |
| PPF-2 | Type-1 | MW880907 | Fusarium kyushuense | 100 | 100 | KC466546 |
| YPF-2 | Type-2 | MW880894 | Fusarium concentricum | 100 | 99 | MN341308 |
| YPF-12 | Type-2 | MW880904 | Fusarium concentricum | 100 | 100 | LC317601 |
| PPF-8 | Type-2 | MW880913 | Fusarium concentricum | 100 | 100 | LC317601 |
| YPF-10 | Type-3 | MW880902 | Colletotrichum truncatum | 100 | 100 | JQ936246 |
| YPF-11 | Type-3 | MW880903 | Colletotrichum truncatum | 100 | 100 | JQ936246 |
| PPF-9 | Type-4 | MW880914 | Alternaria alternata | 100 | 100 | MN547372 |
| PPF-10 | Type-4 | MW880915 | Alternaria alternata | 100 | 100 | MN547372 |
| PPF-11 | Type-4 | MW880916 | Alternaria alternata | 100 | 100 | MT482506 |
| PPF-12 | Type-4 | MW880917 | Alternaria alternata | 100 | 100 | MN547372 |
| PPF-13 | Type-4 | MW880918 | Alternaria alternata | 100 | 100 | MN547372 |
| YPF-9 | Type-5 | MW880901 | Cladosporium tenuissimum | 100 | 100 | MF422152 |
| PPF-3 | Type-5 | MW880908 | Cladosporium tenuissimum | 100 | 100 | MF422152 |
| PPF-4 | Type-5 | MW880909 | Cladosporium tenuissimum | 100 | 100 | MF422152 |
| PPF-5 | Type-5 | MW880910 | Cladosporium tenuissimum | 100 | 100 | MF422152 |
| YPF-13 | Type-6 | MW880905 | Fusarium equiseti | 100 | 100 | KR364600 |
| YPF-3 | Type-7 | MW880895 | Aspergillus aculeatus | 99 | 99 | KU203321 |
| YPF-4 | Type-7 | MW880896 | Aspergillus aculeatus | 100 | 100 | EU645733 |
| PPF-7 | Type-7 | MW880912 | Aspergillus aculeatus | 100 | 100 | LC514695 |
| YPF-5 | Type-8 | MW880897 | Aspergillus europaeus | 100 | 100 | FR727118 |
| YPF-6 | Type-9 | MW880898 | Aspergillus flavus | 100 | 100 | MG228413 |
| YPF-7 | Type-10 | MW880899 | Penicillium chermesinum | 100 | 100 | MK450679 |
| PPF-6 | Type-11 | MW880911 | Penicillium paxilli | 100 | 98 | AB933278 |
| YPF-8 | Type-12 | MW880900 | Microdochium phragmitis | 99 | 97 | AM502263 |
| Fruit Peels of Cultivar | SA (µg kg−1) | JA (µg kg−1) | JA-Ile (µg kg−1) | cis-OPDA (µg kg−1) | ABA (µg kg−1) |
|---|---|---|---|---|---|
| Purple | 504 ± 81 | 34 ± 4 | 1.3± 0.3 | 3.1± 0.4 | 1133 ± 56 |
| Yellow | 449 ± 73 | 24 ± 4 | 2.2± 0.4 | 21.9 ± 3 | 2644 ± 94 |
| Unigenes | Log2 Purple vs. Yellow | Gene Annotation |
|---|---|---|
| 060661 | 3.2 | ABC transporter C family member |
| 073495 | 3.2 | PLAT domain-containing protein |
| 012452 | 3.2 | leucoanthocyanidin reductase |
| 066444 | 3.2 | ABC transporter G family member |
| 078965 | 3.2 | Toll-like receptor |
| 079007 | 3.2 | ABC transporter B family member |
| 157353 | 3.2 | glutathione S-transferase |
| 075520 | 3.3 | disease resistance protein |
| 066058 | 3.3 | AP2-like ethylene-responsive transcription factor |
| 014357 | 3.3 | disease resistance protein |
| 002704 | 3.5 | RALF protein |
| 075276 | 3.5 | ethylene-responsive transcription factor |
| 019034 | 3.6 | LRR receptor-like serine/threonine-protein kinase |
| 080936 | 3.6 | LRR receptor-like serine/threonine-protein kinase |
| 115445 | 3.6 | cellulose synthase subunit |
| 006373 | 3.6 | allene oxide synthase |
| 079086 | 3.6 | ethylene-responsive transcription factor |
| 013300 | 3.7 | detoxification protein |
| 072633 | 3.7 | MLO-like protein |
| 058972 | 3.7 | ent-kaurene oxidase |
| 065344 | 3.7 | callose synthase |
| 012846 | 3.7 | disease resistance protein |
| 077385 | 3.7 | ethylene-responsive transcription factor |
| 013346 | 3.8 | salicylate carboxymethyltransferase |
| 066699 | 3.9 | mechanosensitive ion channel |
| 081472 | 3.9 | disease-resistance receptor-like protein kinase |
| 045952 | 3.9 | pathogenesis-related protein |
| 081029 | 3.9 | disease resistance protein |
| 074186 | 3.9 | detoxification protein |
| 092433 | 3.9 | ethylene-responsive transcription factor |
| 076826 | 4.0 | ethylene-responsive transcription factor |
| 029255 | 4.0 | remorin |
| 099452 | 4.1 | PLAT domain-containing protein |
| 034958 | 4.1 | flavonol synthase |
| 005394 | 4.1 | detoxification protein |
| 058910 | 4.2 | elicitor-responsive protein |
| 151621 | 4.2 | 1-amino-cyclopropane-1-carboxylic acid oxidase |
| 077400 | 4.2 | respiratory burst oxidase homolog protein C |
| 078602 | 4.3 | Downy Mildew Resistance protein |
| 080595 | 4.3 | TMV resistance protein |
| 153053 | 4.3 | remorin |
| 153975 | 4.3 | 1-aminocyclopropane-1-carboxylate oxidase |
| 051447 | 4.5 | ethylene-responsive transcription factor |
| 073788 | 4.6 | leucoanthocyanidin reductase |
| 135950 | 4.7 | pathogenesis-related genes transcriptional activator |
| 077933 | 4.7 | MLO-like protein |
| 078748 | 4.7 | ethylene-responsive element-binding protein |
| 018995 | 4.7 | monodehydroascorbate reductase |
| 063867 | 4.8 | ethylene-responsive transcription factor |
| 004586 | 4.8 | phenylalanine ammonia-lyase |
| 081281 | 4.8 | leucine-rich repeat receptor-like serine/threonine kinase |
| 013377 | 4.8 | 4-coumarate-CoA ligase |
| 080912 | 4.9 | ABC transporter B family member |
| 078836 | 4.9 | EIN3 domain-containing protein |
| 079312 | 4.9 | ABC transporter G family member |
| 078811 | 4.9 | multidrug resistance P-glycoprotein |
| 009128 | 5.0 | ethylene receptor-like protein |
| 058955 | 5.2 | monoterpene synthase |
| 079862 | 5.2 | shikimate O-hydroxycinnamoyltransferase |
| 076879 | 5.3 | linoleate 13S-lipoxygenase |
| 115915 | 5.5 | disease resistance protein |
| 017693 | 5.6 | terpene synthase |
| 080270 | 5.6 | glutathione S-transferase |
| 076381 | 5.6 | ABC transporter G family member |
| 147063 | 5.7 | ethylene-responsive transcription factor |
| 073453 | 5.9 | flavonoid C-glucosyltransferase |
| 141905 | 6.5 | chalcone-flavonone isomerase |
| 077293 | 6.7 | terpene synthase |
| 065968 | 7.1 | LRR receptor-like serine/threonine-protein kinase |
| 074648 | 7.1 | ethylene-responsive transcription factor |
| 058571 | 7.6 | flavonoid hydroxylase |
| 073617 | 7.6 | remorin |
| 134682 | 7.7 | leucoanthocyanidin reductase |
| 063666 | 8.0 | sieve element occlusion protein |
| 065392 | 8.2 | caffeoyl-CoA O-methyltransferase |
| 081612 | 8.3 | malonyl-CoA:anthocyanidin 5-O-glucoside |
| 006561 | 8.8 | naringenin-chalcone synthase |
| 148145 | 9.4 | glutathione S-transferase |
| 079417 | 9.7 | phenylalanine ammonia-lyase |
| 015737 | 12.0 | glutathione S-transferase |
| 051561 | 12.5 | naringenin,2-oxoglutarate 3-dioxygenase |
| 079297 | 12.8 | leucoanthocyanidin dioxygenase |
| Peak No. | Molecular Formula | m/z Measured and Ionization Mode | Mean Intensity Purple | Mean Intensity Yellow | t-Test | Identification |
|---|---|---|---|---|---|---|
| 1 | C5H10N2O3 | 147.0765-pos | 115,676 | 192,815 | 0.000 | glutamine |
| 2 | C6H12O6 | 219.02657-pos | 102,721 | 56,375 | 0.000 | glucose |
| 3 | C5H9N1O4 | 148.06045-pos | 30,462 | 33,943 | 0.002 | glutamic acid |
| 4 | C17H31N1O15 | 490.17676-pos | 26,190 | 32,155 | 0.001 | unknown |
| 5 | C12H22O11 | 341.1089-neg | 100,948 | 93,073 | 0.092 | sucrose |
| 6 | C6H8O6 | 175.02481-neg | 119,067 | 65,947 | 0.001 | ascorbic acid |
| 7 | C4H6O5 | 133.01433-neg | 164,764 | 153,107 | 0.028 | malic acid |
| 8 | C8H8O7 | 191.01975-neg | 154,828 | 18,986 | 0.000 | citric acid |
| 9 | C10H17N3O6S | 308.0911-pos | 59,473 | 48,847 | 0.035 | glutathione |
| 10 | C10H13N5O4 | 268.10409-pos | 220,255 | 301,724 | 0.002 | adenosine |
| 11 | C5H4O4 | 129.01819-pos | 21,815 | 7486 | 0.000 | unknown |
| 12 | C5H9N1O2 | 116.0705-pos | 3322 | 3144 | 0.445 | proline |
| 13 | C9H11N1O3 | 182.08121-pos | 10,101 | 8574 | 0.323 | tyrosine |
| 15 | C5H7N1O3 | 130.04991-pos | 219,080 | 63,362 | 0.000 | 5-oxoproline |
| 16 | C8H10N1 | 166.0862-pos | 16,439 | 69,894 | 0.000 | phenylalanine |
| 17 | C15H18N4O11 | 431.1048-pos | 56,678 | 6094 | 0.001 | unknown |
| 18 | C15H14O7 | 305.06663-neg | 39,016 | 0 | 0.000 | gallocatechin |
| 19 | C22H22O11 | 463.12378-pos | 296,537 | 578 | 0.000 | peonidin-3-glucoside |
| 20 | C15H14O6 | 291.08636-pos | 55,091 | 3370 | 0.000 | catechin |
| 21 | C21H20O11 | 449.10812-pos | 114,061 | 2179 | 0.000 | cyanidin-3-O-glucoside |
| 22 | C13H18O8 | 301.09273-neg | 54,632 | 2111 | 0.000 | unknown |
| 23 | C26H34O12 | 537.1979-neg | 9365 | 5135 | 0.001 | citrusin A/hyuganoside III |
| 24 | C21H22O11 | 449.10839-neg | 96,753 | 0 | 0.000 | unknown flavonoid |
| 25 | C21H22O11 | 451.12317-pos | 94,078 | 0 | 0.000 | unknown flavonoid |
| 26 | C26H34O12 | 537.19771-neg | 37,255 | 30,798 | 0.048 | citrusin A/hyuganoside III |
| 27 | C15H14O6 | 291.08639-pos | 105,416 | 0 | 0.000 | epicatechin |
| 28 | C20H27N1O10 | 440.1564-neg | 278,197 | 173,262 | 0.005 | prunasin-rhamnoside |
| 29 | C14H17N1O6 | 296.1128-pos | 45,051 | 45,072 | 0.993 | prunasin |
| 30 | C21H20O11 | 447.09301-neg | 7203 | 131,254 | 0.000 | C-glycosidic flavonoid |
| 31 | C19H28O10 | 415.1602-neg | 1437 | 0 | 0.000 | unknown |
| 32 | C27H30O14 | 579.17129-pos | 299,342 | 0 | 0.000 | unknown flavonoid |
| 33 | C27H30O16 | 609.14553-neg | 125,726 | 1676 | 0.000 | unknown flavonoid |
| 34 | C15H12O7 | 303.05087-neg | 32,535 | 0 | 0.000 | unknown |
| 35 | C26H32O11 | 521.2031-neg | 22,200 | 7319 | 0.000 | unknown |
| 36 | C21H20O12 | 463.08775-neg | 97,156 | 0 | 0.000 | unknown flavonoid |
| 37 | C16H20O9 | 395.07389-pos | 26,622 | 49,305 | 0.117 | unknown |
| 38 | C21H20O10 | 431.09818-neg | 622 | 30,081 | 0.000 | unknown |
| 39 | C17H19N1O9 | 382.11328-pos | 274,985 | 470,037 | 0.000 | prunasin malonate |
| 40 | C27H30O13 | 561.16079-neg | 5768 | 129,117 | 0.000 | unknown |
| 41 | C27H28O14 | 575.14075-neg | 82,622 | 1955 | 0.001 | unknown flavonoid |
| 42 | C21H20O10 | 433.11323-pos | 115,448 | 941 | 0.000 | unknown |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rizwan, H.M.; Zhimin, L.; Harsonowati, W.; Waheed, A.; Qiang, Y.; Yousef, A.F.; Munir, N.; Wei, X.; Scholz, S.S.; Reichelt, M.; et al. Identification of Fungal Pathogens to Control Postharvest Passion Fruit (Passiflora edulis) Decays and Multi-Omics Comparative Pathway Analysis Reveals Purple Is More Resistant to Pathogens than a Yellow Cultivar. J. Fungi 2021, 7, 879. https://doi.org/10.3390/jof7100879
Rizwan HM, Zhimin L, Harsonowati W, Waheed A, Qiang Y, Yousef AF, Munir N, Wei X, Scholz SS, Reichelt M, et al. Identification of Fungal Pathogens to Control Postharvest Passion Fruit (Passiflora edulis) Decays and Multi-Omics Comparative Pathway Analysis Reveals Purple Is More Resistant to Pathogens than a Yellow Cultivar. Journal of Fungi. 2021; 7(10):879. https://doi.org/10.3390/jof7100879
Chicago/Turabian StyleRizwan, Hafiz Muhammad, Lin Zhimin, Wiwiek Harsonowati, Abdul Waheed, Yang Qiang, Ahmed Fathy Yousef, Nigarish Munir, Xiaoxia Wei, Sandra S. Scholz, Michael Reichelt, and et al. 2021. "Identification of Fungal Pathogens to Control Postharvest Passion Fruit (Passiflora edulis) Decays and Multi-Omics Comparative Pathway Analysis Reveals Purple Is More Resistant to Pathogens than a Yellow Cultivar" Journal of Fungi 7, no. 10: 879. https://doi.org/10.3390/jof7100879
APA StyleRizwan, H. M., Zhimin, L., Harsonowati, W., Waheed, A., Qiang, Y., Yousef, A. F., Munir, N., Wei, X., Scholz, S. S., Reichelt, M., Oelmüller, R., & Chen, F. (2021). Identification of Fungal Pathogens to Control Postharvest Passion Fruit (Passiflora edulis) Decays and Multi-Omics Comparative Pathway Analysis Reveals Purple Is More Resistant to Pathogens than a Yellow Cultivar. Journal of Fungi, 7(10), 879. https://doi.org/10.3390/jof7100879