
How Dopamine Influences Survival and Cellular Immune Response of Rhipicephalus microplus Inoculated with Metarhizium anisopliae

 ,
,  , , , and
, , , and 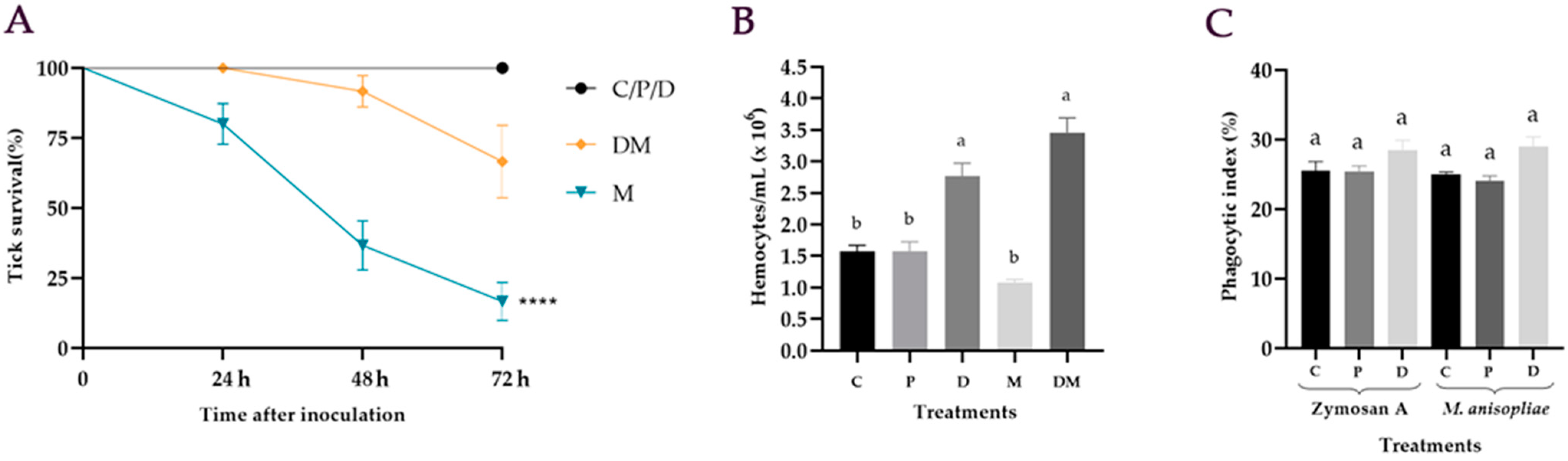{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ticks
2.2. Entomopathogenic Fungus
2.3. Effect of Dopamine on the Survival of Ticks Treated with M. anisopliae
2.4. Hemocytes Quantification after M. anisopliae Infection
2.5. In Vitro Phagocytic Assay
2.6. Phenoloxidase Activity in the Hemolymph of R. microplus Inoculated with M. anisopliae
2.7. Detection of Dopamine in Hemocytes of R. microplus Treated with M. anisopliae
2.8. Statistical Analysis
3. Results
3.1. Effect of Dopamine on the Survival of Ticks Treated with M. anisopliae
3.2. Hemocytes Quantification after M. anisopliae Infection
3.3. In Vitro Phagocytic Assay
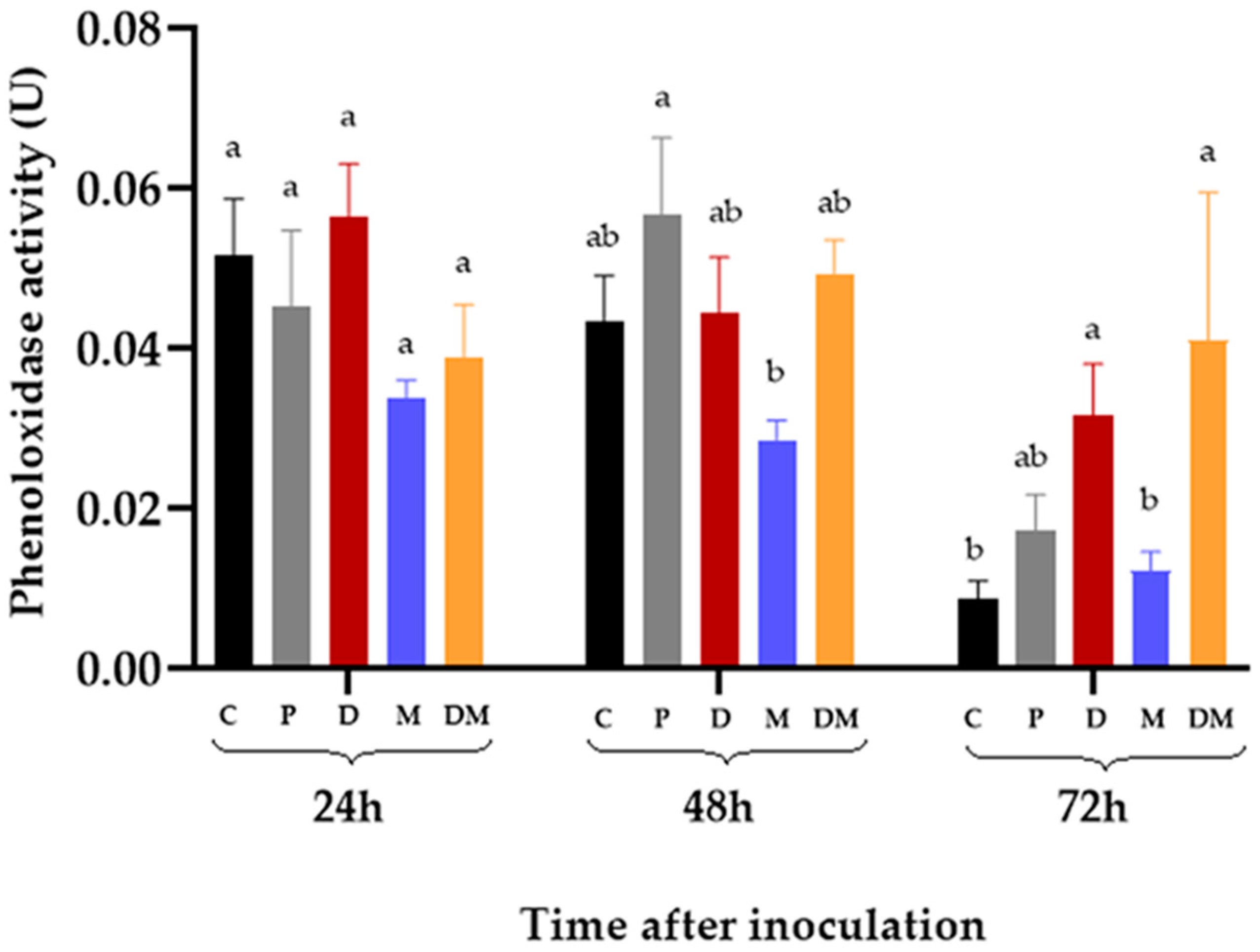
3.4. Phenoloxidase Activity in the Hemolymph of R. microplus Inoculated with M. anisopliae
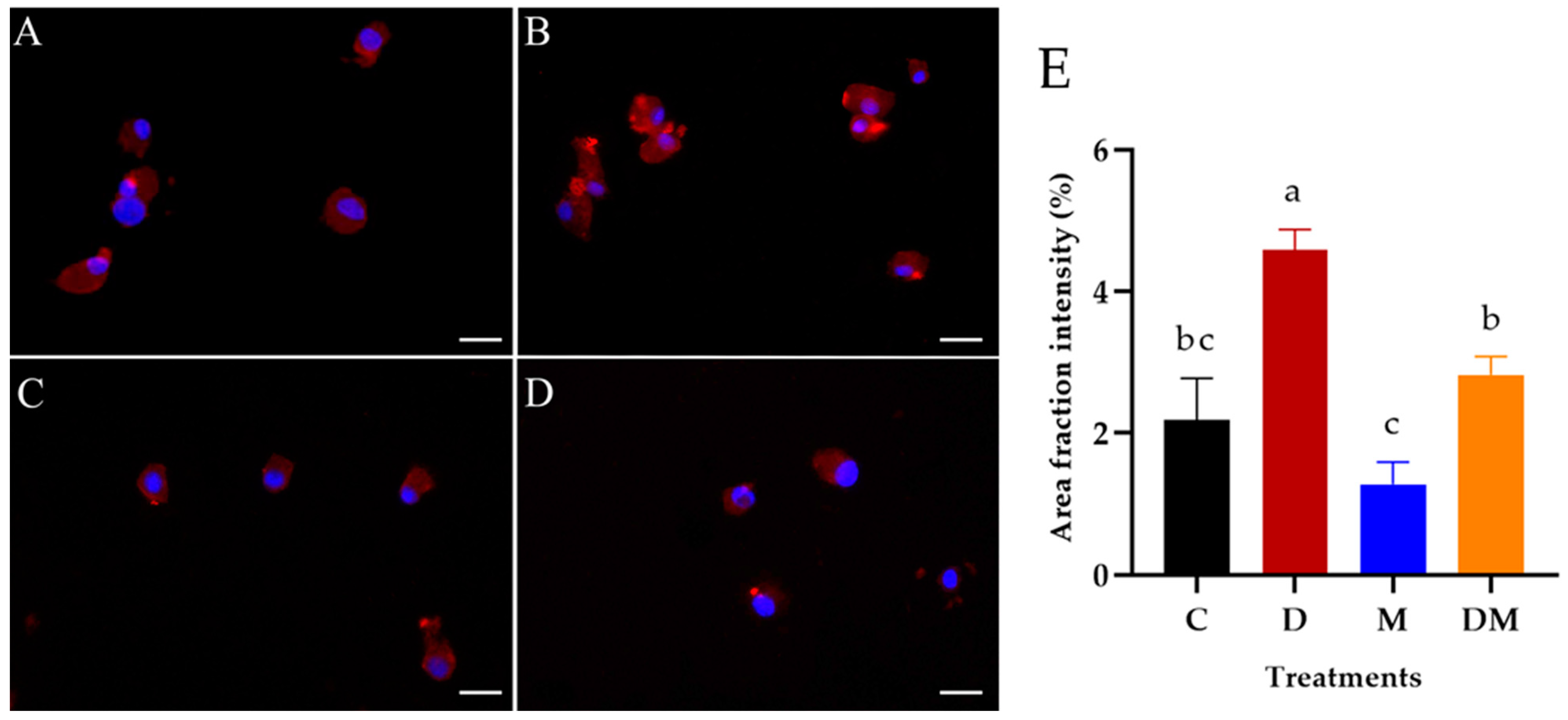
3.5. Detection of Dopamine in Hemocytes of R. microplus Treated with M. anisopliae
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grisi, L.; Leite, R.C.; Martins, J.R.S.; Barros, A.T.M.; Andreotti, R.; Cançado, P.H.D.; Léon, A.A.P.; Pereira, J.B.; Villela, H.S. Reassessment of potential economic impact of cattle parasites in Brazil. J. Vet. Parasitol. 2014, 23, 150–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klafke, G.M.; Castro-Janer, E.; Mendes, M.C.; Namindome, A.; Schumaker, T.T.S. Applicability of in vitro bioassays for the diagnosis of ivermectin resistance in Rhipicephalus microplus (Acari: Ixodidae). Vet. Parasitol. 2012, 184, 212–220. [Google Scholar] [CrossRef]
- Kernif, T.; Leulmi, H.; Raoult, D.; Parola, P. Emerging Tick-Borne Bacterial Pathogens. Microbiol. Spectr. 2016, 4, 10. [Google Scholar] [CrossRef] [PubMed]
- Reck, J.; Klafke, G.M.; Webster, A.; Dall’Agnol, B.; Scheffer, R.; Souza, U.A.; Corassini, V.B.; Vargas, R.; dos Santos, J.S.; Martins, J.R. First report of fluazuron resistance in Rhipicephalus microplus: A field tick population resistant to six classes of acaricides. Vet. Parasitol. 2014, 201, 128–136. [Google Scholar] [CrossRef]
- Klafke, G.; Webster, A.; Dall Agnol, B.; Pradel, E.; Silva, J.; de La Canal, L.H.; Becker, M.; Osório, M.F.; Mansson, M.; Barreto, R.; et al. Multiple resistance to acaricides in field populations of Rhipicephalus microplus from Rio Grande do Sul State; Southern Brazil. Ticks Tick Borne Dis. 2017, 8, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Camargo, M.G.; Nogueira, M.R.; Marciano, A.F.; Perinotto, W.M.; Coutinho-Rodrigues, C.J.; Scott, F.B.; Angelo, I.C.; Prata, M.C.; Bittencourt, V.R. Metarhizium anisopliae for controlling Rhipicephalus microplus ticks under field conditions. Vet. Parasitol. 2016, 223, 38–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camargo, M.G.; Marciano, A.F.; Sá, F.A.; Perinotto, W.M.; Quinelato, S.; Gôlo, P.S.; Angelo, I.C.; Prata, M.C.; Bittencourt, V.R. Commercial formulation of Metarhizium anisopliae for the control of Rhipicephalus microplus in a pen study. Vet. Parasitol. 2014, 205, 271–276. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Salas, A.; Alonso-Díaz, M.A.; Alonso-Morales, R.A.; Lezama-Gutiérrez, R.; Rodríguez-Rodríguez, J.C.; Cervantes-Chávez, J.A. Acaricidal activity of Metarhizium anisopliae isolated from paddocks in the Mexican tropics against two populations of the cattle tick Rhipicephalus microplus. Med. Vet. Entomol. 2017, 31, 36–43. [Google Scholar] [CrossRef]
- Bernardo, C.C.; Barreto, L.P.; e Silva, C.D.S.R.; Luz, C.; Arruda, W.; Fernandes, É.K.K. Conidia and blastospores of Metarhizium spp. and Beauveria bassiana s.l.: Their development during the infection process and virulence against the tick Rhipicephalus microplus. Ticks Tick Borne Dis. 2018, 9, 1334–1342. [Google Scholar] [CrossRef]
- Mascarin, G.M.; Lopes, R.B.; Delalibera, Í.; Fernandes, É.K.K.; Luz, C.; Faria, M. Current status and perspectives of fungal entomopathogens used for microbial control of arthropod pests in Brazil. J. Invertebr. Pathol. 2019, 165, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Marciano, A.F.; Mascarin, G.M.; Franco, R.F.F.; Golo, P.S.; Jaronski, S.T.; Fernandes, É.K.K.; Bittencourt, V.R. Innovative granular formulation of Metarhizium robertsii microsclerotia and blastospores for cattle tick control. Sci. Rep. 2021, 11, 4972. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, É.K.K.; Bittencourt, V.R.E.P.; Roberts, D.W. Perspectives on the potential of entomopathogenic fungi in biological control of ticks. Exp. Parasitol. 2012, 130, 300–305. [Google Scholar] [CrossRef]
- Angelo, I.C.; Gôlo, P.S.; Camargo, M.G.; Kluck, G.E.G.; Folly, E.; Bittencourt, V.R.E.P. Haemolymph protein and lipid profile of Rhipicephalus (Boophilus) microplus infected by fungi. Transbound Emerg. Dis. 2010, 57, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Fiorotti, J.; Gôlo, P.S.; Marciano, A.F.; Camargo, M.G.; Angelo, I.C.; Bittencourt, V.R.E.P. Disclosing hemolymph collection and inoculation of Metarhizium blastospores into Rhipicephalus microplus ticks towards invertebrate pathology studies. J. Vis. Exp. 2019, 148, e59899. [Google Scholar] [CrossRef]
- Fogaça, A.C.; Sousa, G.; Pavanelo, D.B.; Esteves, E.; Martins, L.A.; Urbanová, V.; Kopáček, P.; Daffre, S. Tick immune system: What is known; the interconnections; the gaps; and the challenges. Front. Immunol. 2021, 12, 628054. [Google Scholar] [CrossRef] [PubMed]
- Fiorotti, J.; Menna-Barreto, R.F.S.; Gôlo, P.S.; Coutinho-Rodrigues, C.J.B.; Bitencourt, R.O.B.; Spadacci-Morena, D.D.; Angelo, I.C.; Bittencourt, V.R.E.P. Ultrastructural and Cytotoxic Effects of Metarhizium robertsii Infection on Rhipicephalus microplus Hemocytes. Front. Physiol. 2019, 10, 654. [Google Scholar] [CrossRef] [PubMed]
- Adamo, S.A. Stress responses sculpt the insect immune system; optimizing defense in an ever-changing world. Dev. Comp. Immunol. 2016, 66, 24–32. [Google Scholar] [CrossRef]
- Kong, H.; Dong, C.; Tian, Z.; Mao, N.; Wang, C.; Cheng, Y.; Zhang, L.; Jiang, X.; Luo, L. Altered immunity in crowded Mythimna separata is mediated by octopamine and dopamine. Sci. Rep. 2018, 8, 3215. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.F.; Xu, G.; Stanley, D.; Huang, J.; Ye, G.Y. Dopamine modulates hemocyte phagocytosis via a D1-like receptor in the rice stem borer; Chilo suppressalis. Sci. Rep. 2015, 5, 12247. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Šimo, L.; Vancová, M.; Urban, J.; Park, Y. Neural and endocrine regulation of osmoregulatory organs in tick: Recent discoveries and implications. Gen. Comp. Endocrinol. 2019, 278, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Vancová, M.; Bílý, T.; Nebesářová, J.; Grubhoffer, L.; Bonnet, S.; Park, Y.; Šimo, L. Ultrastructural mapping of salivary gland innervation in the tick Ixodes ricinus. Sci. Rep. 2019, 9, 6860. [Google Scholar] [CrossRef]
- Cerenius, L.; Lee, B.L.; Söderhäll, K. The proPO-system: Pros and cons for its role in invertebrate immunity. Trends Immunol. 2008, 29, 263–271. [Google Scholar] [CrossRef]
- Bali, G.K.; Kaur, S.; Kour, B.G. Phenoloxidase activity in haemolymph of Spodoptera litura (Fabricius) mediating immune responses challenge with entomopathogenic fungus; Beauveria bassiana (Balsamo) Vuillemin. J. Entomol. Zool. Stud. 2013, 1, 118–123. [Google Scholar]
- Gillespie, J.P.; Bailey, A.M.; Cobb, B.; Vilcinskas, A. Fungi as elicitors of insect immune responses. Arch. Insect Biochem. Physiol. 2000, 44, 49–68. [Google Scholar] [CrossRef]
- Napolitano, A.; Di Donato, P.; Prota, G. New regulatory mechanisms in the biosynthesis of pheomelanins: Rearrangement vs. redox exchange reaction routes of a transient 2H-1,4-benzothia- zine-o-quinonimine intermediate. Acta Gen. 2000, 1475, 47–54. [Google Scholar] [CrossRef]
- Nappi, A.J.; Christensen, B.M. Melanogenesis and associated cytotoxic reactions: Applications to insect innate immunity. Insect Biochem. Mol. Biol. 2005, 35, 443–459. [Google Scholar] [CrossRef]
- Kadota, K.; Satoh, E.; Ochiai, M.; Inoue, N.; Tsuji, N.; Igarashi, I.; Nagasaka, H.; Mikami, T.; Claveria, F.G.; Fujisaki, K. Existence of phenol oxidase in the argasid tick Ornithodoros moubata. Parasitol. Res. 2002, 88, 781–784. [Google Scholar] [CrossRef]
- Feitosa, A.P.S.; Chaves, M.M.; Veras, D.L.; de Deus, D.M.V.; Portela, N.C.; Araújo, A.R.; Alves, L.C.; Brayner, F.A. Assessing the cellular and humoral immune response in Rhipicephalus sanguineus sensu lato (Acari: Ixodidae) infected with Leishmania infantum (Nicolle; 1908). Ticks Tick Borne Dis. 2018, 9, 1421–1430. [Google Scholar] [CrossRef] [PubMed]
- Mesquita, E.; Marciano, A.F.; Corval, A.R.C.; Fiorotti, J.; Corrêa, T.A.; Quinelato, S.; Bittencourt, V.R.E.P.; Gôlo, P.S. Efficacy of a native isolate of the entomopathogenic fungus Metarhizium anisopliae against larval tick outbreaks under semifield conditions. BioControl 2020, 65, 353–362. [Google Scholar] [CrossRef]
- Bitencourt, R.O.B.; Mallet, J.R.S.; Mesquita, E.; Gôlo, P.S.; Fiorotti, J.; Bittencourt, V.R.E.P.; Pontes, E.G.; Angelo, I.C. Larvicidal activity; route of interaction and ultrastructural changes in Aedes aegypti exposed to entomopathogenic fungi. Acta Trop. 2021, 213, 105732. [Google Scholar] [CrossRef]
- De Paulo, J.F.; Camargo, M.G.; Coutinho-Rodrigues, C.J.B.; Marciano, A.F.; de Freita, M.C.; da Silva, E.M.; Gôlo, P.S.; Spadacci-Morena, D.D.; Angelo, I.C.; Bittencourt, V.R.E.P. Rhipicephalus microplus infected by Metarhizium: Unveiling hemocyte quantification; GFP-fungi virulence; and ovary infection. Parasitol. Res. 2018, 117, 1847–1856. [Google Scholar] [CrossRef] [PubMed]
- Kuklinski, N.J.; Berglund, E.C.; Engelbreksson, J.; Ewing, A.G. Determination of salsolinol; norsalsolinol; and twenty-one biogenic amines using micellar electrokinetic capillary chromatography-electrochemical detection. Electrophoresis 2010, 31, 1886–1893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brayner, F.A.; Araújo, H.R.C.; Cavalcanti, M.G.S.; Alves, L.C.; Peixoto, C.A. Ultrastructural characterization of the hemocytes of Culex quinquefasciatus (Diptera: Culicidae). Micron 2005, 36, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Maina, U.M.; Galadima, I.B.; Gambo, F.M.; Zakaria, D. A review on the use of entomopathogenic fungi in the management of insect pests of field crops. J. Entomol. Zool. Stud. 2018, 6, 27–32. [Google Scholar]
- Bernardo, C.C.; Pereira-Junior, R.A.; Luz, C.; Mascarin, G.M.; Fernandes, É.K.K. Differential susceptibility of blastospores and aerial conidia of entomopathogenic fungi to heat and UV-B stresses. Fungal Biol. 2020, 124, 714–722. [Google Scholar] [CrossRef] [PubMed]
- Nattion, J.L. Insect Physiology and Biochemistry; University of Florida: Gainesville, FL, USA, 2016; 672p. [Google Scholar]
- Chang, C.C.; Wu, Z.R.; Kuo, C.M.; Cheng, W. Dopamine depresses immunity in the tiger shrimp Penaeus monodon. Fish Shellfish Immunol. 2007, 23, 24–33. [Google Scholar] [CrossRef]
- Loosová, G.; Jindrák, L.; Kopáček, P. Mortality caused by experimental infection with the yeast Candida haemulonii in the adults of Ornithodoros moubata (Acarina: Argasidae). Folia Parasitol. 2001, 48, 149–153. [Google Scholar] [CrossRef] [Green Version]
- Pereira, L.S.; Oliveira, P.L.; Barja-Fidalgo, C.; Daffre, S. Production of reactive oxygen species by hemocytes from the cattle tick Boophilus microplus. Exp. Parasitol. 2001, 99, 66–72. [Google Scholar] [CrossRef]
- Buresova, V.; Hajdusek, O.; Franta, Z.; Sojka, D.; Kopacek, P. IrAM-An alpha2-macroglobulin from the hard tick Ixodes ricinus: Characterization and function in phagocytosis of a potential pathogen Chryseobacterium indologenes. Dev. Comp. Immunol. 2009, 33, 489–498. [Google Scholar] [CrossRef] [PubMed]
- Urbanová, V.; Hartmann, D.; Grunclová, L.; Šíma, R.; Flemming, T.; Hajdušek, O. IrFC—An Ixodes ricinus injury-responsive molecule related to Limulus Factor C. Dev. Comp. Immunol. 2014, 46, 439–447. [Google Scholar] [CrossRef]
- Urbanova, V.; Hajdušek, O.; Mondekova, H.H.; Šima, R.; Kopaček, P. Tick thioester-containing proteins and phagocytosis do not affect transmission of Borrelia afzelii from the competent Vector Ixodes ricinus. Front. Cell. Infect. Microbiol. 2017, 7, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiorotti, J.; Urbanová, V.; Gôlo, P.S.; Bittencourt, V.R.E.P.; Kopáček, P. The role of complement in the tick cellular immune defense against the entomopathogenic fungus Metarhizium robertsii. Dev. Comp. Immunol. 2022, 126, 104234. [Google Scholar] [CrossRef]
- Cerenius, L.; Söderhäll, K. The prophenoloxidase-activating system in invertebrates. Immunol. Rev. 2004, 198, 116–126. [Google Scholar] [CrossRef] [PubMed]
- Mello, C.B.; Garcia, E.S.; Ratcliffe, N.A.; Azambuja, P. Trypanosoma cruzi and Trypanosoma rangeli: Interplay with hemolymph components of Rhodnius prolixus. J. Invertebr. Pathol. 1995, 65, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Castro, D.P.; Moraes, C.S.; Gonzalez, M.S.; Ratcliffe, N.A.; Azambuja, P.; Garcia, E.S. Trypanosoma cruzi immune response modulation decreases microbiota in Rhodnius prolixus gut and is crucial for parasite survival and development. PLoS ONE 2012, 7, e36591. [Google Scholar] [CrossRef] [Green Version]
- Laino, A.; Lavarías, S.; Suárez, G.; Lino, A.; Cunningham, M. Characterization of phenoloxidase activity from spider Polybetes pythagoriucus hemocyanin. J. Exp. Zool. 2015, 323, 547–555. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Corrêa, T.A.; Fiorotti, J.; Mesquita, E.; Meirelles, L.N.; Camargo, M.G.; Coutinho-Rodrigues, C.J.B.; Marciano, A.F.; Bittencourt, V.R.E.P.; Golo, P.S. How Dopamine Influences Survival and Cellular Immune Response of Rhipicephalus microplus Inoculated with Metarhizium anisopliae. J. Fungi 2021, 7, 950. https://doi.org/10.3390/jof7110950
Corrêa TA, Fiorotti J, Mesquita E, Meirelles LN, Camargo MG, Coutinho-Rodrigues CJB, Marciano AF, Bittencourt VREP, Golo PS. How Dopamine Influences Survival and Cellular Immune Response of Rhipicephalus microplus Inoculated with Metarhizium anisopliae. Journal of Fungi. 2021; 7(11):950. https://doi.org/10.3390/jof7110950
Chicago/Turabian StyleCorrêa, Thaís Almeida, Jéssica Fiorotti, Emily Mesquita, Laura Nóbrega Meirelles, Mariana Guedes Camargo, Caio Junior Balduino Coutinho-Rodrigues, Allan Felipe Marciano, Vânia Rita Elias Pinheiro Bittencourt, and Patrícia Silva Golo. 2021. "How Dopamine Influences Survival and Cellular Immune Response of Rhipicephalus microplus Inoculated with Metarhizium anisopliae" Journal of Fungi 7, no. 11: 950. https://doi.org/10.3390/jof7110950
APA StyleCorrêa, T. A., Fiorotti, J., Mesquita, E., Meirelles, L. N., Camargo, M. G., Coutinho-Rodrigues, C. J. B., Marciano, A. F., Bittencourt, V. R. E. P., & Golo, P. S. (2021). How Dopamine Influences Survival and Cellular Immune Response of Rhipicephalus microplus Inoculated with Metarhizium anisopliae. Journal of Fungi, 7(11), 950. https://doi.org/10.3390/jof7110950