
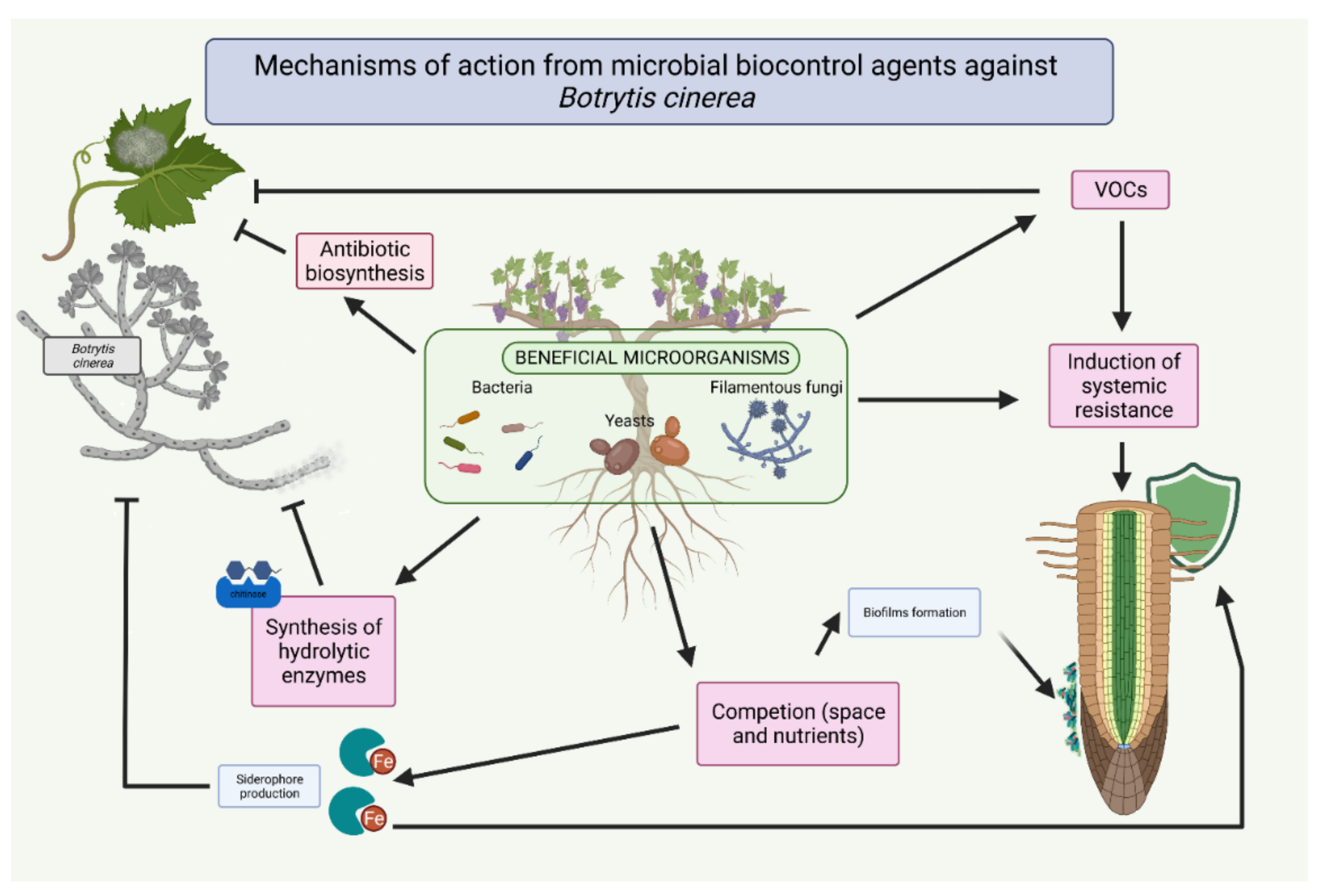
Mechanisms of Action of Microbial Biocontrol Agents against Botrytis cinerea




Abstract
:1. Introduction
2. Production of Diffusible Molecules
2.1. Antimicrobial Molecules
2.2. Siderophores
3. Synthesis of Volatile Organic Compounds
4. Hydrolytic Enzymes
5. Other Mechanisms
5.1. Competition
5.2. Induction of Systemic Resistance
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Lazarovits, G.; Turnbull, A.; Johnston-Monje, D. Plant Health Management: Biological Control of Plant Pathogens. In Encyclopedia of Agriculture and Food Systems; Elsevier Ltd.: London, UK, 2014; pp. 388–399. [Google Scholar]
- Timothy, O.; Adejumo, R.T. Biopesticides: Botanicals and Microorganisms for Improving Agriculture and Human Health; Ed. Logos-Verlag Berlin: Berlin, Germany, 2021; pp. 106–111. [Google Scholar]
- Fisher, M.C.; Henk, D.A.; Briggs, C.J.; Brownstein, J.S.; Madoff, L.C.; McCraw, S.L.; Gurr, S.J. Emerging fungal threats to animal, plant and ecosystem health. Nature 2012, 484, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Bolívar-Anillo, H.J.; Garrido, C.; Collado, I.G. Endophytic microorganisms for biocontrol of the phytopathogenic fungus Botrytis cinerea. Phytochem. Rev. 2020, 19, 721–740. [Google Scholar] [CrossRef]
- Doehlemann, G.; Ökmen, B.; Zhu, W.; Sharon, A. Plant Pathogenic Fungi. In The Fungal Kingdom; American Society of Microbiology, Ed.; Wiley: Hoboken, NJ, USA, 2017; pp. 703–726. [Google Scholar]
- Rodriguez-Moreno, L.; Ebert, M.K.; Bolton, M.D.; Thomma, B.H. Tools of the crook-infection strategies of fungal plant pathogens. Plant J. 2018, 93, 664–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cordero-Bueso, G.; Mangieri, N.; Maghradze, D.; Foschino, R.; Valdetara, F.; Cantoral, J.M.; Vigentini, I. Wild Grape-Associated Yeasts as Promising Biocontrol Agents against Vitis vinifera Fungal Pathogens. Front. Microbiol. 2017, 8, 2025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casadevall, A. Determinants of virulence in the pathogenic fungi. Fungal Biol. Rev. 2007, 21, 130–132. [Google Scholar] [CrossRef] [Green Version]
- Taylor, T.N.; Krings, M.; Taylor, E.L. Fungal Spores. In Fossil Fungi; Elsevier: London, UK, 2015; pp. 221–238. [Google Scholar]
- Hua, L.; Yong, C.; Zhanquan, Z.; Boqiang, L.; Guozheng, Q.; Shiping, T. Pathogenic mechanisms and control strategies of Botrytis cinerea causing post-harvest decay in fruits and vegetables. Food Qual. Saf. 2018, 2, 111–119. [Google Scholar] [CrossRef] [Green Version]
- Dean, R.; Van Kan, J.A.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [Green Version]
- Shaw, M.W.; Emmanuel, C.J.; Emilda, D.; Terhem, R.B.; Shafia, A.; Tsamaidi, D.; Emblow, M.; van Kan, J.A. Analysis of Cryptic, Systemic Botrytis Infections in Symptomless Hosts. Front. Plant Sci. 2016, 7, 625. [Google Scholar] [CrossRef] [Green Version]
- Pinedo, C.; Moraga, J.; Collado, I.G.; Aleu, J. Botrytis species as biocatalysts. Phytochem. Rev. 2020, 19, 529–558. [Google Scholar] [CrossRef]
- Elad, Y. Mechanisms involved in the biological control of Botrytis cinerea incited diseases. Eur. J. Plant Pathol. 1996, 102, 719–732. [Google Scholar] [CrossRef]
- Williamson, B.; Tudzynski, B.; Tudzynski, P.; Van Kan, J.A. Botrytis cinerea: The cause of grey mould disease. Mol. Plant Pathol. 2007, 8, 561–580. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Glawe, D.A.; Kramer, E.; Weller, D.; Okubara, A. Biological Control of Botrytis cinerea: Interactions with Native Vineyard Yeasts from Washington State. Phytopathology 2018, 108, 691–701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, J.F.; Sun, C.Q. Isolation, identification, and biocontrol of antagonistic bacterium against Botrytis cinerea after tomato harvest. Braz. J. Microbiol. 2017, 48, 706–714. [Google Scholar] [CrossRef] [PubMed]
- Plesken, C.; Pattar, P.; Reiss, B.; Noor, Z.N.; Zhang, L.; Klug, K.; Huettel, B.; Hahn, M. Genetic Diversity of Botrytis cinerea Revealed by Multilocus Sequencing, and Identification of B. cinerea Populations Showing Genetic Isolation and Distinct Host Adaptation. Front. Plant Sci. 2021, 12, 663027. [Google Scholar] [CrossRef]
- Chen, P.H.; Chen, R.Y.; Chou, J.Y. Screening and evaluation of yeast antagonists for biological control of Botrytis cinerea on strawberry fruits. Mycobiology 2018, 46, 33–46. [Google Scholar] [CrossRef] [Green Version]
- González-Domínguez, E.; Caffi, T.; Ciliberti, N.; Rossi, V. A mechanistic model of botrytis cinerea on grapevines that includes weather, vine growth stage, and the main infection pathways. PLoS ONE 2015, 10, e0140444. [Google Scholar] [CrossRef] [Green Version]
- Elmer, A.G.; Michailides, T.J. Epidemiology of Botrytis cinerea in Orchard and Vine Crops. In Botrytis: Biology, Pathology and Control; Springer: Dordrecht, The Netherlands, 2007; pp. 243–272. [Google Scholar]
- Van Kan, J.A. Infection Strategies of Botrytis cinerea. Acta Hortic. 2005, 669, 77–90. [Google Scholar] [CrossRef]
- Doss, R.; Potter, S.W.; Chastagner, G.A.; Christian, J.K. Adhesion of Nongerminated Botrytis cinerea Conidia to Several Substrata. Appl. Environ. Microbiol. 1993, 59, 1786–1791. [Google Scholar] [CrossRef] [Green Version]
- Cole, G.T. Basic Biology of Fungi. In Medical Microbiology, 4th ed.; University of Texas Medical Branch at Galveston: Galveston, TX, USA, 1996. [Google Scholar]
- Williamson, B.; Duncan, G.H.; Harrison, J.G.; Harding, L.A.; Elad, Y.; Zimand, G. Effect of humidity on infection of rose petals by dry-inoculated conidia of Botrytis cinerea. Mycol. Res. 1995, 99, 1303–1310. [Google Scholar] [CrossRef]
- Harper, A.M.; Strange, R.N.; Langcake, P. Characterization of the nutrients required by Botrytis cinerea to infect broad bean leaves. Physiol. Plant Pathol. 1981, 19, 153–167. [Google Scholar] [CrossRef]
- Elad, Y.; Volpin, H. The involvement of ethylene and calcium in gray mold of pelargonium, ruscus, and rose plants. Phytoparasitiea 1988, 16, 119–131. [Google Scholar] [CrossRef]
- McNicol, R.J.; Williamson, B.; Young, K. Ethylene production by black currant flowers infected by Botrytis cinerea. Acta Hortic. 1989, 262, 209–216. [Google Scholar] [CrossRef]
- Wojtaszek, P. Oxidative burst: An early plant response to pathogen infection. Biochem. J. 1997, 322, 681–692. [Google Scholar] [CrossRef] [Green Version]
- Rebordinos, L.; Cantoral, J.M.; Prieto, M.V.; Hanson, J.R.; Collado, I.G. The phytotoxic activity of some metabolites of Botrytis cinerea. Phytochemistry 1996, 42, 383–387. [Google Scholar] [CrossRef]
- Tiedemann, A.V. Evidence for a primary role of active oxygen species in induction of host cell death during infection of bean leaves with Botrytis cinerea. Physiol. Mol. Plant Pathol. 1997, 50, 151–166. [Google Scholar] [CrossRef]
- Ten Have, A.; Tenberge, K.B.; Benen, J.A.; Tudzynski, P.; Visser, J.; van Kan, J.A. The Contribution of Cell Wall Degrading Enzymes to Pathogenesis of Fungal Plant Pathogens. In Agricultural Applications; Springer: Berlin/Heidelberg, Germany, 2002; pp. 341–358. [Google Scholar]
- Lian, Q.; Zhang, J.; Gan, L.; Ma, Q.; Zong, Z.; Wang, Y. The biocontrol efficacy of Streptomyces pratensis LMM15 on Botrytis cinerea in Tomato. Biomed. Res. Int. 2017, 2017, 9486794. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.C.; Hur, J.Y.; Park, S.K. Biocontrol of Botrytis cinerea by chitin-based cultures of Paenibacillus elgii HOA73. Eur. J. Plant Pathol. 2019, 155, 253–263. [Google Scholar] [CrossRef]
- Jiao, X.; Takishita, Y.; Zhou, G.; Smith, D.L. Plant Associated Rhizobacteria for Biocontrol and Plant Growth Enhancement. Front. Plant Sci. 2021, 12, 17. [Google Scholar] [CrossRef]
- Leroch, M.; Plesken, C.; Weber, R.W.; Kauff, F.; Scalliet, G.; Hahn, M. Gray mold populations in German strawberry fields are resistant to multiple fungicides and dominated by a novel clade closely related to Botrytis cinerea. Appl. Environ. Microbiol. 2013, 79, 159–167. [Google Scholar] [CrossRef] [Green Version]
- Pertot, I.; Giovannini, O.; Benanchi, M.; Caffi, T.; Rossi, V.; Mugnai, L. Combining biocontrol agents with different mechanisms of action in a strategy to control Botrytis cinerea on grapevine. Crop Prot. 2017, 97, 85–93. [Google Scholar] [CrossRef]
- Haidar, R.; Fermaud, M.; Calvo-Garrido, C.; Roudet, J.; Deschamps, A. Modes of action for biological control of Botrytis cinerea by antagonistic bacteria. Phytopathol. Mediterr. 2017, 55, 301–322. [Google Scholar]
- Damalas, C.A.; Koutroubas, S.D. Current Status and Recent Developments in Biopesticide Use. Agriculture 2018, 8, 13. [Google Scholar] [CrossRef] [Green Version]
- Chandler, D.; Bailey, A.S.; Tatchell, G.M.; Davidson, G.; Greaves, J.; Grant, W.P. The development, regulation and use of biopesticides for integrated pest management. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 1987–1998. [Google Scholar] [CrossRef] [PubMed]
- Schalchli, H.; Tortella, G.R.; Rubilar, O.; Parra, L.; Hormazabal, E.; Quiroz, A. Fungal volatiles: An environmentally friendly tool to control pathogenic microorganisms in plants. Crit. Rev. Biotechnol. 2016, 36, 144–152. [Google Scholar] [CrossRef]
- Ons, L.; Bylemans, D.; Thevissen, K.; Cammue, B.P. Combining Biocontrol Agents with Chemical Fungicides for Integrated Plant Fungal Disease Control. Microorganisms 2020, 8, 1930. [Google Scholar] [CrossRef]
- Hahn, M. The rising threat of fungicide resistance in plant pathogenic fungi: Botrytis as a case study. J. Chem. Biol. 2014, 7, 133–141. [Google Scholar] [CrossRef] [Green Version]
- Fan, F.; Hamada, M.S.; Li, N.; Li, G.Q.; Luo, C.X. Multiple fungicide resistance in Botrytis cinerea from greenhouse strawberries in Hubei Province, China. Plant Dis. 2017, 101, 601–606. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, J.E.; Gaudin, A.C. What is the agronomic potential of biofertilizers for maize? A meta-analysis. FEMS Microbiol. Ecol. 2018, 94, fiy094. [Google Scholar] [CrossRef]
- Umesha, S.; Singh, K.; Singh, R. Microbial Biotechnology and Sustainable Agriculture. In Biotechnology for Sustainable Agriculture: Emerging Approaches and Strategies; Elsevier: London, UK, 2017; pp. 185–205. [Google Scholar]
- Compant, S.; Duffy, B.; Nowak, J.; Clément, C.; Barka, E.A. Use of plant growth-promoting bacteria for biocontrol of plant diseases: Principles, mechanisms of action, and future prospects. Appl. Environ. Microbiol. Am. Soc. Microbiol. ASM 2005, 71, 4951–4959. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.L.; Waqas, M.; Kang, S.M.; Al-Harrasi, A.; Hussain, J.; Al-Rawahi, A.; Al-Khiziri, S.; Ullah, I.; Ali, L.; Jung, H.Y.; et al. Bacterial endophyte Sphingomonas sp. LK11 produces gibberellins and IAA and promotes tomato plant growth. J. Microbiol. 2014, 52, 689–695. [Google Scholar] [CrossRef]
- Nair, D.N.; Padmavathy, S. Impact of endophytic microorganisms on plants, environment and humans. Sci. World J. 2014, 2014, 250693. [Google Scholar] [CrossRef] [Green Version]
- Beneduzi, A.; Ambrosini, A.; Passaglia, L.M. Plant growth-promoting rhizobacteria [PGPR]: Their potential as antagonists and biocontrol agents. Genet. Mol. Biol. Soc. Bras. Genética 2012, 35, 1044–1051. [Google Scholar] [CrossRef] [Green Version]
- De Silva, N.I.; Brooks, S.; Lumyong, S.; Hyde, K.D. Use of endophytes as biocontrol agents. Fungal Biol. Rev. 2019, 33, 133–148. [Google Scholar] [CrossRef]
- Vellasamy, S.; Hariharan, H. Controlling Strategies (Diagnosis/Quarantine/Eradication) of Plant Pathogenic Bacteria. Sustain. Approaches Control Plant Pathog. Bact. 2015, 98–127. [Google Scholar]
- Jiang, C.H.; Liao, M.J.; Wang, H.K.; Zheng, M.Z.; Xu, J.J.; Guo, J.H. Bacillus velezensis, a potential and efficient biocontrol agent in control of pepper gray mold caused by Botrytis cinerea. Biol. Control. 2018, 126, 147–157. [Google Scholar] [CrossRef]
- Kasfi, K.; Taheri, P.; Jafarpour, B.; Tarighi, S. Identification of epiphytic yeasts and bacteria with potential for biocontrol of grey mold disease on table grapes caused by Botrytis cinerea. Span. J. Agric. Res. 2018, 16, 1002. [Google Scholar] [CrossRef]
- Sansinenea, E.; Ortiz, A. Secondary metabolites of soil Bacillus spp. Biotechnol. Lett. 2011, 33, 1523–1538. [Google Scholar] [CrossRef]
- Toral, L.; Rodríguez, M.; Béjar, V.; Sampedro, I. Antifungal Activity of Lipopeptides from Bacillus XT1 CECT 8661 against Botrytis cinerea. Front. Microbiol. 2018, 9, 1315. [Google Scholar] [CrossRef]
- Raaijmakers, J.M.; De Bruijn, I.; Nybroe, O.; Ongena, M. Natural functions of lipopeptides from Bacillus and Pseudomonas: More than surfactants and antibiotics. FEMS Microbiol. Rev. 2010, 34, 1037–1062. [Google Scholar] [CrossRef] [Green Version]
- Elad, Y.; Williamson, B.; Tudzynski, P.; Delen, N. Botrytis: Biology, Pathology and Control; Springer Science & Business Media: Germany, Berlin/Heidelberg, 2004; pp. 1–403. [Google Scholar]
- Wang, X.; Zhou, X.; Cai, Z.; Guo, L.; Chen, X.; Chen, X.; Liu, J.; Feng, M.; Qiu, Y.; Zhang, Y.; et al. A biocontrol strain of Pseudomonas aeruginosa CQ-40 promote growth and control Botrytis cinerea in tomato. Pathogens 2021, 10, 22. [Google Scholar] [CrossRef]
- Ash, C.; Priest, F.G.; Collins, M.D. Molecular identification of rRNA group 3 bacilli [Ash, Farrow, Wallbanks and Collins] using a PCR probe test. Antonie Leeuwenhoek 1993, 64, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Thambugala, K.M.; Daranagama, D.A.; Phillips, A.J.; Kannangara, S.D.; Promputtha, I. Fungi vs. Fungi in Biocontrol: An Overview of Fungal Antagonists Applied against Fungal Plant Pathogens. Front. Cell. Infect. Microbiol. 2020, 10, 718. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Sui, Y.; Wisniewski, M.; Droby, S.; Liu, Y. Review: Utilization of antagonistic yeasts to manage postharvest fungal diseases of fruit. Int. J. Food Microbiol. 2013, 167, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Parafati, L.; Vitale, A.; Polizzi, G.; Restuccia, C.; Cirvilleri, G. Understanding the mechanism of biological control of postharvest phytopathogenic moulds promoted by food isolated yeasts. Acta Hortic. 2016, 1144, 93–100. [Google Scholar] [CrossRef]
- Saravanakumar, D.; Ciavorella, A.; Spadaro, D.; Garibaldi, A.; Gullino, M.L. Metschnikowia pulcherrima strain MACH1 outcompetes Botrytis cinerea, Alternaria alternata and Penicillium expansum in apples through iron depletion. Postharvest Biol. Technol. 2008, 49, 121–128. [Google Scholar] [CrossRef]
- Freimoser, F.M.; Rueda-Mejia, M.; Tilocca, B.; Migheli, Q. Biocontrol yeasts: Mechanisms and applications. World J. Microbiol. Biotechnol. 2019, 35, 154. [Google Scholar] [CrossRef] [Green Version]
- Jijakli, M.H.; Lepoivre, P. Characterization of an exo-β-1,3-glucanase produced by Pichia anomala strain K, antagonist of Botrytis cinerea on apples. Phytopathology 1998, 88, 335–343. [Google Scholar] [CrossRef] [Green Version]
- Masih, E.I.; Alie, I.; Paul, B. Can the grey mould disease of the grape-vine be controlled by yeast? FEMS Microbiol. Lett. 2000, 189, 233–237. [Google Scholar] [CrossRef] [Green Version]
- Leroux, P. Chemical control of Botrytis and Its Resistance To Chemical Fungicides. In Botrytis: Biology, Pathology and Control; Springer: Dordrecht, The Netherlands, 2007; pp. 195–222. [Google Scholar]
- Lim, S.M.; Yoon, M.Y.; Choi, G.J.; Choi, Y.H.; Jang, K.S.; Shin, T.S.; Park, H.W.; Yu, N.H.; Kim, Y.H.; Kim, J.C. Diffusible and volatile antifungal compounds produced by an antagonistic Bacillus velezensis G341 against various phytopathogenic fungi. Plant Pathol. J. 2017, 33, 488–498. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Song, C.; Bai, Y.; Hu, J.; Pan, H. Cytotoxic and antimicrobial activities of secondary metabolites isolated from the deep-sea-derived Actinoalloteichus cyanogriseus 12A22. 3 Biotech 2021, 11, 283. [Google Scholar] [CrossRef]
- Barka, E.A.; Belarbi, A.; Hachet, C.; Nowak, J.; Audran, J.C. Enhancement of in vitro growth and resistance to gray mould of Vitis vinifera co-cultured with plant growth-promoting rhizobacteria. FEMS Microbiol. Lett. 2000, 186, 91–95. [Google Scholar] [CrossRef] [Green Version]
- Su, Z.; Chen, X.; Liu, X.; Guo, Q.; Li, S.; Lu, X.; Zhang, X.; Wang, P.; Dong, L.; Zhao, W.; et al. Genome mining and UHPLC–QTOF–MS/MS to identify the potential antimicrobial compounds and determine the specificity of biosynthetic gene clusters in Bacillus subtilis NCD-2. BMC Genom. 2020, 21, 767. [Google Scholar]
- Nifakos, K.; Tsalgatidou, C.; Thomloudi, E.E.; Skagia, A.; Kotopoulis, D.; Baira, E.; Delis, C.; Papadimitriou, K.; Markellou, E.; Venieraki, A.; et al. Genomic Analysis and Secondary Metabolites Production of the Endophytic Bacillus velezensis Bvel1: A Biocontrol Agent against Botrytis cinerea Causing Bunch Rot in Post-Harvest Table Grapes. Plants 2021, 10, 1716. [Google Scholar] [CrossRef]
- Lambrese, Y.; Guiñez, M.; Calvente, V.; Sansone, G.; Cerutti, S.; Raba, J.; Sanz Ferramola, M.I. Production of siderophores by the bacterium Kosakonia radicincitans and its application to control of phytopathogenic fungi. Bioresour. Technol. Rep. 2018, 3, 82–87. [Google Scholar] [CrossRef] [Green Version]
- Priyanka, R.; Nakkeeran, S. Ochrobactrum ciceri mediated induction of defence genes and antifungal metabolites enhance the biocontrol efficacy for the management of Botrytis leaf blight of Lilium under protected conditions. J. Plant Pathol. 2019, 101, 323–337. [Google Scholar] [CrossRef]
- De Moura, G.G.; de Barros, A.V.; Machado, F.; Martins, A.D.; da Silva, C.M.; Durango, L.G.; Forim, M.; Alves, E.; Pasqual, M.; Doriad, J. Endophytic bacteria from strawberry plants control gray mold in fruits via production of antifungal compounds against Botrytis cinerea L. Microbiol. Res. 2021, 251, 126793. [Google Scholar] [CrossRef]
- Aznar, A.; Dellagi, A. New insights into the role of siderophores as triggers of plant immunity: What can we learn from animals? J. Exp. Bot. 2015, 66, 3001–3010. [Google Scholar] [CrossRef] [Green Version]
- Haggag, W.M.; Abo, M.; Soud, E. Production and Optimization of Pseudomonas fluorescens Biomass and Metabolites for Biocontrol of Strawberry Grey Mould. Am. J. Plant Sci. 2012, 3, 836–845. [Google Scholar] [CrossRef] [Green Version]
- Sansone, G.; Rezza, I.; Fernández, G.; Calvente, V.; Benuzzi, D.; Sanz, M.I. Inhibitors of polygalacturonase and laccase of Botrytis cinerea and their application to the control of this fungus. Int. Biodeterior. Biodegradation. 2011, 65, 243–247. [Google Scholar] [CrossRef]
- De Cesare, G.B.; Cristy, S.A.; Garsin, D.A.; Lorenz, M.C. Antimicrobial peptides: A new frontier in antifungal therapy. MBio. 2020, 11, e02123-20. [Google Scholar]
- Coleman, J.J.; Ghosh, S.; Okoli, I.; Mylonakis, E. Antifungal activity of microbial secondary metabolites. PLoS ONE 2011, 6, 25321. [Google Scholar] [CrossRef] [Green Version]
- Di Pietro, A. Endochitinase from Gliocladium virens: Isolation, Characterization, and Synergistic Antifungal Activity in Combination with Gliotoxin. Phytopathology 1993, 83, 308. [Google Scholar] [CrossRef]
- Brunner, K.; Zeilinger, S.; Ciliento, R.; Woo, S.L.; Lorito, M.; Kubicek, C.; Mach, R.L. Improvement of the Fungal Biocontrol Agent Trichoderma atroviride To Enhance both Antagonism and Induction of Plant Systemic Disease Resistance. Appl. Environ. Microbiol. 2005, 71, 3959. [Google Scholar] [CrossRef] [Green Version]
- Marín-Chacón, M.A.; Rivera-Coto, G.; Villalobos-Moya, K.; Orozco-Rodríguez, R.; Orozco-Cayasso, S. Evaluation of Botrytis cinerea Pers., antagonist fungi in blackberry plantations of Costa Rica. Agron. Costarric. 2017, 41, 7–18. [Google Scholar]
- Prasongsuk, S.; Lotrakul, P.; Ali, I.; Bankeeree, W.; Punnapayak, H. The current status of Aureobasidium pullulans in biotechnology. Folia Microbiol. 2018, 63, 129–140. [Google Scholar] [CrossRef]
- Santos, A.; Sánchez, A.; Marquina, D. Yeasts as biological agents to control Botrytis cinerea. Microbiol. Res. 2004, 159, 331–338. [Google Scholar] [CrossRef]
- Sansone, G.; Rezza, I.; Calvente, V.; Benuzzi, D.; Sanz De Tosetti, M.I. Control of Botrytis cinerea strains resistant to iprodione in apple with rhodotorulic acid and yeasts. Postharvest. Biol. Technol. 2005, 35, 245–251. [Google Scholar] [CrossRef]
- Belda, I.; Ruiz, J.; Alonso, A.; Marquina, D.; Santos, A. The Biology of Pichia membranifaciens Killer Toxins. Toxins 2017, 9, 112. [Google Scholar] [CrossRef] [Green Version]
- Heydari, A.; Pessarakli, M. A review on biological control of fungal plant pathogens using microbial antagonists. J. Biol. Sci. 2010, 10, 273–290. [Google Scholar] [CrossRef] [Green Version]
- Nagórska, K.; Bikowski, M.; Obuchowski, M. Multicellular behaviour and production of a wide variety of toxic substances support usage of Bacillus subtilis as a powerful biocontrol agent. Acta Biochim. Pol. 2007, 54, 495–508. [Google Scholar] [CrossRef] [Green Version]
- Stein, T. Bacillus subtilis antibiotics: Structures, syntheses and specific functions. Mol. Microbiol. 2005, 56, 845–857. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.; Huang, Z.; Chen, Y.; Zhou, L.; Chen, M.; Sun, J.; Lu, Z.; Lu, Y. Effect of combined Bacillomycin D and chitosan on growth of Rhizopus stolonifer and Botrytis cinerea and cherry tomato preservation. J. Sci. Food Agric. 2021, 101, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Borriss, R. Bacillus, a Plant-Beneficial Bacterium. In Principles of Plant-Microbe Interactions; Springer: Cham, Switzerland, 2015; pp. 379–391. [Google Scholar]
- Henry, G.; Deleu, M.; Jourdan, E.; Thonart, P.; Ongena, M. The bacterial lipopeptide surfactin targets the lipid fraction of the plant plasma membrane to trigger immune-related defence responses. Cell Microbiol. 2011, 13, 1824–1837. [Google Scholar] [CrossRef]
- Souto, G.I.; Correa, O.S.; Montecchia, M.S.; Kerber, N.L.; Pucheu, N.L.; Bachur, M.; García, A.F. Genetic and functional characterization of a Bacillus s strain excreting surfactin and antifungal metabolites partially identified as iturin-like compounds. J. Appl. Microbiol. 2004, 97, 1247–1256. [Google Scholar] [CrossRef]
- Chitarra, G.S.; Breeuwer, P.; Nout, M.J.; van Aelst, A.C.; Rombouts, F.M.; Abee, T. An antifungal compound produced by Bacillus subtilis YM 10-20 inhibits germination of Penicillium roqueforti conidiospores. J. Appl. Microbiol. 2003, 94, 159–166. [Google Scholar] [CrossRef]
- Hu, F.; Liu, Y.; Li, S. Rational strain improvement for surfactin production: Enhancing the yield and generating novel structures. Microb. Cell Factories 2019, 18, 42. [Google Scholar] [CrossRef] [Green Version]
- Nakkeeran, S.; Priyanka, R.; Rajamanickam, S.; Sivakumar, U. Bacillus amyloliquefaciens alters the diversity of volatile and non-volatile metabolites and induces the expression of defence genes for the management of Botrytis leaf blight of Lilium under protected conditions. J. Plant Pathol. 2020, 102, 1179–1189. [Google Scholar] [CrossRef]
- Mengistu, A.A. Endophytes: Colonization, Behaviour, and Their Role in Defense Mechanism. Int. J. Microbiol. 2020, 2020, 6927219. [Google Scholar] [CrossRef]
- Ait Barka, E.; Gognies, S.; Nowak, J.; Audran, J.C.; Belarbi, A. Inhibitory effect of endophyte bacteria on Botrytis cinerea and its influence to promote the grapevine growth. Biol. Control. 2002, 24, 135–142. [Google Scholar] [CrossRef]
- Tapwal, A.; Singh, U.; Teixeira, J.A.; Singh, G.; Garg, S.; Kumar, R. In Vitro Antagonism of Trichoderma viride against Five Phytopathogens. Pest Technol. 2011, 5, 59–62. [Google Scholar]
- Tiwari, S.; Prasad, V.; Lata, C. Bacillus: Plant Growth Promoting Bacteria For Sustainable Agriculture and Environment. In New and Future Developments in Microbial Biotechnology and Bioengineering: Microbial Biotechnology in Agro-Environmental Sustainability; Elsevier: London, UK, 2019; pp. 43–55. [Google Scholar]
- Carmona-Hernandez, S.; Reyes-Pérez, J.J.; Chiquito-Contreras, R.G.; Rincon-Enriquez, G.; Cerdan-Cabrera, C.R.; Hernandez-Montiel, L.G. Biocontrol of postharvest fruit fungal diseases by bacterial antagonists: A review. Agronomy 2019, 9, 121. [Google Scholar] [CrossRef] [Green Version]
- Page, M.G. The Role of Iron and Siderophores in Infection, and the Development of Siderophore Antibiotics. Clin. Infect. Dis. 2019, 69, 529–537. [Google Scholar] [CrossRef]
- Di Francesco, A.; Baraldi, E. How siderophore production can influence the biocontrol activity of Aureobasidium pullulans against Monilinia laxa on peaches. Biol. Control. 2021, 152, 104456. [Google Scholar] [CrossRef]
- Shanmugaiah, V.; Nithya, K.; Harikrishnan, H.; Jayaprakashvel, M.; Balasubramanian, N. Biocontrol Mechanisms of Siderophores against Bacterial Plant Pathogens. In Sustainable Approaches to Controlling Plant Pathogenic Bacteria; CRC Press: Boca Raton, USA, 2015; pp. 167–190. [Google Scholar]
- Oleńska, E.; Małek, W.; Wójcik, M.; Swiecicka, I.; Thijs, S.; Vangronsveld, J. Beneficial features of plant growth-promoting rhizobacteria for improving plant growth and health in challenging conditions: A methodical review. Sci. Total Environ. 2020, 743, 140682. [Google Scholar] [CrossRef]
- Crowley, D.E. Microbial Siderophores in the Plant Rhizosphere. In Iron Nutrition in Plants and Rhizospheric Microorganisms; Springer: Dordrecht, The Netherlands, 2006; pp. 169–198. [Google Scholar]
- Höfte, M. Classes of Microbial Siderophores. In Iron Chelation Plants Soil Microorg; Acad Press: New York, NY, USA, 1992; pp. 3–26. [Google Scholar]
- Crosa, J.H.; Walsh, C.T. Genetics and assembly line enzymology of siderophore biosynthesis in bacteria. Microbiol. Mol. Biol. Rev. 2002, 66, 223–249. [Google Scholar] [CrossRef] [Green Version]
- Saha, M.; Sarkar, S.; Sarkar, B.; Sharma, B.K.; Bhattacharjee, S.; Tribedi, P. Microbial siderophores and their potential applications: A review. Environ. Sci. Pollut. Res. Int. 2016, 23, 3984–3999. [Google Scholar] [CrossRef]
- Renshaw, J.C.; Robson, G.D.; Trinci, A.J.; Wiebe, M.G.; Livens, F.R.; Collison, D.; Taylor, R.J. Fungal siderophores: Structures, functions and applications. Mycol. Res. 2002, 106, 1123–1142. [Google Scholar] [CrossRef]
- Winkelmann, G. Structural and stereochemical aspects of iron transport in fungi. Biotechnol. Adv. 1990, 8, 207–231. [Google Scholar] [CrossRef]
- Welzel, K.; Eisfeld, K.; Antelo, L.; Anke, T.; Anke, H. Characterization of the ferrichrome A biosynthetic gene cluster in the homobasidiomycete Omphalotus olearius. FEMS Microbiol. Lett. 2005, 249, 157–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jalal, M.A.; Galles, J.L.; Van der Helm, D. Structure of des[diserylglycyl]ferrirhodin, DDF, a novel siderophore from Aspergillus ochraceous. J. Org. Chem. 2002, 50, 5642–5645. [Google Scholar] [CrossRef]
- Calvente, V.; Benuzzi, D.; De Tosetti, M.I. Antagonistic action of siderophores from Rhodotorula glutinis upon the postharvest pathogen Penicillium expansum. Int. Biodeterior. Biodegrad. 1999, 43, 167–172. [Google Scholar] [CrossRef]
- Choi, G.J.; Kim, J.C.; Jang, K.S.; Nam, M.H.; Lee, S.W.; Kim, H.T. Biocontrol activity of Acremonium strictum BCP against Botrytis diseases. Plant Pathol. J. 2009, 25, 165–171. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.S.; Gao, J.; Munir, I.; Zhang, M.; Liu, Y.; Moe, T.S.; Xue, J.; Zhang, X. Characterization of Endophytic Fungi, Acremonium s, from Lilium davidii and Analysis of Its Antifungal and Plant Growth-Promoting Effects. Biomed. Res. Int. 2021, 2021, 9930210. [Google Scholar] [CrossRef] [PubMed]
- Ferramola, M.; Benuzzi, D.; Calvente, V.; Sansone, G.; Cerutti, S.; Raba, J. The Use of Siderophores for Improving the Control Of Postharvest Diseases In Stored Fruits And Vegetables. In Microbial Pathogens and Strategies for Combating Them: Science, Technology and Education; Formatex Research Center: Badajoz, Spain, 2013. [Google Scholar]
- Sipiczki, M. Metschnikowia strains isolated from botrytized grapes antagonize fungal and bacterial growth by iron depletion. Appl. Environ. Microbiol. 2006, 72, 6716–6724. [Google Scholar] [CrossRef] [Green Version]
- Sipiczki, M. Metschnikowia pulcherrima and related pulcherrimin-producing yeasts: Fuzzy species boundaries and complex antimicrobial antagonism. Microorganisms 2020, 8, 1029. [Google Scholar] [CrossRef]
- Dave, B.; Kena Anshuman, H. Siderophores of halophilic archaea and their chemical characterization. Indian J. Exp. Biol. 2006, 44, 340–344. [Google Scholar]
- Payne, S.M.; Niesel, D.W.; Peixotto, S.S.; Lawlor, K.M. Expression of Hydroxamate and Phenolate Siderophores by Shigella flexneri. J. Bacteriol. 1983, 155, 949–955. [Google Scholar] [CrossRef] [Green Version]
- Dertz, E.A.; Stintzi, A.; Raymond, K.N. Siderophore-mediated iron transport in Bacillus subtilis and Corynebacterium glutamicum. JBIC J. Biol. Inorg. Chem. 2006, 11, 1087–1097. [Google Scholar] [CrossRef]
- Pal, K.K.; Tilak, K.V.; Saxena, A.K.; Dey, R.; Singh, C.S. Suppression of maize root diseases caused by Macrophomina phaseolina, Fusarium moniliforme and Fusarium graminearum by plant growth promoting rhizobacteria. Microbiol. Res. 2001, 156, 209–223. [Google Scholar] [CrossRef]
- Yu, X.; Ai, C.; Xin, L.; Zhou, G. The siderophore-producing bacterium, Bacillus subtilis CAS15, has a biocontrol effect on Fusarium wilt and promotes the growth of pepper. Eur. J. Soil Biol. 2011, 47, 138–145. [Google Scholar] [CrossRef]
- Dimopoulou, A.; Theologidis, I.; Benaki, D.; Koukounia, M.; Zervakou, A.; Tzima, A.; Diallinas, G.; Hatzinikolaou, D.G.; Skandalis, N. Direct Antibiotic Activity of Bacillibactin Broadens the Biocontrol Range of Bacillus amyloliquefaciens MBI600. mSphere 2021, 6, e00376-21. [Google Scholar] [CrossRef]
- Trapet, P.; Avoscan, L.; Klinguer, A.; Pateyron, S.; Citerne, S.; Chervin, C.; Mazurier, S.; Lemanceau, P.; Wendehenne, D.; Besson-Bard, A. The Pseudomonas fluorescens Siderophore Pyoverdine Weakens Arabidopsis thaliana Defense in Favor of Growth in Iron-Deficient Conditions. Plant Physiol. 2016, 171, 675–693. [Google Scholar] [CrossRef] [Green Version]
- Maindad, D.V.; Kasture, V.M.; Chaudhari, H.; Dhavale, D.D.; Chopade, B.A.; Sachdev, D. Characterization and Fungal Inhibition Activity of Siderophore from Wheat Rhizosphere Associated Acinetobacter calcoaceticus Strain HIRFA32. Indian J. Microbiol. 2014, 54, 315–322. [Google Scholar] [CrossRef] [Green Version]
- Gull, M.; Hafeez, F.Y. Characterization of siderophore producing bacterial strain Pseudomonas fluorescens Mst 8.2 as plant growth promoting and biocontrol agent in wheat. Afr. J. Microbiol. Res. 2012, 6, 6308–6318. [Google Scholar] [CrossRef] [Green Version]
- Smith, M.J.; Neilands, J.B. Rhizobactin, a siderophore from Rhizobium meliloti. J. Plant Nutr. 1984, 7, 449–458. [Google Scholar] [CrossRef]
- Meiwes, J.; Fiedler, H.; Haag, H.; Zähner, H.; Konetschny-Rapp, S.; Jung, G. Isolation and characterization of staphyloferrin A, a compound with siderophore activity from Staphylococcus hyicus DSM 20459. FEMS Microbiol. Lett. 1990, 55, 201–206. [Google Scholar] [CrossRef]
- Das, K.; Prasanna, R.; Saxena, A.K. Rhizobia: A potential biocontrol agent for soilborne fungal pathogens. Folia Microbiol. 2017, 62, 425–435. [Google Scholar] [CrossRef]
- De Meyer, G.; Höfte, M. Salicylic acid produced by the rhizobacterium Pseudomonas aeruginosa 7NSK2 induces resistance to leaf infection by Botrytis cinerea on bean. Phytopathology 1997, 87, 588–593. [Google Scholar] [CrossRef] [Green Version]
- Audenaert, K.; Pattery, T.; Cornelis, P.; Höfte, M. Induction of systemic resistance to Botrytis cinerea in tomato by Pseudomonas aeruginosa 7NSK2: Role of salicylic acid, pyochelin, and pyocyanin. Mol. Plant-Microbe Interact. 2002, 15, 1147–1156. [Google Scholar] [CrossRef] [Green Version]
- Meziane, H.; van der Sluis, I.; van Loon, L.C.; Höfte, M.; Bakker, A.H. Determinants of Pseudomonas putida WCS358 involved in inducing systemic resistance in plants. Mol. Plant Pathol. 2005, 6, 177–185. [Google Scholar] [CrossRef]
- Schulz-Bohm, K.; Martín-Sánchez, L.; Garbeva, P. Microbial volatiles: Small molecules with an important role in intra- and inter-kingdom interactions. Front. Microbiol. 2017, 8, 2484. [Google Scholar] [CrossRef]
- Gao, Z.; Zhang, B.; Liu, H.; Han, J.; Zhang, Y. Identification of endophytic Bacillus velezensis ZSY-1 strain and antifungal activity of its volatile compounds against Alternaria solani and Botrytis cinerea. Biol. Control 2017, 105, 27–39. [Google Scholar] [CrossRef]
- Chaouachi, M.; Marzouk, T.; Jallouli, S.; Elkahoui, S.; Gentzbittel, L.; Ben, C.; Djébali, N. Activity assessment of tomato endophytic bacteria bioactive compounds for the postharvest biocontrol of Botrytis cinerea. Postharvest. Biol. Technol. 2021, 172, 111389. [Google Scholar] [CrossRef]
- Wang, C.; Wang, Y.; Wang, L.; Fan, W.; Zhang, X.; Chen, X.; Wang, M.; Wang, J. Biocontrol potential of volatile organic compounds from Pseudomonas chlororaphis ZL3 against postharvest gray mold caused by Botrytis cinerea on Chinese cherry. Biol. Control 2021, 159, 104613. [Google Scholar] [CrossRef]
- Prigigallo, M.I.; De Stradis, A.; Anand, A.; Mannerucci, F.; L’Haridon, F.; Weisskopf, L.; Bubici, G. Basidiomycetes Are Particularly Sensitive to Bacterial Volatile Compounds: Mechanistic Insight into the Case Study of Pseudomonas protegens Volatilome against Heterobasidion abietinum. Front. Microbiol. 2021, 12, 1382. [Google Scholar] [CrossRef]
- Simionato, A.S.; Navarro, M.O.; de Jesus, M.L.; Barazetti, A.R.; da Silva, C.S.; Simões, G.C.; Balbi-Peña, M.I.; de Mello, J.C.; Panagio, L.A.; de Almeida, R.S.; et al. The Effect of Phenazine-1-Carboxylic Acid on Mycelial Growth of Botrytis cinerea Produced by Pseudomonas aeruginosa LV Strain. Front. Microbiol. 2017, 8, 1102. [Google Scholar] [CrossRef]
- Zhong, T.; Wang, Z.; Zhang, M.; Wei, X.; Kan, J.; Zalán, Z.; Wang, K.; Du, M. Volatile organic compounds produced by Pseudomonas fluorescens ZX as potential biological fumigants against gray mold on postharvest grapes. Biol. Control. 2021, 163, 104754. [Google Scholar] [CrossRef]
- Ayed, A.; Kalai-Grami, L.; Slimene, I.; Chaouachi, M.; Mankai, H.; Karkouch, I.; Djebali, N.; Elkahoui, S.; Tabbene, O.; Limam, F. Antifungal activity of volatile organic compounds from Streptomyces s strain S97 against Botrytis cinerea. Biocontrol. Sci. Technol. 2021, 31, 1330–1348. [Google Scholar] [CrossRef]
- Miles, L.A.; Lopera, C.A.; González, S.; de García, M.C.; Franco, A.E.; Restrepo, S. Exploring the biocontrol potential of fungal endophytes from an Andean Colombian Paramo ecosystem. BioControl 2012, 57, 697–710. [Google Scholar] [CrossRef]
- Haidar, R.; Amira, Y.; Roudet, J.; Marc, F.; Patrice, R. Application methods and modes of action of Pantoea agglomerans and Paenibacillus sp. to control the grapevine trunk disease-pathogen, Neofusicoccum parvum. OENO One 2021, 55, 1–16. [Google Scholar] [CrossRef]
- De Simone, N.; Capozzi, V.; de Chiara, M.L.; Amodio, M.L.; Brahimi, S.; Colelli, G.; Drider, D.; Spano, G.; Russo, P. Screening of lactic acid bacteria for the bio-control of botrytis cinerea and the potential of Lactiplantibacillus plantarum for eco-friendly preservation of fresh-cut kiwifruit. Microorganisms 2021, 9, 773. [Google Scholar] [CrossRef]
- Chaudhari, S.S.; Gokhale, D.V. Phenyllactic acid: A potential aantimicrobial compound in lactic acid bacteria. J. Bacteriol. Mycol. 2016, 2, 121–125. [Google Scholar]
- Tomsheck, A.R.; Strobel, G.A.; Booth, E.; Geary, B.; Spakowicz, D.; Knighton, B.; Floerchinger, C.; Sears, J.; Liarzi, O.; Ezra, D. Hypoxylon sp., an endophyte of Persea indica, producing 1,8-Cineole and Other Bioactive Volatiles with Fuel Potential. Microb. Ecol. 2010, 60, 903–914. [Google Scholar] [CrossRef]
- Guigón-López, C.; Holguín-Ibarra, D.; Torres-Zapien, J.H.; García- Cruz, I.; Villapando, I.; Salas-Salazar, N.A. Metarhizium anisopliae reduces conidial germination and mycelium growth of the apple gray mold Botrytis cinerea. Biol. Control. 2021, 160, 104660. [Google Scholar] [CrossRef]
- Singh, S.K.; Strobel, G.A.; Knighton, B.; Geary, B.; Sears, J.; Ezra, D. An Endophytic Phomopsis s Possessing Bioactivity and Fuel Potential with its Volatile Organic Compounds. Microb. Ecol. 2011, 61, 729–739. [Google Scholar] [CrossRef]
- Fu, J.; Zhou, Y.; Li, H.F.; Ye, Y.H.; Guo, J.H. Antifungal metabolites from Phomopsis s By254, an endophytic fungus in Gossypium hirsutum. Afr. J. Microbiol. Res. 2011, 5, 1231–1236. [Google Scholar]
- Di Francesco, A.; Zajc, J.; Gunde-Cimerman, N.; Aprea, E.; Gasperi, F.; Placì, N.; Caruso, F.; Baraldi, E. Bioactivity of volatile organic compounds by Aureobasidium species against gray mold of tomato and table grape. World J. Microbiol. Biotechnol. 2020, 36, 171. [Google Scholar] [CrossRef]
- Huang, R.; Che, H.J.; Zhang, J.; Yang, L.; Jiang, D.H.; Li, G.Q. Evaluation of Sporidiobolus pararoseus strain YCXT3 as biocontrol agent of Botrytis cinerea on post-harvest strawberry fruits. Biol. Control 2012, 62, 53–63. [Google Scholar] [CrossRef]
- Huang, R.; Li, G.Q.; Zhang, J.; Yang, L.; Che, H.J.; Jiang, D.H.; Huang, H.C. Control of Postharvest Botrytis Fruit Rot of Strawberry by Volatile Organic Compounds of Candida intermedia. Phytopathology 2011, 101, 859–869. [Google Scholar] [CrossRef] [Green Version]
- Zou, X.; Wei, Y.; Dai, K.; Xu, F.; Wang, H.; Shao, X. Yeasts from intertidal zone marine sediment demonstrate antagonistic activities against Botrytis cinerea in vitro and in strawberry fruit. Biol. Control 2021, 158, 104612. [Google Scholar] [CrossRef]
- Kanchiswamy, C.N.; Malnoy, M.; Maffei, M.E. Chemical diversity of microbial volatiles and their potential for plant growth and productivity. Front. Plant Sci. 2015, 6, 151. [Google Scholar] [CrossRef] [Green Version]
- Insam, H.; Seewald, M.S. Volatile organic compounds [VOCs] in soils. Biol. Fertil. Soils 2010, 46, 199–213. [Google Scholar] [CrossRef] [Green Version]
- Korpi, A.; Järnberg, J.; Pasanen, A.L. Microbial volatile organic compounds. Crit. Rev. Toxico. 2009, 39, 139–193. [Google Scholar] [CrossRef]
- Nicot, C.; Stewart, A.; Bardin, M.; Elad, Y. Biological Control and Biopesticide Suppression of Botrytis-Incited Diseases. In Botrytis—The Fungus, the Pathogen and its Management in Agricultural Systems; Springer International Publishing: London, UK, 2015; pp. 165–187. [Google Scholar]
- Gow, N.A.; Latge, J.P.; Munro, C.A. The Fungal Cell Wall: Structure, Biosynthesis, and Function. In The Fungal Kingdom; American Society of Microbiology: Washington, DC, USA, 2017; pp. 267–292. [Google Scholar]
- Latgé, J. The cell wall: A carbohydrate armour for the fungal cell. Mol. Microbiol. 2007, 66, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Cantu, D.; Greve, L.C.; Labavitch, J.M.; Powell, A.L. Characterization of the cell wall of the ubiquitous plant pathogen Botrytis cinerea. Mycol. Res. 2009, 113, 1396–1403. [Google Scholar] [CrossRef]
- Daulagala, W.H. Chitinolytic Endophytic Bacteria as Biocontrol Agents for Phytopathogenic Fungi and Nematode Pests: A Review. Asian J. Res. Bot. 2021, 5, 14–24. [Google Scholar]
- Maung, C.E.; Baek, W.S.; Choi, T.G.; Kim, K.Y. Control of grey mould disease on strawberry using the effective agent, Bacillus amyloliquefaciens Y1. Biocontrol. Sci. Technol. 2021, 31, 468–482. [Google Scholar] [CrossRef]
- Wang, F.; Xiao, J.; Zhang, Y.; Li, R.; Liu, L.; Deng, J. Biocontrol ability and action mechanism of Bacillus halotolerans against Botrytis cinerea causing grey mould in postharvest strawberry fruit. Postharvest. Biol. Technol. 2021, 174, 111456. [Google Scholar] [CrossRef]
- Lu, Y.; Ma, D.; He, X.; Wang, F.; Wu, J.; Liu, Y.; Jiaoa, J.; Deng, J. Bacillus subtilis KLBC BS6 induces resistance and defence-related response against Botrytis cinerea in blueberry fruit. Physiol. Mol. Plant Pathol. 2021, 114, 101599. [Google Scholar] [CrossRef]
- Zhang, W.; Ma, J.; Yan, Q.; Jiang, Z.; Yang, S. Biochemical characterization of a novel acidic chitinase with antifungal activity from Paenibacillus xylanexedens Z2–4. Int. J. Biol. Macromol. 2021, 182, 1528–1536. [Google Scholar] [CrossRef]
- Frankowski, J.; Lorito, M.; Scala, F.; Schmid, R.; Berg, G.; Bahl, H. Purification and properties of two chitinolytic enzymes of Serratia plymuthica HRO-C48. Arch. Microbiol. 2001, 176, 421–426. [Google Scholar] [CrossRef] [PubMed]
- Essghaier, B.; Fardeau, M.L.; Cayol, J.L.; Hajlaoui, M.R.; Boudabous, A.; Jijakli, H.; Sadfi-Zouaoui, N. Biological control of grey mould in strawberry fruits by halophilic bacteria. J. Appl. Microbiol. 2009, 106, 833–846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Chang, P.; Gao, L.; Wang, X. The Endophytic Fungus Albifimbria verrucaria from Wild Grape as an Antagonist of Botrytis cinerea and Other Grape Pathogens. Phytopathology 2020, 110, 843–850. [Google Scholar] [CrossRef] [PubMed]
- Pachenari, A.; Dix, N.J. Production of toxins and wall degrading enzymes by Gliocladium roseum. Trans. Br. Mycol. Soc. 1980, 74, 561–566. [Google Scholar] [CrossRef]
- Lorito, M.; Hayes, C.K.; Zoina, A.; Scala, F.; Del Sorbo, G.; Woo, S.L.; Harman, G.E. Potential of genes and gene products from Trichoderma s and Gliocladium s for the development of biological pesticides. Mol. Biotechnol. 1994, 2, 209–217. [Google Scholar] [CrossRef]
- Elad, Y.; Kapat, A. The Role of Trichoderma harzianum Protease in the Biocontrol of Botrytis cinerea. Eur. J. Plant Pathol. 1999, 105, 177–189. [Google Scholar] [CrossRef]
- Heyens, K.; Aerts, R.; Seels, B.; Vogels, L.; Bruno, A. Biological control of Botrytis cinerea in tomato. Acta Hortic. 2011, 914, 361–363. [Google Scholar] [CrossRef]
- Zhang, D.; Spadaro, D.; Valente, S.; Garibaldi, A.; Gullino, M.L. Cloning, characterization, expression and antifungal activity of an alkaline serine protease of Aureobasidium pullulans PL5 involved in the biological control of postharvest pathogens. Int. J. Food Microbiol. 2012, 153, 453–464. [Google Scholar] [CrossRef]
- Friel, D.; Gomez Pessoa, N.M.; Vandenbol, M.; Jijakli, M.H. Separate and combined disruptions of two exo-beta-1,3-glucanase genes decrease the efficiency of Pichia anomala [strain K] biocontrol against Botrytis cinerea on apple. Mol. Plant Microbe Interact. 2007, 20, 371–379. [Google Scholar] [CrossRef] [Green Version]
- Lima, G.; de Curtis, F.; Piedimonte, D.; Spina, A.M.; de Cicco, V. Integration of biocontrol yeast and thiabendazole protects stored apples from fungicide sensitive and resistant isolates of Botrytis cinerea. Postharvest Biol. Technol. 2006, 40, 301–307. [Google Scholar] [CrossRef]
- Kredics, L.; Antal, Z.; Szekeres, A.; Hatvani, L.; Manczinger, L.; Vágvölgyi, C.; Nagy, E. Extracellular proteases of Trichoderma species: A review. Acta Microbiol. Immunol. Hung. 2005, 52, 169–184. [Google Scholar] [CrossRef]
- De Simone, N.; Pace, B.; Grieco, F.; Chimienti, M.; Tyibilika, V.; Santoro, V.; Capozzi, V.; Colelli, G.; Spano, G.; Russo, P. Botrytis cinerea and Table Grapes: A Review of the Main Physical, Chemical, and Bio-Based Control Treatments in Post-Harvest. Foods 2020, 9, 1138. [Google Scholar] [CrossRef]
- Salvatierra-Martinez, R.; Arancibia, W.; Araya, M.; Aguilera, S.; Olalde, V.; Bravo, J.; Stoll, A. Colonization ability as an indicator of enhanced biocontrol capacity—An example using two Bacillus amyloliquefaciens strains and Botrytis cinerea infection of tomatoes. J. Phytopathol. 2018, 166, 601–612. [Google Scholar] [CrossRef]
- Zhou, C.; Zhu, J.; Qian, N.; Guo, J.; Yan, C. Bacillus subtilis SL18r Induces Tomato Resistance Against Botrytis cinerea, Involving Activation of Long Non-coding RNA, MSTRG18363, to Decoy miR1918. Front. Plant Sci. 2021, 11, 2317. [Google Scholar] [CrossRef]
- Miotto-Vilanova, L.; Jacquard, C.; Courteaux, B.; Wortham, L.; Michel, J.; Clément, C.; Barka, E.A.; Sanchez, L. Burkholderia phytofirmans PsJN Confers Grapevine Resistance against Botrytis cinerea via a Direct Antimicrobial Effect Combined with a Better Resource Mobilization. Front. Plant Sci. 2016, 7, 1236. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Cao, Z.; Li, J.; Tao, C.; Feng, Y.; Han, Y. A novel endophytic strain of Lactobacillus plantarum CM-3 with antagonistic activity against Botrytis cinerea on strawberry fruit. Biol. Control 2020, 148, 104306. [Google Scholar] [CrossRef]
- Gasser, F.; Cardinale, M.; Schildberger, B.; Berg, G. Biocontrol of Botrytis cinerea by successful introduction of Pantoea ananatis in the grapevine phyllosphere. Int. J. Wine Res. 2012, 4, 53–63. [Google Scholar]
- Van Loon, L.C. Induced resistance in plants and the role of pathogenesis-related proteins. Eur. J. Plant Pathol. 1997, 103, 753–765. [Google Scholar] [CrossRef]
- Islam, A.; Kabir, S.; Khair, A. Characterization and evaluation of Bacillus siamensis isolate for its growth promoting potential in tomato. Agriculture 2019, 65, 42–50. [Google Scholar] [CrossRef]
- Mari, M.; Guizzardi, M.; Pratella, G.C. Biological control of gray mold in pears by antagonistic bacteria. Biol. Control. 1996, 7, 30–37. [Google Scholar] [CrossRef]
- Sutton, J.C.; Peng, G. Biocontrol of Botrytis cinerea in Strawberry Leaves. Phytopathology 1993, 83, 615–621. [Google Scholar] [CrossRef]
- Liu, C.M.; Liu, S.Y.; Liao, C.K.; Lo, C.T.; Lin, K.C.; Peng, K.C. Cabbage defense response provoked by Trichoderma Th-LAAO. Arch. Microbiol. 2021, 203, 1641–1647. [Google Scholar] [CrossRef]
- Fiori, S.; Fadda, A.; Giobbe, S.; Berardi, E.; Migheli, Q. Pichia angusta is an effective biocontrol yeast against postharvest decay of apple fruit caused by Botrytis cinerea and Monilia fructicola. FEMS Yeast Res. 2008, 8, 961–963. [Google Scholar] [CrossRef] [Green Version]
- Hamdache, A.; Ezziyyani, M.; Lamarti, A. Effect of preventive and simultaneous inoculations of Bacillus amyloliquefaciens [Fukumoto] strains on conidial germination of Botrytis cinerea Pers. Fr. An Biol. 2018, 40, 65–72. [Google Scholar] [CrossRef]
- Raynaldo, F.A.; Dhanasekaran, S.; Ngea, G.L.; Yang, Q.; Zhang, X.; Zhang, H. Investigating the biocontrol potentiality of Wickerhamomyces anomalus against postharvest gray mold decay in cherry tomatoes. Sci. Hortic. 2021, 285, 110137. [Google Scholar] [CrossRef]
- Ingold, C.T.; Hudson, H.J.; Ingold, C.T.; Hudson, H.J. Fungi as Plant Pathogens. In The Biology of Fungi; Springer: Dordrecht, The Netherlands, 1993; pp. 159–182. [Google Scholar]
- Hernández-Fernández, M.; Cordero-Bueso, G.; Ruiz-Muñoz, M.; Cantoral, J.M. Culturable yeasts as biofertilizers and biopesticides for a sustainable agriculture: A comprehensive review. Plants 2021, 10, 822. [Google Scholar] [CrossRef]
- Hilber-Bodmer, M.; Schmid, M.; Ahrens, C.H.; Freimoser, F.M. Competition assays and physiological experiments of soil and phyllosphere yeasts identify Candida subhashii as a novel antagonist of filamentous fungi. BMC Microbiol. 2017, 17, 4. [Google Scholar] [CrossRef] [Green Version]
- Kloepper, J.W.; Schippers, B.; Bakker, A.H. Proposed Elimination of the Term Endorhizosphere. Phytopathology 1992, 82, 726–727. [Google Scholar]
- Ferrari, S.; Galletti, R.; Denoux, C.; De Lorenzo, G.; Ausubel, F.M.; Dewdney, J. Resistance to Botrytis cinerea induced in arabidopsis by elicitors is independent of salicylic acid, ethylene, or jasmonate signaling but requires Phytoalexin Deficient3. Plant Physiol. 2007, 144, 367–379. [Google Scholar] [CrossRef] [Green Version]
- Govrin, E.M.; Levine, A. Infection of Arabidopsis with a necrotrophic pathogen, Botrytis cinerea, elicits various defense responses but does not induce systemic acquired resistance [SAR]. Plant Mol. Biol. 2002, 48, 267–276. [Google Scholar] [CrossRef]
- Vallad, G.E.; Goodman, R.M. Systemic acquired resistance and induced systemic resistance in conventional agriculture. Crop Sci. 2004, 44, 1920–1934. [Google Scholar] [CrossRef] [Green Version]
- Peng, K.C.; Lin, C.C.; Liao, C.F.; Yu, H.C.; Lo, C.T.; Yang, H.H.; Lin, K.C. Expression of L-amino acid oxidase of Trichoderma harzianum in tobacco confers resistance to Sclerotinia sclerotiorum and Botrytis cinerea. Plant Sci. 2021, 303, 110772. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Antifungal Microorganisms | Condition/Plant | Antifungal Metabolite | Antifungal Effect | Reference |
|---|---|---|---|---|
| Bacteria | ||||
| Actinoalloteichus cyanogriseus 12A22 | In vitro | 2-Hydroxyethyl-3-methyl-1,4-naphthoquinone | Growth inhibition | [70] |
| Bacillus amyloliquefaciens VB7 | In vitro and foliar application | Phthalic acid, hept-3-yl isobutyl ester and propanoic acid,2-hydroxy-, methyl ester | Conidia parasitationSuppression of mycelial growth | [71] |
| Bacillus subtilis NCD-2 | Apple fruit | Fengycin | Open pores in the plasma membrane | [72] |
| Bacillus velezensis Bvel1 | Pepper and grape plants | Bacillibactin | Suppression of fungal growth by chelating the available ferric iron | [73] |
| Bacillus velezensis XT1 | In vitro and in fruits (Tomatoes, grapes, strawberries) | Surfactin, fengycin, and bacillomycin | Open pores in the plasma membrane | [56] |
| Kosakonia radicincitans DSM 16656 | In vitro and in apple fruit | Enterochelin | Blocking the polygalacturonase | [74] |
| Ochrobactrum cicero MM17 | In vitro and Lilium L. | Propanoic acid, −hydoxy-methyl ester; phthalic acid, hex-3-yl isobutyl ester and phthalic acid, hept-3-yl isobutyl ester | Suppression of mycelial growth | [75] |
| Pantoea sp. MQT16M1 | In vitro and in strawberry fruits | Salicylamide, maculosin, and herniarin | Disruption of cell wall components | [76] |
| Paraburkholderia phytofirmans PsJN (Pseudomonas sp. PsJN) | In vitro | Lipopolysaccharides | Cytoplasm coagulation | [71] |
| Pseudomonas aeruginosa 7NSK2 | Tomato plants and Arabidopsis thaliana | Pyochelin and pyocyanin | Induction of systemic resistance (ISR) | [77] |
| Pseudomonas fluorescens fp-5 | Strawberry plants | Hydroxamate-type siderophores | Prevention of plant infection | [78] |
| Pseudomonas sp. CHA0 | Soil | Pyoverdine or pseudobactin | Iron depletion | [50] |
| Rahnella aquatilis BNM | In vitro | Enterochelin | Blockage of polygalacturonase | [79] |
| Fungi | ||||
| Acremonium persicinum MF-347833 | In vitro | VL-2397 (cyclic hexapeptide) | Suppression of hyphal elongation | [80] |
| Aspergillus fumigatus AF293 | In vitro | Gliotoxin | DNA disruption | [81] |
| Gliocladium virens 41 | In vitro | Gliotoxin | Inhibition of spore germination | [82] |
| Trichoderma atroviride SJ3-4 | In vitro and in Phaseolus vulgaris cv. Borlotto seeds | Glucose oxidase dermadin, trichovirdin and sesquiterpene, heptalic acid | Effect on spore germination | [83] |
| Trichoderma sp. BV1 | Rubus sp. | Gliotoxin | Suppression of conidial germination | [84] |
| Yeasts | ||||
| Aureobasidium pullulans L47 | Post-harvest grape berries, kiwi fruit, and strawberries | Aureobasidins | Inhibition of inositol phosphoryl ceramide synthase | [85] |
| Metschnikowia pulcherrima MPR3 | In vitro | Pulcherrimin | Iron sequestration | [64] |
| Pichia membranifaciens CYC 1106 | Apple fruits | Killer toxin | Damage on membrane, glucanase activity, inhibition of β-1,3-glucansynthase, cell cycle arrestation, and inhibition of calcium uptake | [86] |
| Rhodotorula glutinis ySL 30 | In vitro | Rhodotorulic acid | Inhibition of polygalacturonase and laccase | [87] |
| Saccharomyces cerevisiae CBS8112 | Post-harvest pears | Killer toxin | Damage to the membrane, glucanase activity, inhibition of β-1,3-glucansynthase, cell cycle arrestation, and inhibition of calcium uptake | [88] |
| Antifungal Microorganisms | Condition/Plant | Antifungal Metabolite | Antifungal Effect | Reference |
|---|---|---|---|---|
| Bacteria | ||||
| Bacillus amyloliquefaciens VB7 | In vitro and foliar application | Phthalic acid, hept-3-yl isobutyl ester and propanoic acid,2-hydroxy-, methyl ester | Conidia parasitation Suppression of mycelial growth | [98] |
| Bacillus nakamurai TR2, Bacillus pseudomycoides DHT2, Bacillus proteolyticus H2F1 and Bacillus thuringiensis H1R2 | In vitro and in organic tomato fruits | 3-methylbutan-1-ol, sulfur-containing compounds, 2-heptanone, and dodecanal | Suppression of mycelial growth | [139] |
| Bacillus velezensis ZSY-1 | In vitro | Pyrazine [2,5-dimethyl], benzothiazole, phenol (4-chloro-3-methyl), and phenol-2,4-bis (1,1-dimethylethyl) | Suppression of mycelial growth and sporulation | [138] |
| Lactiplantibacillus plantarum UFG 121 | In vitro and in kiwifruits | Phenyllactic acid and 4-hydroxyphenyllactic acid | Suppression of mycelial growth | [147] |
| Pantoea sp. MQT16M1 | Grapevine plants | Phenylethyl alcohol | Reduction of the length of internal necrosis | [76] |
| Pseudomonas aeruginosa LV | In vitro | Phenazine-1-carboxylic acid | Suppression of mycelial growth | [142] |
| Pseudomonas chlororaphis ZL3 | In vitro and in Chinese cherry | 1-dodecene and dimethyl disulfide | Reduction of disease incidence and lesion diameter | [140] |
| Pseudomonas fluorescens ZX | In vivo and in grapes | Dimethyl trisulfide and geranyl formate | Suppression of mycelial growth and spore germination. Reduction of disease incidence and the disease index | [143] |
| Pseudomonas protegens CHAo | In vitro | Ammonia and dimethyl trisulfide | Suppression of mycelial growth | [141] |
| Streptomyces sp. S97 | In vitro and in strawberries | 3-carene 2,5-dione, geosmin, beta-cubebene, and one phenolic compound | Inhibition of Botrytis cinerea decay on strawberries and suppression of germination | [144] |
| Fungi | ||||
| Hypoxylon sp. CI-4 | In vitro | 1,8-cineole, 1-methyl-1,4-cyclohexadiene | Suppression of mycelial growth | [149] |
| Metarhizium anisopliae Ma70 | In vitro and in apple fruits | 1-octanol | Suppression of conidia germination and mycelium growth | [150] |
| Phomopsis sp. By 254 | In vitro | Sabinene, 1-butanol, 3-methyl; benzene ethanol; 1-propanol, 2-methyl, and 2-propanone | Suppression of mycelial growth | [152] |
| Trichoderma spp. | In vitro | Trichodermol, harzianum A, and harzianolide | Suppression of mycelial growth | [101] |
| Yeasts | ||||
| Aureobasidium pullulans EXF-6519 | In vitro and in tomato fruits and grapes | 3-methyl-1-butanol | Suppression of mycelial growth and reduction of fungal incidence | [153] |
| Candida intermedia C410 | In vitro and in strawberry fruits | 1, 3, 5, 7-cyclooctatetraene, 3-methyl- 1-butanol, 2-nonanone, and phenylethyl alcohol | Suppression spore germination and mycelial growth | [155] |
| Scheffersomyces spartinae W9, Candida pseudolambica W16 | In vitro and in strawberry fruits in planta | Unknown | Suppression of mycelial growth and reduction of disease incidence in fruits | [156] |
| Sporidiobolus pararoseus YCXT3 | In vitro and in strawberry fruits | 2-ethyl-1-hexanol | Suppression of spore germination and mycelial growth | [154] |
| Antifungal Microorganisms | Condition/Plant | Antifungal Metabolite | Antifungal Effect | Reference |
|---|---|---|---|---|
| Bacteria | ||||
| Bacillus amyloliquefaciens Y1 | In vitro | β-1,3-glucanase | Suppression of mycelial growth. Modification of the hyphal structure | [165] |
| Bacillus halotolerans KLBC XJ-5 | In vitro and in strawberry fruits | Chitinase and β-1, 3-glucanase | Suppression of mycelial growth and reduction of conidial germination | [166] |
| Bacillus subtilis KLBC BS6 | In vitro and in blueberry fruits | Chitinase | Suppression of mycelial growth and reduction of conidial germination | [167] |
| Paenibacillus xylanexedens Z2–4 | In vitro | Chitinase | Suppression of mycelial growth | [168] |
| Pseudomonas elgii HOA73 | In vitro | Chitinase | Suppression of spore germination | [34] |
| Serratia plymuthica C48 | In vitro | Chitinase | Suppression of spore germination and germ-tube elongation | [169] |
| Virgibacillus marismortui M3-23, Terribacillus halophilus J31, Halomonas elongate L80, Planococcus rifietoensis M2-26, Staphylococcus equorum B1-35 and Staphylococcus sp. J23 | In vitro and in strawberry fruits | Chitinase, β-1,3-glucanase, cellulase and protease | Reduction of gray mold rot incidence and fungal growth | [170] |
| Fungi | ||||
| Albifimbria verrucaria SYE-1 | In vitro and in grape leaves | Chitinase | Suppression of conidium germination and mycelial growth | [171] |
| Gliocladium roseum Bainier | In vitro | β l-3 glucanase | Breakdown of glucans in hyphal walls | [172] |
| Gliocladium virens 41 | In vitro | Endochitinase | Suppression of spore germination and hyphal elongation | [173] |
| Trichoderma harzianum NCIM1185 | Bean leaves | Extracellular proteases | Inactivation of fungal hydrolytic enzymes and attack of fungal cell wall components | [174] |
| Trichoderma harzianum T39 | Bean leaves | Exo- and endo-polygalacturonase, pectin methyl esterase, pectate lyase, cutinase, chitinase, and β-1,3-glucanase | Reduction of spore germination | [175] |
| Yeasts | ||||
| Aureobasidium pullulans PI1 | In vitro and in grape and mandarin fruits | β-1,3-glucanase, pectinase, and protease | Disruption of cell wall components and competition for nutrients | [63] |
| Aureobasidium pullulans PL5 | In vitro | Alkaline serine protease | Reduction of spore germination and germ-tube length | [176] |
| Galactomyces candidum JYC1146 | In vitro and in strawberry fruits | Chitinase | Control of fungal growth and reduction of disease severity | [19] |
| Pichia anomala K | Apple fruits | Exo-β-glucanases | Disruption of cell wall components | [177] |
| Cryptococcus laurentii LS28 | Apple fruits | β-1,3-glucanase | Reduction of disease appearance in postharvest conditions | [178] |
| Antifungal Microorganisms | Condition/Plant | Antifungal Metabolite | Antifungal Effect | Reference |
|---|---|---|---|---|
| Bacteria | ||||
| Bacillus amyloliquefaciens BBC047 | Tomato leaves | - | Biofilm formation | [181] |
| Bacillus subtilis FB17 | Tomato plants | - | Induction of systemic resistance [ISR] | [182] |
| Bacillus velezensis Bvel1 | Arabidopsis thaliana | Azelaic acid | Induction of systemic resistance [ISR] | [73] |
| Burkholderia phytofirmans PsJN | Grapevines | H2O2 production | Induction of systemic resistance [ISR] | [183] |
| Lactobacillus plantarum CM-3 | Strawberry wounds | - | Colonization | [184] |
| Pantoea ananatis BLBT1-08 | Grapevine leaves | - | Colonization | [185] |
| Pseudomonas aeruginosa 7NSK2, Pseudomonas fluorescens CHA0, Pseudomonas aeruginosa 7NSK2, and Serratia marcescens 90-166 | Bean plants | Salicylic acid | Induction of systemic resistance [ISR] | [186,187] |
| Pseudomonas syringae L-59-66 | Pear fruits | - | Competition for nutrients | [188] |
| Fungi | ||||
| Aureobasidium pullulans L1 | In vitro and in peach fruits | Hydroxamate-type siderophore | Iron competition | [105] |
| Gliocladium roseum | Strawberry leaves | - | Nutrient competition | [189] |
| Trichoderma arundinaceum IBT 40837 | Tomato plants | Trichodiene | Induction of defense-related genes | [99] |
| Trichoderma harzianum Th-LAAO | Tobacco leaves | L-amino acid oxidase | Induction of defense-related genes | [190] |
| Trichoderma harzianum T39 and Ulocladium atrum 385 | Laboratory conditions | - | Competition for nutrients and colonization of necrotic tissue | [58] |
| Yeasts | ||||
| Pichia angusta ANY-67 | Apple fruit | - | Wound protection | [191] |
| Rhodotorula glutinis F147 and Cryptococcus albidus F131 | In vitro | - | Competition for iron, methionine, leucine, and other nutrients | [192] |
| Candida saitoana 240, Candida oleophila 182, and Metschnikowia fructicola NRRL Y-30752 | Fruits | Overproduction of reactive oxygen species | Induction of innate immune responses | [65] |
| Wickerhamomyces anomalus YE06 | Cherry tomatoes | - | Competition for space and nutrients | [193] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roca-Couso, R.; Flores-Félix, J.D.; Rivas, R. Mechanisms of Action of Microbial Biocontrol Agents against Botrytis cinerea. J. Fungi 2021, 7, 1045. https://doi.org/10.3390/jof7121045
Roca-Couso R, Flores-Félix JD, Rivas R. Mechanisms of Action of Microbial Biocontrol Agents against Botrytis cinerea. Journal of Fungi. 2021; 7(12):1045. https://doi.org/10.3390/jof7121045
Chicago/Turabian StyleRoca-Couso, Rocío, José David Flores-Félix, and Raúl Rivas. 2021. "Mechanisms of Action of Microbial Biocontrol Agents against Botrytis cinerea" Journal of Fungi 7, no. 12: 1045. https://doi.org/10.3390/jof7121045
APA StyleRoca-Couso, R., Flores-Félix, J. D., & Rivas, R. (2021). Mechanisms of Action of Microbial Biocontrol Agents against Botrytis cinerea. Journal of Fungi, 7(12), 1045. https://doi.org/10.3390/jof7121045