
Host-Specific Effects of Arbuscular Mycorrhizal Fungi on Two Caragana Species in Desert Grassland

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Trap Culture of AMF
2.3. Seedling Preparation
2.4. Field Experiment Design
2.5. Field Measurements
2.6. DNA Extraction and Sequencing
2.7. Statistic Analysis
3. Result
3.1. AM Fungal Community Properties
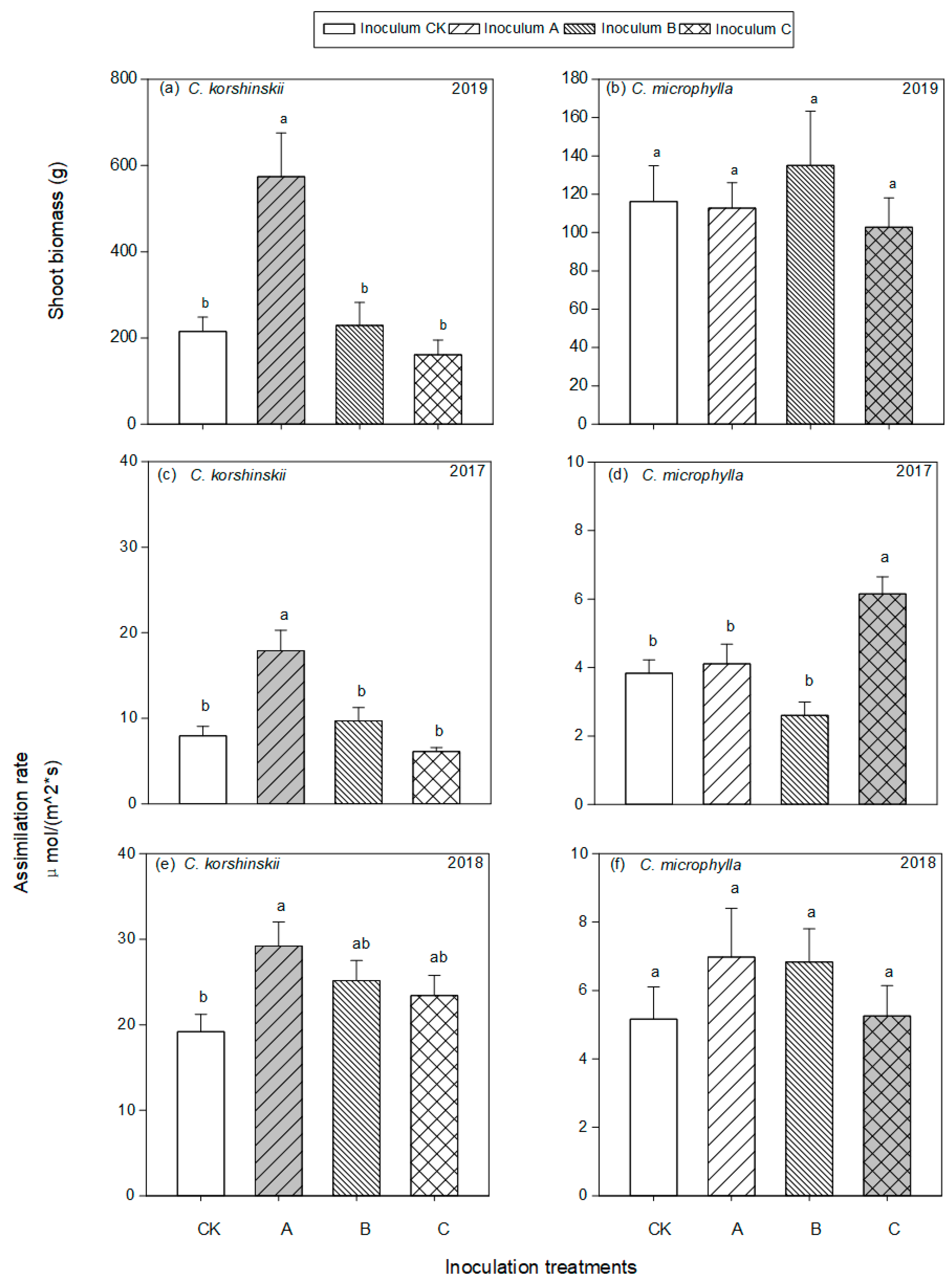
3.2. Plant Response to AMF Inoculation
3.3. The Association between AM Fungal Community and Plant Performance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brundrett, M.C.; Tedersoo, L. Evolutionary history of mycorrhizal symbioses and global host plant diversity. New Phytol. 2018, 220, 1108–1115. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: Oxford, UK, 2008. [Google Scholar]
- Koziol, L.; Schultz, P.A.; House, G.L.; Bauer, J.T.; Middleton, E.L.; Bever, J.D. The Plant Microbiome and Native Plant Restoration: The Example of Native Mycorrhizal Fungi. Bioscience 2018, 68, 996–1006. [Google Scholar] [CrossRef] [Green Version]
- Vahter, T.; Bueno, C.G.; Davison, J.; Herodes, K.; Hiiesalu, I.; Kasari-Toussaint, L.; Oja, J.; Olsson, P.A.; Sepp, S.K.; Zobel, M.; et al. Co-introduction of native mycorrhizal fungi and plant seeds accelerates restoration of post-mining landscapes. J. Appl. Ecol. 2020, 57, 1741–1751. [Google Scholar] [CrossRef]
- Moreno-Mateos, D.; Alberdi, A.; Morrien, E.; van der Putten, W.H.; Rodriguez-Una, A.; Montoya, D. The long-term restoration of ecosystem complexity. Nat. Ecol. Evol. 2020, 4, 676–685. [Google Scholar] [CrossRef]
- Reynolds, H.L.; Vogelsang, K.M.; Hartley, A.E.; Bever, J.D.; Schultz, P.A. Variable responses of old-field perennials to arbuscular mycorrhizal fungi and phosphorus source. Oecologia 2006, 147, 348–358. [Google Scholar] [CrossRef] [PubMed]
- Calonne-Salmon, M.; Plouznikoff, K.; Declerck, S. The arbuscular mycorrhizal fungus Rhizophagus irregularis MUCL 41833 increases the phosphorus uptake and biomass of Medicago truncatula, a benzo[a]pyrene-tolerant plant species. Mycorrhiza 2018, 28, 761–771. [Google Scholar] [CrossRef] [PubMed]
- Frey, B.; Schuepp, H. Acquisition of nitrogen by external hyphae of arbuscular mycorrhizal fungi associated with Zea mays L. New Phytol. 1993, 124, 221–230. [Google Scholar] [CrossRef]
- Volpe, V.; Chitarra, W.; Cascone, P.; Volpe, M.G.; Bartolini, P.; Moneti, G.; Pieraccini, G.; Di Serio, C.; Maserti, B.; Guerrieri, E.; et al. The Association with Two Different Arbuscular Mycorrhizal Fungi Differently Affects Water Stress Tolerance in Tomato. Front. Plant Sci. 2018, 9, 1480. [Google Scholar] [CrossRef]
- Paymaneh, Z.; Sarcheshmehpour, M.; Bukovská, P.; Jansa, J. Could indigenous arbuscular mycorrhizal communities be used to improve tolerance of pistachio to salinity and/or drought? Symbiosis 2019, 79, 269–283. [Google Scholar] [CrossRef]
- Pereyra, M.; Grilli, G.; Galetto, L. Root-associated fungi increase male fitness, while high simulated herbivory decreases indirect defenses in Croton lachnostachyus plants. Plant Ecol. 2018, 220, 29–39. [Google Scholar] [CrossRef]
- Kaur, J.; Chavana, J.; Soti, P.; Racelis, A.; Kariyat, R. Arbuscular mycorrhizal fungi (AMF) influences growth and insect community dynamics in Sorghum-sudangrass (Sorghum × drummondii). Arthropod-Plant Interact. 2020, 14, 301–315. [Google Scholar] [CrossRef]
- Cruz-Silva, A.; Figueiredo, A.; Sebastiana, M. First Insights into the Effect of Mycorrhizae on the Expression of Pathogen Effectors during the Infection of Grapevine with Plasmopara viticola. Sustainability 2021, 13, 1226. [Google Scholar] [CrossRef]
- Ryan, M.H.; Graham, J.H. Little evidence that farmers should consider abundance or diversity of arbuscular mycorrhizal fungi when managing crops. New Phytol. 2018, 220, 1092–1107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albornoz, F.E.; Dixon, K.W.; Lambers, H. Revisiting mycorrhizal dogmas: Are mycorrhizas really functioning as they are widely believed to do? Soil Ecol. Lett. 2021, 3, 73–82. [Google Scholar] [CrossRef]
- Klironomos, J.N. Variation in plant response to native and exotic arbuscular mycorrhizal fungi. Ecology 2003, 84, 2292–2301. [Google Scholar] [CrossRef]
- Pizano, C.; Kitajima, K.; Graham, J.H.; Mangan, S.A. Negative plant-soil feedbacks are stronger in agricultural habitats than in forest fragments in the tropical Andes. Ecology 2019, 100, e02850. [Google Scholar] [CrossRef]
- Hijri, M. Analysis of a large dataset of mycorrhiza inoculation field trials on potato shows highly significant increases in yield. Mycorrhiza 2016, 26, 209–214. [Google Scholar] [CrossRef]
- Pizano, C.; Mangan, S.A.; Graham, J.H.; Kitajima, K. Host-specific effects of soil microbial filtrates prevail over those of arbuscular mycorrhizae in a fragmented landscape. Ecol. Appl. 2017, 27, 1946–1957. [Google Scholar] [CrossRef]
- Bever, J.D.; Richardson, S.C.; Lawrence, B.M.; Holmes, J.; Watson, M. Preferential allocation to beneficial symbiont with spatial structure maintains mycorrhizal mutualism. Ecol. Lett. 2009, 12, 13–21. [Google Scholar] [CrossRef]
- Kiers, E.T.; Duhamel, M.; Beesetty, Y.; Mensah, J.A.; Franken, O.; Verbruggen, E.; Fellbaum, C.R.; Kowalchuk, G.A.; Hart, M.M.; Bago, A.; et al. Reciprocal rewards stabilize cooperation in the mycorrhizal symbiosis. Science 2011, 333, 880–882. [Google Scholar] [CrossRef] [Green Version]
- Ji, B.M.; Bever, J.D. Plant preferential allocation and fungal reward decline with soil phosphorus: Implications for mycorrhizal mutualism. Ecosphere 2016, 7, e01256. [Google Scholar] [CrossRef] [Green Version]
- Garbeva, P.; Veen, J.A.V.; Elsas, J.D.v. MICROBIAL DIVERSITY IN SOIL: Selection of Microbial Populations by Plant and Soil Type and Implications for Disease Suppressiveness. Annu. Rev. Phytopathol. 2004, 42, 243–270. [Google Scholar] [CrossRef] [PubMed]
- Ji, B.; Bentivenga, S.P.; Casper, B.B. Evidence for ecological matching of whole AM fungal communities to the local plant-soil environment. Ecology 2010, 91, 3037–3046. [Google Scholar] [CrossRef] [PubMed]
- Feng, Q.; Ma, H.; Jiang, X.; Wang, X.; Cao, S. What Has Caused Desertification in China? Sci. Rep. 2015, 5, 15998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guoqian, W.; Xuequan, W.; Bo, W.; Qi, L. Desertification and Its Mitigation Strategy in China. J. Resour. Ecol. 2012, 3, 97–104. [Google Scholar] [CrossRef]
- Carrillo-Garcia, Á.; De La Luz, J.L.L.; Bashan, Y.; Bethlenfalvay, G.J. Nurse Plants, Mycorrhizae, and Plant Establishment in a Disturbed Area of the Sonoran Desert. Restor. Ecol. 2002, 7, 321–335. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Xue, Z.; He, X.; Liu, C.; Steinberger, Y. Shifts in composition and diversity of arbuscular mycorrhizal fungi and glomalin contents during revegetation of desertified semiarid grassland. Appl. Soil Ecol. 2017, 115, 60–67. [Google Scholar] [CrossRef]
- Hu, D.; Baskin, J.M.; Baskin, C.C.; Wang, Z.; Zhang, S.; Yang, X.; Huang, Z. Arbuscular mycorrhizal symbiosis and achene mucilage have independent functions in seedling growth of a desert shrub. J. Plant Physiol. 2019, 232, 1–11. [Google Scholar] [CrossRef]
- Chen, Z.; He, X.; Guo, H.; Yao, X.; Chen, C. Diversity of arbuscular mycorrhizal fungi in the rhizosphere of three host plants in the farming–pastoral zone, north China. Symbiosis 2012, 57, 149–160. [Google Scholar] [CrossRef]
- Wu, Y.Q.; Wang, Y.Y.; Lang, Y.F.; Bai, J.H.; Liu, W.J. Dynamics of Arbuscular Mycorrhizal Fungi and Dark Septate Endophytic Fungi under the Canopy of Caragana korshinskii Kom. Bangladesh J. Bot. 2018, 47, 633–641. [Google Scholar]
- Koske, R.E.; Gemma, J.N. Mycorrhizae and succession in plantings of beachgrass in sand dunes. Am. J. Bot. 1997, 84, 118–130. [Google Scholar] [CrossRef]
- Lal, R. Restoring Soil Quality to Mitigate Soil Degradation. Sustainability 2015, 7, 5875–5895. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; He, L.; An, L.; Helgason, T.; Feng, H. Arbuscular mycorrhizal dynamics in a chronosequence of Caragana korshinskii plantations. FEMS Microbiol. Ecol. 2009, 67, 81–92. [Google Scholar] [CrossRef] [Green Version]
- Li, X.R.; Xiao, H.L.; Zhang, J.G.; Wang, X.P. Long-term ecosystem effects of sand-binding vegetation in the Tengger Desert, northern China. Restor. Ecol. 2004, 12, 376–390. [Google Scholar] [CrossRef]
- Ma, F.; Na, X.; Xu, T. Drought responses of three closely related Caragana species: Implication for their vicarious distribution. Ecol. Evol. 2016, 6, 2763–2773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koide, R.T.; Li, M. Appropriate controls for vesicular-arbuscular mycorrhiza research. New Phytol. 1989, 111, 35–44. [Google Scholar] [CrossRef]
- Ke, P.J.; Zee, P.C.; Fukami, T. Dynamic plant-soil microbe interactions: The neglected effect of soil conditioning time. New Phytol. 2021, 231, 1546–1558. [Google Scholar] [CrossRef] [PubMed]
- Mc, G.T.; Miller, M.H.; Evans, D.G.; Fairchild, G.L.; Swan, J.A. A new method which gives an objective measure of colonization of roots by vesicular-arbuscular mycorrhizal fungi. New Phytol. 1990, 115, 495–501. [Google Scholar] [CrossRef]
- Murphy, J.; Riley, J.P. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Bremner, J.M.; Tabatabai, M.A. Use of an ammonia electrode for determination of ammonium in Kjeldahl analysis of soils. Commun. Soil Sci. Plant Anal. 2008, 3, 159–165. [Google Scholar] [CrossRef]
- Allen, G.C.; Flores-Vergara, M.A.; Krasynanski, S.; Kumar, S.; Thompson, W.F. A modified protocol for rapid DNA isolation from plant tissues using cetyltrimethylammonium bromide. Nat Protoc. 2006, 1, 2320–2325. [Google Scholar] [CrossRef]
- Muneer, M.A.; Tarin, M.W.K.; Chen, X.; Afridi, M.S.; Iqbal, A.; Munir, M.Z.; Zheng, C.; Zhang, J.; Ji, B. Differential response of mycorrhizal fungi linked with two dominant plant species of temperate grassland under varying levels of N-addition. Appl. Soil Ecol. 2022, 170, 104272. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [Green Version]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Package ‘vegan’. Community Ecol. Package Version 2013, 2, 1–295. [Google Scholar]
- De Caceres, M.; Legendre, P. Associations between species and groups of sites: Indices and statistical inference. Ecology 2009, 90, 3566–3574. [Google Scholar] [CrossRef]
- Shi, Z.Y.; Wang, Y.M.; Xu, S.X.; Lan, Z.J.; Mickan, B.S.; Zhang, X.L.; Wang, F.Y. Arbuscular Mycorrhizal Fungi Enhance Plant Diversity, Density and Productivity of Spring Ephemeral Community in Desert Ecosystem. Not. Bot. Horti Agrobot. Cluj-Napoca 2017, 45, 301–307. [Google Scholar] [CrossRef] [Green Version]
- Hoeksema, J.D.; Chaudhary, V.B.; Gehring, C.A.; Johnson, N.C.; Karst, J.; Koide, R.T.; Pringle, A.; Zabinski, C.; Bever, J.D.; Moore, J.C.; et al. A meta-analysis of context-dependency in plant response to inoculation with mycorrhizal fungi. Ecol. Lett. 2010, 13, 394–407. [Google Scholar] [CrossRef]
- Ma, C.C.; Gao, Y.B.; Liu, H.F.; Wang, J.L.; Guo, H.Y. Interspecific transition among Caragana microphylla, C. davazamcii and C. korshinskii along geographic gradient. I. Ecological and RAPD evidence. Acta Bot. Sin. 2003, 45, 1218–1227. [Google Scholar]
- Xie, L.; Ma, C.; Guo, H.; Li, Q.; Gao, Y. Distribution pattern of Caragana species under the influence of climate gradient in the Inner Mongolia region, China. J. Arid. Land 2013, 6, 311–323. [Google Scholar] [CrossRef] [Green Version]
- Ma, C.C.; Gao, Y.B.; Guo, H.Y.; Wang, J.L. Photosynthesis, transpiration, and water use efficiency of Caragana microphylla, C. intermedia, and C. korshinskii. Photosynthetica 2004, 42, 65–70. [Google Scholar] [CrossRef]
- Ma, C.-C.; Guo, H.-Y.; Wu, J.-B.; Wang, J.-L.; Qi, S.-X.; Wei, Y.-R.; Gao, Y.-B. Acclimation of Photosynthetic Traits of Caragana Species to Desert Environment in Inner Mongolian Plateau. Arid. Land Res. Manag. 2013, 28, 87–101. [Google Scholar] [CrossRef]
- van der Heijden, M.G.A.; Boller, T.; Wiemken, A.; Sanders, I.R. Different Arbuscular Mycorrhizal Fungal Species Are Potential Determinants of Plant Community Structure. Ecology 1998, 79, 2082–2091. [Google Scholar] [CrossRef]
- Frew, A.; Wilson, B.A.L. Different mycorrhizal fungal communities differentially affect plant phenolic-based resistance to insect herbivory. Rhizosphere 2021, 19, 100365. [Google Scholar] [CrossRef]
- Bi, Y.L.; Wang, K.; Wang, J. Effect of different inoculation treatments on AM fungal communities and the sustainability of soil remediation in Daliuta coal mining subsidence area in northwest China. Appl. Soil Ecol. 2018, 132, 107–113. [Google Scholar] [CrossRef]
- Pankova, H.; Lepinay, C.; Rydlova, J.; Voriskova, A.; Janouskova, M.; Dostalek, T.; Munzbergova, Z. Arbuscular mycorrhizal fungi and associated microbial communities from dry grassland do not improve plant growth on abandoned field soil. Oecologia 2018, 186, 677–689. [Google Scholar] [CrossRef]
- Qin, Z.F.; Zhang, H.Y.; Feng, G.; Christie, P.; Zhang, J.L.; Li, X.L.; Gai, J.P. Soil phosphorus availability modifies the relationship between AM fungal diversity and mycorrhizal benefits to maize in an agricultural soil. Soil Biol. Biochem. 2020, 144, 107790. [Google Scholar] [CrossRef]
- Ruiz-Lozano, J.M.; Azcón, R. Symbiotic efficiency and infectivity of an autochthonous arbuscular mycorrhizal Glomus sp. from saline soils and Glomus deserticola under salinity. Mycorrhiza 2000, 10, 137–143. [Google Scholar] [CrossRef]
- Middleton, E.L.; Richardson, S.; Koziol, L.; Palmer, C.E.; Yermakov, Z.; Henning, J.A.; Schultz, P.A.; Bever, J.D. Locally adapted arbuscular mycorrhizal fungi improve vigor and resistance to herbivory of native prairie plant species. Ecosphere 2015, 6, 276. [Google Scholar] [CrossRef]
- Harris-Valle, C.; Esqueda, M.; Gutierrez, A.; Castellanos, A.E.; Gardea, A.A.; Berbara, R. Physiological response of Cucurbita pepo var. pepo mycorrhized by Sonoran desert native arbuscular fungi to drought and salinity stresses. Braz. J. Microbiol. 2018, 49, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Bauer, J.T.; Koziol, L.; Bever, J.D. Local adaptation of mycorrhizae communities changes plant community composition and increases aboveground productivity. Oecologia 2020, 192, 735–744. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.-S.; Zou, Y.-N.; Huang, Y.-M. The arbuscular mycorrhizal fungus Diversispora spurca ameliorates effects of waterlogging on growth, root system architecture and antioxidant enzyme activities of citrus seedlings. Fungal Ecol. 2013, 6, 37–43. [Google Scholar] [CrossRef]
- Maherali, H.; Klironomos, J.N. Influence of phylogeny on fungal community assembly and ecosystem functioning. Science 2007, 316, 1746–1748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Y.; Feng, J.; Han, M.; Zhu, B. Responses of arbuscular mycorrhizal fungi to nitrogen addition: A meta-analysis. Glob. Chang. Biol. 2020, 26, 7229–7241. [Google Scholar] [CrossRef]
- Weber, S.E.; Diez, J.M.; Andrews, L.V.; Goulden, M.L.; Aronson, E.L.; Allen, M.F. Responses of arbuscular mycorrhizal fungi to multiple coinciding global change drivers. Fungal Ecol. 2019, 40, 62–71. [Google Scholar] [CrossRef]
- Yang, W.; Guo, X.J.; Wang, Q. Contribution of different arbuscular mycorrhizal fungal inoculum to Elymus nutans under nitrogen addition. Ann. Microbiol. 2018, 68, 689–696. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Height | Canopy | Photosynthetic Rate | Transpiration Rate | Vapor Pressure Deficit | Stomatal Conductance | Colonization Rate | Shoot Biomass | Foliar N | Foliar P | Foliar N:P | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Host | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.2394 | <0.001 | <0.001 | 0.003 | 0.011 |
| Inoculation | <0.001 | 0.001 | 0.001 | 0.189 | 0.000 | 0.002 | 0.0241 | <0.001 | 0.102 | 0.163 | 0.076 |
| Year | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | |||||
| Host:Inoculation | <0.001 | 0.001 | 0.029 | 0.026 | 0.501 | 0.015 | 0.0646 | 0.0119 | 0.0406 | 0.0130 | 0.5085 |
| Host:Year | <0.001 | 0.012 | <0.001 | <0.001 | <0.001 | 0.002 | |||||
| Inoculation:Year | 0.026 | 0.001 | 0.584 | 0.435 | 0.307 | 0.213 | |||||
| Host:Inoculation:Year | 0.769 | 0.341 | 0.035 | 0.151 | 0.033 | 0.109 |
| C. korshinskii | C. microphylla | |||||||
|---|---|---|---|---|---|---|---|---|
| Inoculated vs. Un-Inoculated | Home vs. Away | Inoculated vs. Un-Inoculated | Home vs. Away | |||||
| Shoot biomass | + | 0.144 | + | <0.001 | + | 0.977 | − | 0.38 |
| Photosynthetic rate (2017) | + | 0.132 | + | <0.001 | + | 0.376 | + | <0.001 |
| Photosynthetic rate (2018) | + | 0.017 | + | 0.102 | − | 0.426 | − | 0.144 |
| Foliar N | + | 0.854 | + | 0.002 | − | 0.002 | + | 0.089 |
| Foliar P | + | 0.244 | + | 0.009 | − | 0.008 | − | 0.444 |
| Foliar N:P | − | 0.397 | − | 0.365 | + | 0.383 | + | 0.003 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, X.; Wang, Z.; Zhang, J.; Wang, P.; Li, Y.; Ji, B. Host-Specific Effects of Arbuscular Mycorrhizal Fungi on Two Caragana Species in Desert Grassland. J. Fungi 2021, 7, 1077. https://doi.org/10.3390/jof7121077
Guo X, Wang Z, Zhang J, Wang P, Li Y, Ji B. Host-Specific Effects of Arbuscular Mycorrhizal Fungi on Two Caragana Species in Desert Grassland. Journal of Fungi. 2021; 7(12):1077. https://doi.org/10.3390/jof7121077
Chicago/Turabian StyleGuo, Xin, Zhen Wang, Jing Zhang, Ping Wang, Yaoming Li, and Baoming Ji. 2021. "Host-Specific Effects of Arbuscular Mycorrhizal Fungi on Two Caragana Species in Desert Grassland" Journal of Fungi 7, no. 12: 1077. https://doi.org/10.3390/jof7121077
APA StyleGuo, X., Wang, Z., Zhang, J., Wang, P., Li, Y., & Ji, B. (2021). Host-Specific Effects of Arbuscular Mycorrhizal Fungi on Two Caragana Species in Desert Grassland. Journal of Fungi, 7(12), 1077. https://doi.org/10.3390/jof7121077