
One Century of Study: What We Learned about Paracoccidioides and How This Pathogen Contributed to Advances in Antifungal Therapy

 , ,
, , 
Abstract
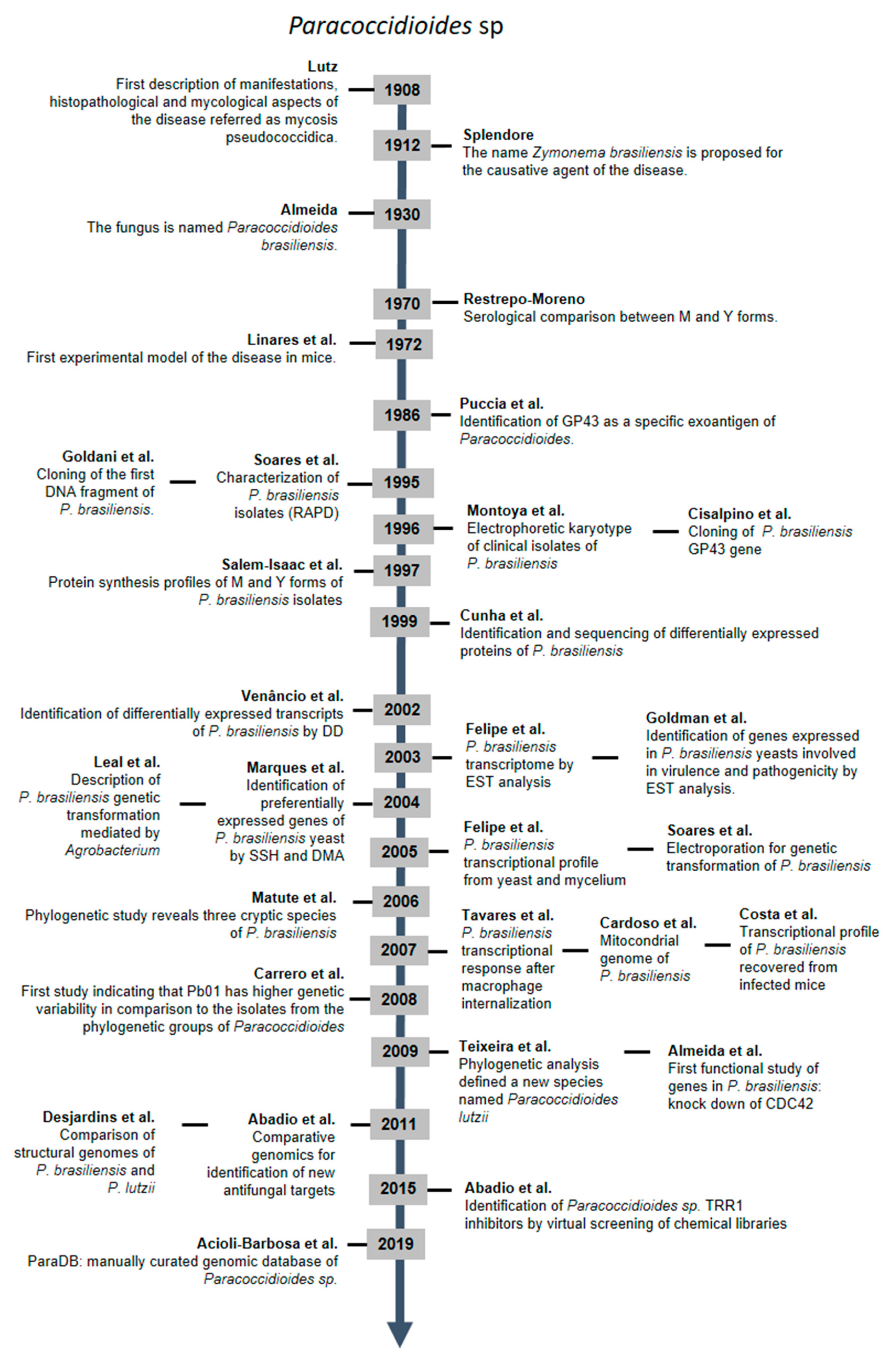
:1. From the First Description of the Disease to Functional Genomics—112 Years of Gathered Knowledge. A Brazilian Middle West Perspective
2. Search for New Therapeutic Options—A Journey
2.1. A Starting Point—Available Therapeutic Options
2.2. Comparative Genomics Searching for Drug Targets
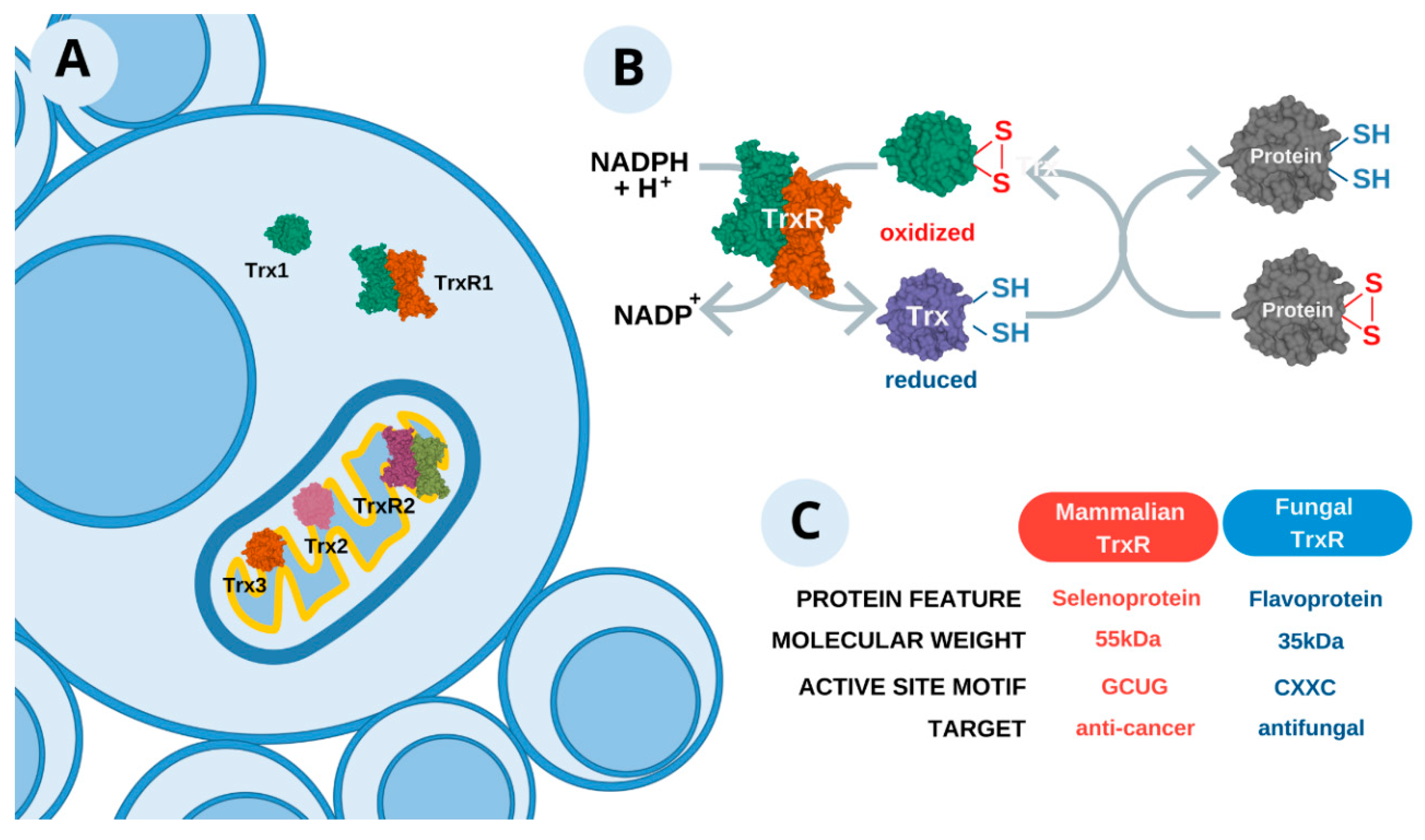
2.3. Thioredoxin System—A Promising Approach
2.3.1. General Definition
2.3.2. Fungus Selectivity
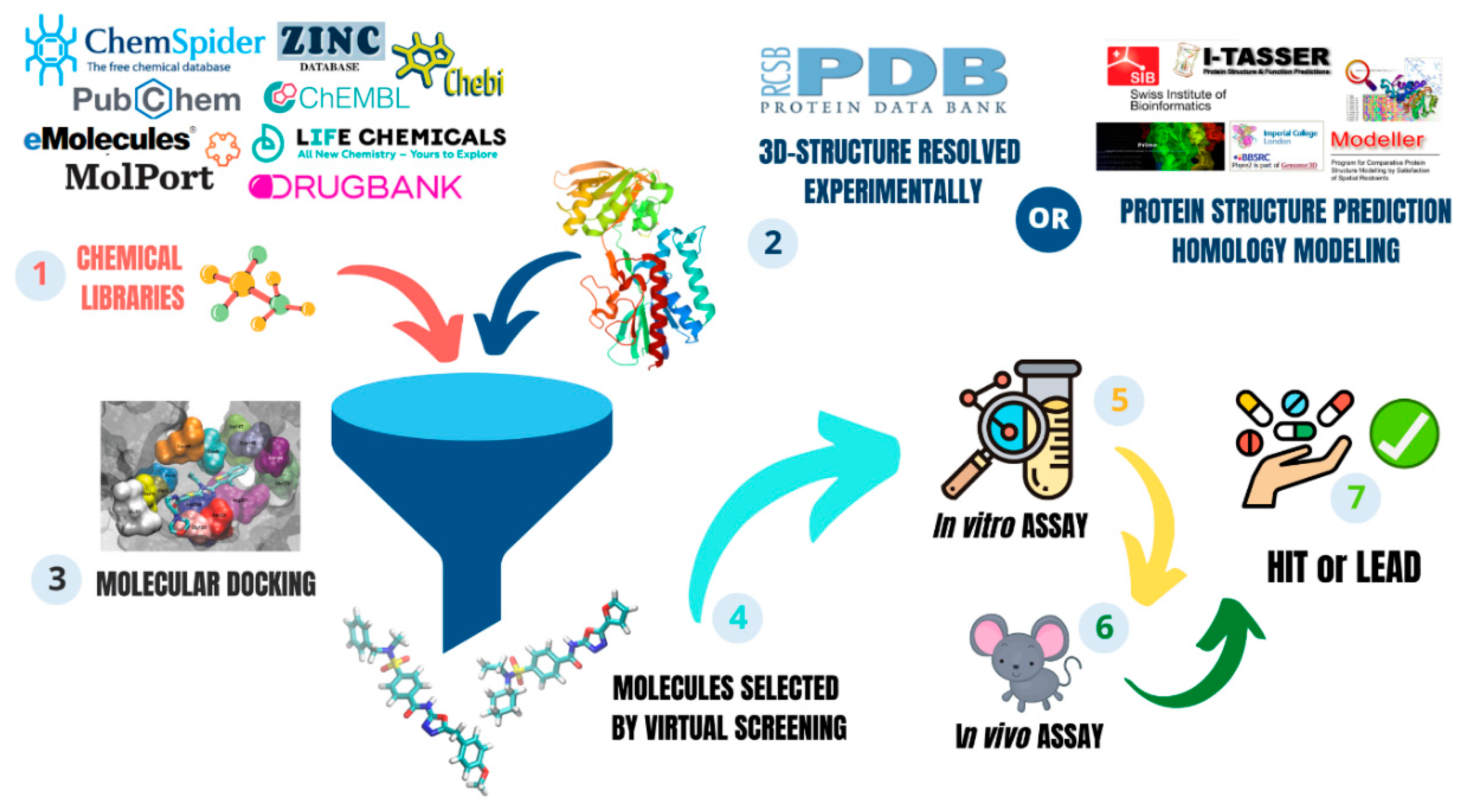
2.4. In Silico Strategy for New Antifungal Development
2.4.1. Virtual Screening
2.4.2. Drug Repositioning
2.5. Drug Targets Discovered from Natural Products
3. Nanotechnology as an Approach for Alternative Therapies
4. Challenges and Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lutz, A. A pseudococcidic mycosis localized in the mouth and observed in Brazil. A contribution to the knowledge of the American hyphoblastomycoses (in Portuguese). Brazil-Medico 1908, 22, 121–124. [Google Scholar]
- Lacaz, C.S. Evolução dos Conhecimentos Sobre a Paracoccidioidomicose. Um Pouco de Sua História. In Paracoccidoidomicose-Blastomicose Sul-Americana; Del Negro, G., Lacaz, C.S., Fiorillo, A.M., Eds.; Sarvier-Eduso: São Paulo, Brazil, 1982. [Google Scholar]
- Almeida, F.P. Estudos comparativos do granuloma coccidiótico nos Estados Unidos e no Brasil: Novo gênero para o parasito brasileiro. An. Fac. Med. Sao Paulo 1930, 5, 125–141. [Google Scholar]
- Bocca, A.L.; Amaral, A.C.; Teixeira, M.M.; Sato, P.K.; Shikanai-Yasuda, M.A.; Soares Felipe, M.S. Paracoccidioidomycosis: Eco-epidemiology, taxonomy and clinical and therapeutic issues. Future Microbiol. 2013, 8, 1177–1191. [Google Scholar] [CrossRef] [PubMed]
- Shikanai-Yasuda, M.A.; Mendes, R.P.; Colombo, A.L.; Queiroz-Telles, F.; Kono, A.S.G.; Paniago, A.M.M.; Nathan, A.; Valle, A.C.F.D.; Bagagli, E.; Benard, G.; et al. Brazilian guidelines for the clinical management of paracoccidioidomycosis. Rev. Soc. Bras. Med. Trop. 2017, 50, 715–740, Erratum in Rev. Soc. Bras. Med. Trop. 2017, 50, 879–880. [Google Scholar] [CrossRef]
- Barbosa, W.; Daher, R.; Oliveira, A.R. Lymphatic abdominal forms of South American blastomycosis. Rev. Inst. Med. Trop. Sao Paulo 1968, 10, 12. [Google Scholar]
- Andrade, A.L.S.S.D. Paracoccidioidomicose linfático-abdominal-Contribuição ao seu estudo. Rev. Patol. Trop. 1983, 12, 165–256. [Google Scholar]
- Martinez, R. New Trends in Paracoccidioidomycosis Epidemiology. J. Fungi 2017, 3, 1. [Google Scholar] [CrossRef] [Green Version]
- Vieira, G.D.D.; Alves, T.D.C.; Lima, S.M.D.D.; Camargo, L.M.A.; Sousa, C.M.D. Paracoccidioidomycosis in a western Brazilian Amazon State: Clinical-epidemiologic profile and spatial distribution of the disease. Rev. Soc. Bras. Med. Trop. 2014, 47, 63–68. [Google Scholar] [CrossRef] [Green Version]
- Del Negro, G.M.; Benard, G.; de Assis, C.M.; Vidal, M.S.; Garcia, N.M.; Otani, C.; Shikanai-Yasuda, M.A.; Lacaz, C. Lack of reactivity of paracoccidioidomycosis sera in the double immunodiffusion test with the gp43 antigen: Report of two cases. J. Med. Vet. Mycol. 1995, 33, 113–116. [Google Scholar] [CrossRef]
- Vidal, M.S.; Benard, G.; Brito, T.; Dantas, K.C.; Pereira, C.N.; França, F.O.; da Silva, A.M.; Martins, J.E. Atypical serological response marked by a lack of detectable anti-gp43 antibodies in a patient with disseminated paracoccidioidomycosis. J. Clin. Microbiol. 2005, 43, 3014–3016. [Google Scholar] [CrossRef] [Green Version]
- Batista, J., Jr.; de Camargo, Z.P.; Fernandes, G.F.; Vicentini, A.P.; Fontes, C.J.; Hahn, R.C. Is the geographical origin of a Paracoccidioides brasiliensis isolate important for antigen production for regional diagnosis of paracoccidioidomycosis? Mycoses 2010, 53, 176–180. [Google Scholar] [CrossRef] [PubMed]
- Molinari-Madlum, E.E.W.I.; Felipe, M.S.S.; Soares, C.M.A. Virulence of Paracoccidioides brasiliensis isolates can be correlated to groups defined by random amplified polymorphic DNA analysis. Med. Mycol. 1999, 37, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Hahn, R.C.; Macedo, A.M.; Santos, N.L.; Resende, J.C.; Hamdan, J.S. Characterization of Paracoccidioides brasiliensis atypical isolates by random amplified polymorphic DNA analysis. Rev. Iberoam. Micol. 2002, 19, 49–51. [Google Scholar] [PubMed]
- Hahn, R.C.; Macedo, A.M.; Fontes, C.J.; Batista, R.D.; Santos, N.L.; Hamdan, J.S. Randomly amplified polymorphic DNA as a valuable tool for epidemiological studies of Paracoccidioides brasiliensis. J. Clin. Microbiol. 2003, 41, 2849–2854. [Google Scholar] [CrossRef] [Green Version]
- Taylor, J.W.; Jacobson, D.J.; Kroken, S.; Kasuga, T.; Geiser, D.M.; Hibbett, D.S.; Fisher, M.C. Phylogenetic species recognition and species concepts in fungi. Fungal Genet. Biol. 2000, 31, 21–32. [Google Scholar] [CrossRef] [Green Version]
- Matute, D.R.; Sepúlveda, V.E. Fungal species boundaries in the genomics era. Fungal Genet. Biol. 2019, 131, 103249. [Google Scholar] [CrossRef]
- Matute, D.R.; McEwen, J.G.; Puccia, R.; Montes, B.A.; San-Blas, G.; Bagagli, E.; Rauscher, J.T.; Restrepo, A.; Morais, F.; Niño-Vega, G.; et al. Cryptic speciation and recombination in the fungus Paracoccidioides brasiliensis as revealed by gene genealogies. Mol. Biol. Evol. 2006, 23, 65–73. [Google Scholar] [CrossRef]
- Carrero, L.L.; Niño-Vega, G.; Teixeira, M.M.; Carvalho, M.J.; Soares, C.M.; Pereira, M.; Jesuino, R.S.; McEwen, J.G.; Mendoza, L.; Taylor, J.W.; et al. New Paracoccidioides brasiliensis isolate reveals unexpected genomic variability in this human pathogen. Fungal Genet. Biol. 2008, 45, 605–612. [Google Scholar] [CrossRef]
- Da Silva, S.P.; Borges-Walmsley, M.I.; Pereira, I.S.; Soares, C.M.; Walmsley, A.R.; Felipe, M.S. Differential expression of an hsp70 gene during transition from the mycelial to the infective yeast form of the human pathogenic fungus Paracoccidioides brasiliensis. Mol. Microbiol. 1999, 31, 1039–1050. [Google Scholar] [CrossRef]
- Teixeira, M.M.; Theodoro, R.C.; de Carvalho, M.J.; Fernandes, L.; Paes, H.C.; Hahn, R.C.; Mendoza, L.; Bagagli, E.; San-Blas, G.; Felipe, M.S. Phylogenetic analysis reveals a high level of speciation in the Paracoccidioides genus. Mol. Phylogenet. Evol. 2009, 52, 273–283. [Google Scholar] [CrossRef]
- Teixeira, M.M.; Theodoro, R.C.; Oliveira, F.F.; Machado, G.C.; Hahn, R.C.; Bagagli, E.; San-Blas, G.; Felipe, M.S. Paracoccidioides lutzii sp. nov.: Biological and clinical implications. Med. Mycol. 2014, 52, 19–28. [Google Scholar] [CrossRef] [Green Version]
- Turissini, D.A.; Gomez, O.M.; Teixeira, M.M.; McEwen, J.G.; Matute, D.R. Species boundaries in the human pathogen Paracoccidioides. Fung. Genet. Biol. 2017, 106, 9–25. [Google Scholar] [CrossRef] [PubMed]
- Desjardins, C.A.; Champion, M.D.; Holder, J.W.; Muszewska, A.; Goldberg, J.; Bailão, A.M.; Brigido, M.M.; Ferreira, M.E.; Garcia, A.M.; Grynberg, M.; et al. Comparative genomic analysis of human fungal pathogens causing paracoccidioidomycosis. PLoS Genet. 2011, 7, e1002345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teixeira, M.M.; Cattana, M.E.; Matute, D.R.; Muñoz, J.F.; Arechavala, A.; Isbell, K.; Schipper, R.; Santiso, G.; Tracogna, F.; de los Ángeles Sosa, M.; et al. Genomic diversity of the human pathogen Paracoccidioides across the South American continent. Fung. Genet. Biol. 2020, 140, 103395. [Google Scholar] [CrossRef]
- Mavengere, H.; Mattox, K.; Teixeira, M.M.; Sepulveda, V.; Gomez, O.; Hernandez, O.; McEwen, J.; Matute, D.R. Paracoccidioides genomes reflect high levels of species divergence and little interspecific gene flow. mBio 2020, 11, e01999-20. [Google Scholar] [CrossRef]
- Muñoz, J.F.; Gallo, J.E.; Misas, E.; Priest, M.; Imamovic, A.; Young, S.; Zeng, Q.; Clay, O.K.; McEwen, J.G.; Cuomo, C.A. Genome Update of the Dimorphic Human Pathogenic Fungi Causing Paracoccidioidomycosis. PLoS Negl. Trop. Dis. 2014, 8, e3348. [Google Scholar] [CrossRef]
- Aciole Barbosa, D.; Menegidio, F.B.; Alencar, V.C.; Gonçalves, R.S.; Silva, J.F.S.; Vilas Boas, R.O.; Faustino de Maria, Y.N.L.; Jabes, D.L.; Costa de Oliveira, R.; Nunes, L.R. ParaDB: A manually curated database containing genomic annotation for the human pathogenic fungi Paracoccidioides spp. PLoS Negl. Trop. Dis. 2019, 13, e0007576. [Google Scholar] [CrossRef] [Green Version]
- Goldani, L.Z.; Maia, A.L.; Sugar, A.M. Cloning and nucleotide sequence of a specific DNA fragment from Paracoccidioides brasiliensis. J. Clin. Microbiol. 1995, 33, 1652–1654. [Google Scholar] [CrossRef] [Green Version]
- Cisalpino, P.S.; Puccia, R.; Yamauchi, L.M.; Cano, M.I.; da Silveira, J.F.; Travassos, L.R. Cloning, characterization, and epitope expression of the major diagnostic antigen of Paracoccidioides brasiliensis. J. Biol. Chem. 1996, 23, 4553–4560. [Google Scholar] [CrossRef] [Green Version]
- Yarzabal, L.A.; Bout, D.; Naquira, F.; Fruit, J.; Andrieu, S. Identification and purification of the specific antigen of Paracoccidioides brasiliensis responsible for immunoelectrophoretic band E. Sabouraudia 1977, 15, 79–85. [Google Scholar] [CrossRef]
- Salem-Izacc, S.M.; Jesuino, R.S.; Brito, W.A.; Pereira, M.; Felipe, M.S.; Soares, C.M. Protein synthesis patterns of Paracoccidiodes brasiliensis isolates in stage-specific forms and during cellular differentiation. J. Med. Vet. Mycol. 1997, 35, 205–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunha, A.F.; Sousa, M.V.; Silva, S.P.; Jesuíno, R.S.; Soares, C.M.; Felipe, M.S. Identification, N-terminal region sequencing and similarity analysis of differentially expressed proteins in Paracoccidioides brasiliensis. Med. Mycol. 1999, 37, 115–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daher, B.S.; Venancio, E.J.; de Freitas, S.M.; Báo, S.N.; Vianney, P.V.; Andrade, R.V.; Dantas, A.S.; Soares, C.M.; Silva-Pereira, I.; Felipe, M.S. The highly expressed yeast gene pby20 from Paracoccidioides brasiliensis encodes a flavodoxin-like protein. Fungal Genet. Biol. 2005, 42, 434–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venancio, E.J.; Kyaw, C.M.; Mello, C.V.; Silva, S.P.; Soares, C.M.; Felipe, M.S.; Silva-Pereira, I. Identification of differentially expressed transcripts in the human pathogenic fungus Paracoccidioides brasiliensis by differential display. Med. Mycol. 2002, 40, 45–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felipe, M.S.; Andrade, R.V.; Petrofeza, S.S.; Maranhão, A.Q.; Torres, F.A.; Albuquerque, P.; Arraes, F.B.; Arruda, M.; Azevedo, M.O.; Baptista, A.J.; et al. Transcriptome characterization of the dimorphic and pathogenic fungus Paracoccidioides brasiliensis by EST analysis. Yeast 2003, 20, 263–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felipe, M.S.; Andrade, R.V.; Arraes, F.B.; Nicola, A.M.; Maranhão, A.Q.; Torres, F.A.; Silva-Pereira, I.; Poças-Fonseca, M.J.; Campos, E.G.; Moraes, L.M.; et al. PbGenome Network. Transcriptional profiles of the human pathogenic fungus Paracoccidioides brasiliensis in mycelium and yeast cells. J. Biol. Chem. 2005, 280, 24706–24714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felipe, M.S.; Torres, F.A.; Maranhão, A.Q.; Silva-Pereira, I.; Poças-Fonseca, M.J.; Campos, E.G.; Moraes, L.M.; Arraes, F.B.; Carvalho, M.J.; Andrade, R.V.; et al. Functional genome of the human pathogenic fungus Paracoccidioides brasiliensis. FEMS Immunol. Med. Microbiol. 2005, 45, 369–381. [Google Scholar] [CrossRef] [Green Version]
- Andrade, R.V.; Paes, H.C.; Nicola, A.M.; de Carvalho, M.J.; Fachin, A.L.; Cardoso, R.S.; Silva, S.S.; Fernandes, L.; Silva, S.P.; Donadi, E.A.; et al. Cell organisation, sulphur metabolism and ion transport-related genes are differentially expressed in Paracoccidioides brasiliensis mycelium and yeast cells. BMC Genom. 2006, 7, 208. [Google Scholar] [CrossRef] [Green Version]
- Goldman, G.H.; dos Reis Marques, E.; Duarte Ribeiro, D.C.; de Souza Bernardes, L.A.; Quiapin, A.C.; Vitorelli, P.M.; Savoldi, M.; Semighini, C.P.; de Oliveira, R.C.; Nunes, L.R.; et al. Expressed sequence tag analysis of the human pathogen Paracoccidioides brasiliensis yeast phase: Identification of putative homologues of Candida albicans virulence and pathogenicity genes. Eukaryot. Cell 2003, 2, 34–48. [Google Scholar] [CrossRef] [Green Version]
- Marques, E.R.; Ferreira, M.E.; Drummond, R.D.; Felix, J.M.; Menossi, M.; Savoldi, M.; Travassos, L.R.; Puccia, R.; Batista, W.L.; Carvalho, K.C.; et al. Identification of genes preferentially expressed in the pathogenic yeast phase of Paracoccidioides brasiliensis, using suppression subtraction hybridization and differential macroarray analysis. Mol. Genet. Genom. 2004, 271, 667–677. [Google Scholar] [CrossRef]
- Nunes, L.R.; Costa de Oliveira, R.; Leite, D.B.; da Silva, V.S.; dos Reis Marques, E.; da Silva Ferreira, M.E.; Ribeiro, D.C.; de Souza Bernardes, L.A.; Goldman, M.H.; Puccia, R.; et al. Transcriptome analysis of Paracoccidioides brasiliensis cells undergoing mycelium-to-yeast transition. Eukaryot. Cell. 2005, 4, 2115–2128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bastos, K.P.; Bailão, A.M.; Borges, C.L.; Faria, F.P.; Felipe, M.S.; Silva, M.G.; Martins, W.S.; Fiúza, R.B.; Pereira, M.; Soares, C.M. The transcriptome analysis of early morphogenesis in Paracoccidioides brasiliensis mycelium reveals novel and induced genes potentially associated to the dimorphic process. BMC Microbiol. 2007, 7, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tavares, A.H.; Silva, S.S.; Dantas, A.; Campos, E.G.; Andrade, R.V.; Maranhão, A.Q.; Brígido, M.M.; Passos-Silva, D.G.; Fachin, A.L.; Teixeira, S.M.; et al. Early transcriptional response of Paracoccidioides brasiliensis upon internalization by murine macrophages. Microbes Infect. 2007, 9, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Bailão, A.M.; Schrank, A.; Borges, C.L.; Dutra, V.; Molinari-Madlum, E.E.W.I.; Felipe, M.S.S.; Mendes-Giannini, S.M.J.; Martins, W.S.; Pereira, M.; Almeida Soares, C.M. Differential gene expression by Paracoccidioides brasiliensis in host interaction conditions: Representational difference analysis identifies candidate genes associated with fungal pathogenesis. Microbes Infect. 2006, 8, 2686–2697. [Google Scholar] [CrossRef]
- Bailão, A.M.; Shrank, A.; Borges, C.L.; Parente, J.A.; Dutra, V.; Felipe, M.S.; Fiúza, R.B.; Pereira, M.; de Almeida Soares, C.M. The transcriptional profile of Paracoccidioides brasiliensis yeast cells is influenced by human plasma. FEMS Immunol. Med. Microbiol. 2007, 51, 43–57. [Google Scholar] [CrossRef] [Green Version]
- Costa, M.; Borges, C.L.; Bailão, A.M.; Meirelles, G.V.; Mendonça, Y.A.; Dantas, S.F.I.M.; de Faria, F.P.; Felipe, M.S.S.; Molinari-Madlum, E.E.W.I.; Mendes-Giannini, M.J.; et al. Transcriptome profiling of Paracoccidioides brasiliensis yeast-phase cells recovered from infected mice brings new insights into fungal response upon host interaction. Microbiology 2007, 153, 4194–4207. [Google Scholar] [CrossRef] [Green Version]
- Pereira, M.; Bailão, A.M.; Parente, J.A.; Borges, C.L.; Salem-Izacc, S.M.; Soares, C.M. Preferential transcription of Paracoccidioides brasiliensis genes: Host niche and time-dependent expression. Mem. Inst. Oswaldo Cruz. 2009, 104, 486–491. [Google Scholar] [CrossRef] [Green Version]
- Leal, C.V.; Montes, B.A.; Mesa, A.C.; Rua, A.L.; Corredor, M.; Restrepo, A.; McEwen, J.G. Agrobacterium tumefaciens-mediated transformation of Paracoccidioides brasiliensis. Med. Mycol. 2004, 42, 391–395. [Google Scholar] [CrossRef] [Green Version]
- Soares, R.B.; Velho, T.A.; De Moraes, L.M.; Azevedo, M.O.; Soares, C.M.; Felipe, M.S. Hygromycin B-resistance phenotype acquired in Paracoccidioides brasiliensis via plasmid DNA integration. Med. Mycol. 2005, 43, 719–723. [Google Scholar] [CrossRef] [Green Version]
- Almeida, A.J.; Cunha, C.; Carmona, J.A.; Sampaio-Marques, B.; Carvalho, A.; Malavazi, I.; Steensma, H.Y.; Johnson, D.I.; Leão, C.; Logarinho, E.; et al. Cdc42p controls yeast-cell shape and virulence of Paracoccidioides brasiliensis. Fungal Genet. Biol. 2009, 46, 919–926. [Google Scholar] [CrossRef]
- Ruiz, O.H.; Gonzalez, A.; Almeida, A.J.; Tamayo, D.; Garcia, A.M.; Restrepo, A.; McEwen, J.G. Alternative oxidase mediates pathogen resistance in Paracoccidioides brasiliensis infection. PLoS Negl. Trop. Dis. 2011, 5, e1353. [Google Scholar] [CrossRef]
- Goes, T.; Bailão, E.F.; Correa, C.R.; Bozzi, A.; Santos, L.I.; Gomes, D.A.; Soares, C.M.; Goes, A.M. New developments of RNAi in Paracoccidioides brasiliensis: Prospects for high-throughput, genome-wide, functional genomics. PLoS Negl. Trop. Dis. 2014, 8, e3173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, I.; Hernandez, O.; Tamayo, D.; Muñoz, J.F.; García, A.M.; Gómez, B.L.; Restrepo, A.; McEwen, J.G. Paracoccidioides brasiliensis PbP27 gene: Knockdown procedures and functional characterization. FEMS Yeast Res. 2014, 14, 270–280. [Google Scholar] [CrossRef] [Green Version]
- Tamayo, D.; Muñoz, J.F.; Lopez, Á.; Urán, M.; Herrera, J.; Borges, C.L.; Restrepo, Á.; Soares, C.M.; Taborda, C.P.; Almeida, A.J.; et al. Identification and Analysis of the Role of Superoxide Dismutases Isoforms in the Pathogenesis of Paracoccidioides spp. PLoS Negl. Trop. Dis. 2016, 10, e0004481. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, F.F.; Oliveira, A.F.; Landgraf, T.N.; Cunha, C.; Carvalho, A.; Vendruscolo, P.E.; Gonçales, R.A.; Almeida, F.; da Silva, T.A.; Rodrigues, F.; et al. Impact of Paracoccin Gene Silencing on Paracoccidioides brasiliensis Virulence. mBio 2017, 8, e00537-17. [Google Scholar] [CrossRef] [Green Version]
- Tamayo, D.; Muñoz, J.F.; Almeida, A.J.; Puerta, J.D.; Restrepo, Á.; Cuomo, C.A.; McEwen, J.G.; Hernández, O. Paracoccidioides spp. catalases and their role in antioxidant defense against host defense responses. Fungal Genet. Biol. 2017, 100, 22–32. [Google Scholar] [CrossRef] [Green Version]
- Marcos, C.M.; de Oliveira, H.C.; Assato, P.A.; de Andrade, C.R.; Fusco-Almeida, A.M.; Mendes-Giannini, M.J.S. Paracoccidioides brasiliensis 14-3-3 protein is important for virulence in a murine model. Med. Mycol. 2019, 57, 900–904. [Google Scholar] [CrossRef] [PubMed]
- Marcos, C.M.; Tamer, G.; de Oliveira, H.C.; Assato, P.A.; Scorzoni, L.; Santos, C.T.; de Lacorte Singulani, J.; de Fátima da Silva, J.; de Almeida, R.; de Paula, E.; et al. Down-regulation of TUFM impairs host cell interaction and virulence by Paracoccidioides brasiliensis. Sci. Rep. 2019, 9, 17206. [Google Scholar] [CrossRef]
- Oliveira, L.N.; Gonçales, R.A.; Silva, M.G.; Lima, R.M.; Tomazett, M.V.; de Curcio, J.S.; Paccez, J.D.; Cruz-Leite, V.R.M.; Rodrigues, F.; Lima, P.S.; et al. Characterization of a heme-protein responsive to hypoxia in Paracoccidioides brasiliensis. Fungal Genet. Biol. 2020, 144, 103446. [Google Scholar] [CrossRef]
- Nett, J.E.; Andes, D.R. Antifungal Agents: Spectrum of Activity, Pharmacology, and Clinical Indications. Infect. Dis. Clin. N. Am. 2016, 30, 51–83. [Google Scholar] [CrossRef]
- Cohen, B.E. Amphotericin B Membrane Action: Role for Two Types of Ion Channels in Eliciting Cell Survival and Lethal Effects. J. Membr. Biol. 2010, 238, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Baginski, M.; Resat, H.; Mc Cammon, J.A. Molecular Properties of Amphotericin B Membrane Channel: A Molecular Dynamics Simulation. Mol. Pharmacol. 1997, 52, 560–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, T.M.; Clay, M.C.; Cioffi, A.G.; Diaz, K.A.; Hisao, G.S.; Tuttle, M.D.; Nieuwkoop, A.J.; Comellas, G.; Maryum, N.; Wang, S.; et al. Amphotericin forms an extramembranous and fungicidal sterol sponge. Nat. Chem. Biol. 2014, 10, 400–406. [Google Scholar] [CrossRef] [PubMed]
- Mesa-Arango, A.C.; Trevijano-Contador, N.; Román, E.; Sánchez-Fresneda, R.; Casas, C.; Herrero, E.; Argüelles, J.C.; Pla, J.; Cuenca-Estrella, M.; Zaragoza, O. The production of reactive oxygen species is a universal action mechanism of amphotericin B against pathogenic yeasts and contributes to the fungicidal effect of this drug. Antimicrob. Agents Chemother. 2014, 58, 6627–6638. [Google Scholar] [CrossRef] [Green Version]
- Shikanai-Yasuda, M.A. Paracoccidioidomycosis treatment. Rev. Inst. Med. Trop. Sao Paulo 2015, 57, 31–37. [Google Scholar] [CrossRef] [Green Version]
- Queiroz-Telles, F.V.; Peçanha Pietrobom, P.M.; Rosa Júnior, M.; Baptista, R.M.; Peçanha, P.M. New Insights on Pulmonary Paracoccidioidomycosis. Semin. Respir. Crit. Care Med. 2020, 41, 53–68. [Google Scholar] [CrossRef]
- Kato, H.; Hagihara, M.; Yamagishi, Y.; Shibata, Y.; Kato, Y.; Furui, T.; Watanabe, H.; Asai, N.; Koizumi, Y.; Mikamo, H. The evaluation of frequency of nephrotoxicity caused by liposomal amphotericin B. J. Infect. Chemother. 2018, 24, 725–728. [Google Scholar] [CrossRef]
- Groll, A.H.; Rijnders, B.J.A.; Walsh, T.J.; Adler-Moore, J.; Lewis, R.E.; Brüggemann, R.J.M. Clinical Pharmacokinetics, Pharmacodynamics, Safety and Efficacy of Liposomal Amphotericin B. Clin. Infect. Dis. 2019, 68, S260–S274. [Google Scholar] [CrossRef]
- Borges, S.R.; Silva, G.M.; Chambela Mda, C.; Oliveira, R.V.; Costa, R.L.; Wanke, B.; Valle, A.C. Itraconazole vs. trimethoprim-sulfamethoxazole: A comparative cohort study of 200 patients with paracoccidioidomycosis. Med. Mycol. 2014, 52, 303–310. [Google Scholar] [CrossRef] [Green Version]
- Shikanai-Yasuda, M.A.; Benard, G.; Higaki, Y.; Del Negro, G.M.; Hoo, S.; Vaccari, E.H.; Gryschek, R.C.; Segurado, A.A.; Barone, A.A.; Andrade, D.R. Randomized trial with itraconazole, ketoconazole and sulfadiazine in paracoccidioidomycosis. Med. Mycol. 2002, 40, 411–417. [Google Scholar] [CrossRef]
- Menezes, V.M.; Soares, B.G.; Fontes, C.J. Drugs for treating paracoccidioidomycosis. Cochrane Database Syst Rev. 2006, 2006, CD004967. [Google Scholar] [CrossRef]
- Groll, A.H.; Piscitelli, S.C.; Walsh, T.J. Clinical pharmacology of systemic antifungal agents: A comprehensive review of agents in clinical use, current investigational compounds, and putative targets for antifungal drug development. Adv. Pharmacol. 1998, 44, 343–500. [Google Scholar] [CrossRef] [PubMed]
- Peyton, L.R.; Gallagher, S.; Hashemzadeh, M. Triazole antifungals: A review. Drugs Today Barc. 2015, 51, 705–718. [Google Scholar] [CrossRef] [PubMed]
- Naranjo, M.S.; Trujillo, M.; Munera, M.I.; Restrepo, P.; Gomez, I.; Restrepo, A. Treatment of paracoccidioidomycosis with itraconazole. J. Med. Vet. Mycol. 1990, 28, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Goldani, L.Z.; Wirth, F. Animal Models and Antifungal Agents in Paracoccidioidomycosis: An Overview. Mycopathologia 2017, 182, 633–643. [Google Scholar] [CrossRef]
- Shafiei, M.; Peyton, L.; Hashemzadeh, M.; Foroumadi, A. History of the development of antifungal azoles: A review on structures, SAR, and mechanism of action. Bioorg. Chem. 2020, 104, 104240. [Google Scholar] [CrossRef]
- Abadio, A.K.; Kioshima, E.S.; Teixeira, M.M.; Martins, N.F.; Maigret, B.; Felipe, M.S. Comparative genomics allowed the identification of drug targets against human fungal pathogens. BMC Genom. 2011, 12, 75. [Google Scholar] [CrossRef] [Green Version]
- Santos, L.A.; Grisolia, J.C.; Burger, E.; Paula, F.B.A.; Amanda Latércia Tranches Dias, A.L.T.; Malaquias, L.C.C. Virulence factors of Paracoccidioides brasiliensis as therapeutic targets: A review. Antonie Leeuwenhoek 2020, 113, 593–604. [Google Scholar] [CrossRef]
- Rodríguez-Brito, S.; Niño-Vega, G.; San-Blas, G. Caspofungin Affects Growth of Paracoccidioides brasiliensis in Both Morphological Phases. Antimicrob. Agents Chemother. 2010, 54, 5391–5394. [Google Scholar] [CrossRef] [Green Version]
- Nakai, T.; Uno, J.; Ikeda, F.; Tawara, S.; Nishimura, K.; Miyaji, M. In Vitro Antifungal Activity of Micafungin (FK463) against Dimorphic Fungi: Comparison of Yeast-Like and Mycelial Forms. Antimicrob. Agents Chemother. 2003, 47, 1376–1381. [Google Scholar] [CrossRef] [Green Version]
- Espinel-Ingroff, A. In vitro antifungal activities of anidulafungin and micafungin, licensed agents and the investigational triazole posaconazole as determined by NCCLS methods for 12,052 fungal isolates: Review of the literature. Rev. Iberoam. Micol. 2003, 20, 121–136. [Google Scholar] [PubMed]
- Lima, S.L.; Colombo, A.L.; de Almeida Junior, J.N. Fungal Cell Wall: Emerging Antifungals and Drug Resistance. Front. Microbiol. 2019, 10, 2573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Effron, G. Rezafungin-Mechanisms of Action, Susceptibility and Resistance: Similarities and Differences with the Other Echinocandins. J. Fungi 2020, 6, 262. [Google Scholar] [CrossRef] [PubMed]
- Schenone, M.; Dančík, V.; Wagner, B.K.; Clemons, P.A. Target identification and mechanism of action in chemical biology and drug discovery. Nat. Chem. Biol. 2013, 9, 232–240. [Google Scholar] [CrossRef] [Green Version]
- Eder, J.; Herrling, P.L. Trends in Modern Drug Discovery. Handb. Exp. Pharmacol. 2016, 232, 3–22. [Google Scholar] [CrossRef]
- Nicola, A.M.; Albuquerque, P.; Paes, H.C.; Fernandes, L.; Costa, F.F.; Kioshima, E.S.; Abadio, A.K.R.; Bocca, A.L.; Felipe, M.S. Antifungal drugs: New insights in research & development. Pharmacol. Ther. 2019, 195, 21–38. [Google Scholar] [CrossRef]
- Batool, M.; Ahmad, B.; Choi, S. A Structure-Based Drug Discovery Paradigm. Int. J. Mol. Sci. 2019, 20, 2783. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Yella, J.; Jegga, A.G. Transcriptomic Data Mining and Repurposing for Computational Drug Discovery. Methods Mol. Biol. 2019, 1903, 73–95. [Google Scholar] [CrossRef]
- Liu, N.; Wang, C.; Su, H.; Zhang, W.; Sheng, C. Strategies in the discovery of novel antifungal scaffolds. Future Med. Chem. 2016, 8, 1435–1454. [Google Scholar] [CrossRef]
- Gidaro, M.C.; Alcaro, S.; Secci, D.; Rivanera, D.; Mollica, A.; Agamennone, M.; Giampietro, L.; Carradori, S. Identification of new anti-Candida compounds by ligand-based pharmacophore virtual screening. J. Enzyme Inhib. Med. Chem. 2016, 31, 1703–1706. [Google Scholar] [CrossRef] [Green Version]
- Xia, X. Bioinformatics and Drug Discovery. Curr. Top. Med. Chem. 2017, 17, 1709–1726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karatzas, E.; Kolios, G.; Spyrou, G.M. An Application of Computational Drug Repurposing Based on Transcriptomic Signatures. Methods Mol. Biol. 2019, 1903, 149–177. [Google Scholar] [CrossRef] [PubMed]
- Huo, J.; Zhao, B.; Zhang, Z.; Xing, J.; Zhang, J.; Dong, J.; Fan, Z. Structure-Based Discovery and Synthesis of Potential Transketolase Inhibitors. Molecules 2018, 23, 2116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakheet, T.M.; Doig, A.J. Properties and identification of human protein drug targets. Bioinformatics 2009, 25, 451–457. [Google Scholar] [CrossRef] [Green Version]
- Roemer, T.; Jiang, B.; Davison, J.; Ketela, T.; Veillette, K.; Breton, A.; Tandia, F.; Linteau, A.; Sillaots, S.; Marta, C.; et al. Large-scale essential gene identification in Candida albicans and applications to antifungal drug discovery. Mol. Microbiol. 2003, 50, 167–181. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Sillaots, S.; Lemieux, S.; Davison, J.; Kauffman, S.; Breton, A.; Linteau, A.; Xin, C.; Bowman, J.; Becker, J.; et al. Essential gene identification and drug target prioritization in Aspergillus fumigatus. PLoS Pathog. 2007, 3, e24. [Google Scholar] [CrossRef]
- Buurman, E.T.; Westwater, C.; Hube, B.; Brown, A.J.; Odds, F.C.; Gow, N.A. Molecular analysis of CaMnt1p, a mannosyl transferase important for adhesion and virulence of Candida albicans. Proc. Natl. Acad. Sci. USA 1998, 95, 7670–7675. [Google Scholar] [CrossRef] [Green Version]
- Jensen-Pergakes, K.L.; Kennedy, M.A.; Lees, N.D.; Barbuch, R.; Koegel, C.; Bard, M. Sequencing, disruption, and characterization of the Candida albicans sterol methyltransferase (ERG6) gene: Drug susceptibility studies in erg6 mutants. Antimicrob. Agents Chemother. 1998, 42, 1160–1167. [Google Scholar] [CrossRef] [Green Version]
- Perfect, J.R.; Tenor, J.L.; Miao, Y.; Brennan, R.G. Trehalose pathway as an antifungal target. Virulence 2017, 8, 143–149. [Google Scholar] [CrossRef] [Green Version]
- Freitas, E.; Silva, K.S.C.; Silva, L.; Gonçales, R.A.; Neves, B.J.; Soares, C.M.A.; Pereira, M. Setting New Routes for Antifungal Drug Discovery Against Pathogenic Fungi. Curr. Pharm. Des. 2020, 26, 1509–1520. [Google Scholar] [CrossRef]
- Yoshida, T.; Oka, S.; Masutani, H.; Nakamura, H.; Yodoi, J. The role of thioredoxin in the aging process: Involvement of oxidative stress. Antioxid Redox Signal. 2003, 5, 563–570. [Google Scholar] [CrossRef] [PubMed]
- Holmgren, A. Thioredoxin. Annu. Rev. Biochem. 1985, 54, 237–271. [Google Scholar] [CrossRef]
- Moore, E.C.; Reichard, P.; Thelander, L. Enzymatic synthesis of deoxyribonucleotides.v. purification and properties of thioredoxin reductase from Escherichia coli b. J. Biol.Chem. 1964, 239, 3445–3452. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.H., Jr. Mechanism and structure of thioredoxin reductase from Escherichia coli. FASEB J. 1995, 9, 1267–1276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnér, E.S.; Holmgren, A. Physiological functions of thioredoxin and thioredoxin reductase. Eur. J. Biochem. 2000, 267, 6102–6109. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Zou, L.; Lu, J.; Holmgren, A. Selenocysteine in mammalian thioredoxin reductase and application of ebselen as a therapeutic. Free Radic. Biol. Med. 2018, 127, 238–247. [Google Scholar] [CrossRef]
- Zhang, B.; Zhang, J.; Peng, S.; Liu, R.; Li, X.; Hou, Y.; Han, X.; Fang, J. Thioredoxin reductase inhibitors: A patent review. Expert Opin. Ther. Pat. 2017, 27, 547–556. [Google Scholar] [CrossRef]
- Lu, J.; Holmgren, A. The thioredoxin antioxidant system. Free Radic. Biol. Med. 2014, 66, 75–87. [Google Scholar] [CrossRef]
- Saccoccia, F.; Angelucci, F.; Boumis, G.; Carotti, D.; Desiato, G.; Miele, A.E.; Bellelli, A. Thioredoxin reductase and its inhibitors. Curr. Protein Pept. Sci. 2014, 15, 621–646. [Google Scholar] [CrossRef] [Green Version]
- Arnér, E.S.J. Selective Evaluation of Thioredoxin Reductase Enzymatic Activities. Methods Mol. Biol. 2018, 1661, 301–309. [Google Scholar] [CrossRef]
- Novoselov, S.V.; Gladyshev, V.N. Non-animal origin of animal thioredoxin reductases: Implications for selenocysteine evolution and evolution of protein function through carboxy-terminal extensions. Protein Sci. 2003, 12, 372–378. [Google Scholar] [CrossRef] [PubMed]
- Benhar, M. Roles of mammalian glutathione peroxidase and thioredoxin reductase enzymes in the cellular response to nitrosative stress. Free Radic. Biol. Med. 2018, 127, 160–164. [Google Scholar] [CrossRef]
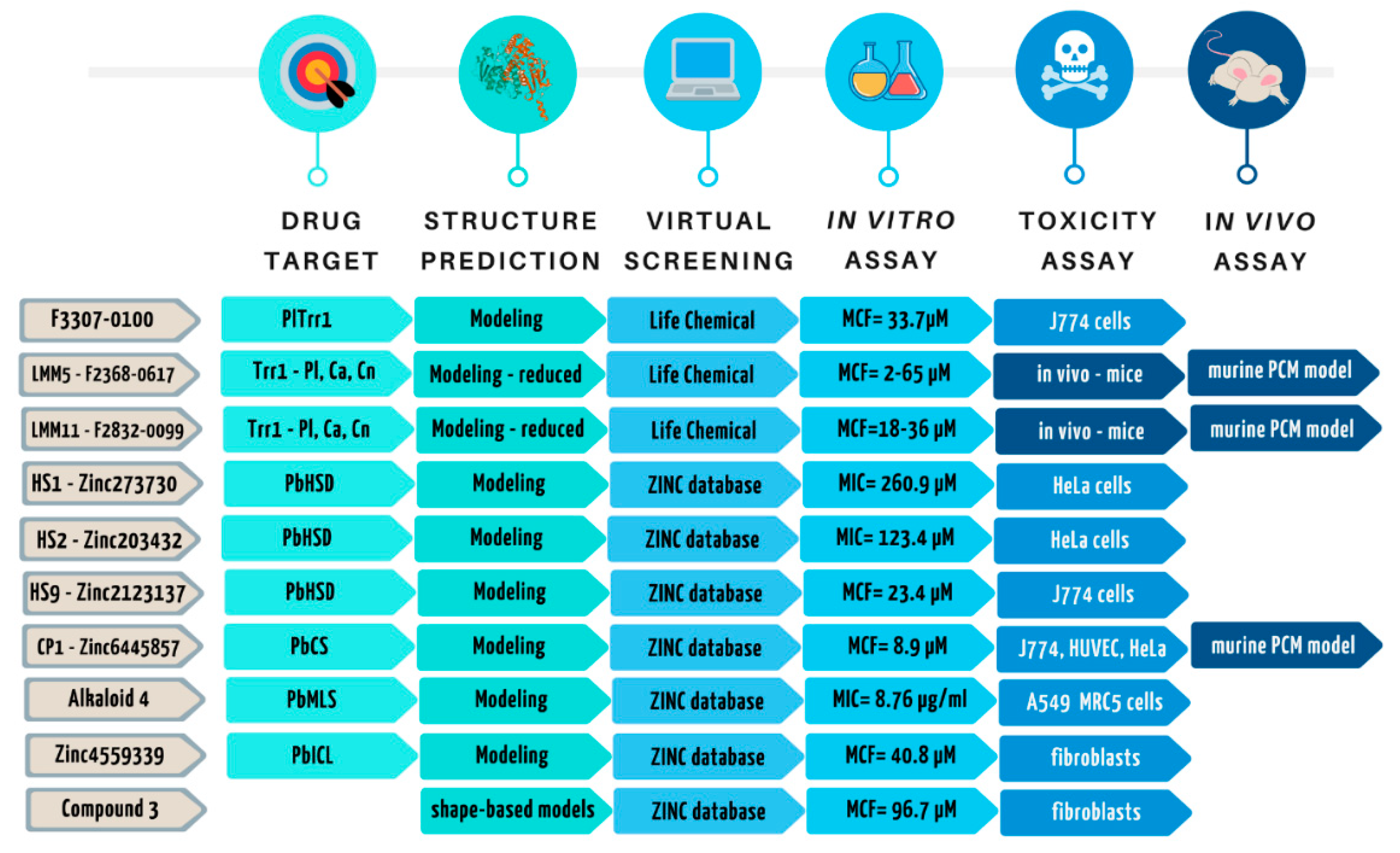
- Abadio, A.K.; Kioshima, E.S.; Leroux, V.; Martins, N.F.; Maigret, B.; Felipe, M.S. Identification of new antifungal compounds targeting thioredoxin reductase of Paracoccidioides genus. PLoS ONE 2015, 10, e0142926. [Google Scholar] [CrossRef]
- Gromer, S.; Urig, S.; Becker, K. The thioredoxin system—From science to clinic. Med. Res. Rev. 2004, 24, 40–89. [Google Scholar] [CrossRef]
- Arnér, E.S.; Holmgren, A. The thioredoxin system in cancer. Semin. Cancer Biol. 2006, 16, 420–426. [Google Scholar] [CrossRef]
- Sun, Q.A.; Su, D.; Novoselov, S.V.; Carlson, B.A.; Hatfield, D.L.; Gladyshev, V.N. Reaction mechanism and regulation of mammalian thioredoxin/glutathione reductase. Biochemistry 2005, 44, 14528–14537. [Google Scholar] [CrossRef]
- Lillig, C.H.; Holmgren, A. Thioredoxin and related molecules—From biology to health and disease. Antioxid. Redox Signal. 2007, 9, 25–47. [Google Scholar] [CrossRef]
- Pedrajas, J.R.; Kosmidou, E.; Miranda-Vizuete, A.; Gustafsson, J.A.; Wright, A.P.; Spyrou, G. Identification and functional characterization of a novel mitochondrial thioredoxin system in Saccharomyces cerevisiae. J. Biol. Chem. 1999, 274, 6366–6373. [Google Scholar] [CrossRef] [Green Version]
- Trotter, E.W.; Grant, C.M. Overlapping roles of the cytoplasmic and mitochondrial redox regulatory systems in the yeast Saccharomyces cerevisiae. Eukaryot. Cell. 2005, 4, 392–400. [Google Scholar] [CrossRef] [Green Version]
- Raffel, J.; Bhattacharyya, A.K.; Gallegos, A.; Cui, H.; Einspahr, J.G.; Alberts, D.S.; Powis, G. Increased expression of thioredoxin-1 in human colorectal cancer is associated with decreased patient survival. J. Lab. Clin. Med. 2003, 142, 46–51. [Google Scholar] [CrossRef]
- Gasdaska, P.Y.; Oblong, J.E.; Cotgreave, I.A.; Powis, G. The predicted amino acid sequence of human thioredoxin is identical to that of the autocrine growth factor human adult T-cell derived factor (ADF): Thioredoxin mRNA is elevated in some human tumors. Biochim. Biophys. Acta 1994, 1218, 292–296. [Google Scholar] [CrossRef]
- Nakamura, H.; Bai, J.; Nishinaka, Y.; Ueda, S.; Sasada, T.; Ohshio, G.; Imamura, M.; Takabayashi, A.; Yamaoka, Y.; Yodoi, J. Expression of thioredoxin and glutaredoxin, redox-regulating proteins, in pancreatic cancer. Cancer Detect. Prev. 2000, 24, 53–60. [Google Scholar] [PubMed]
- Lim, J.Y.; Yoon, S.O.; Hong, S.W.; Kim, J.W.; Choi, S.H.; Cho, J.Y. Thioredoxin and thioredoxin-interacting protein as prognostic markers for gastric cancer recurrence. World J. Gastroenterol. 2012, 18, 5581–5588. [Google Scholar] [CrossRef] [PubMed]
- Stafford, W.C.; Peng, X.; Olofsson, M.H.; Zhang, X.; Luci, D.K.; Lu, L.; Cheng, Q.; Trésaugues, L.; Dexheimer, T.S.; Coussens, N.P.; et al. Irreversible inhibition of cytosolic thioredoxin reductase 1 as a mechanistic basis for anticancer therapy. Sci. Transl. Med. 2018, 10, eaaf7444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ianiri, G.; Idnurm, A. Essential gene discovery in the basidiomycete Cryptococcus neoformans for antifungal drug target prioritization. mBio 2015, 6, e02334-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godoy, J.S.; Kioshima, É.S.; Abadio, A.K.; Felipe, M.S.; de Freitas, S.M.; Svidzinski, T.I. Structural and functional characterization of the recombinant thioredoxin reductase from Candida albicans as a potential target for vaccine and drug design. Appl. Microbiol. Biotechnol. 2016, 100, 4015–4025. [Google Scholar] [CrossRef]
- Capoci, I.R.G.; Sakita, K.M.; Faria, D.R.; Rodrigues-Vendramini, F.A.V.; Arita, G.S.; de Oliveira, A.G.; Felipe, M.S.; Maigret, B.; Bonfim-Mendonça, P.S.; Kioshima, E.S.; et al. Two New 1,3,4-Oxadiazoles With Effective Antifungal Activity Against Candida albicans. Front. Microbiol. 2019, 10, 2130. [Google Scholar] [CrossRef]
- De Oliveira, F.F.M.; Paredes, V.; de Sousa, H.R.; D’Áurea Moura, Á.N.; Riasco-Palacios, J.; Casadevall, A.; Felipe, M.S.S.; Nicola, A.M. Thioredoxin Reductase 1 Is a Highly Immunogenic Cell Surface Antigen in Paracoccidioides spp., Candida albicans, and Cryptococcus neoformans. Front. Microbiol. 2020, 10, 2930. [Google Scholar] [CrossRef]
- Faria, D.R.; Sakita, K.M.; Capoci, I.R.G.; Arita, G.S.; Rodrigues-Vendramini, F.A.V.; de Oliveira Junior, A.G.; Soares Felipe, M.S.; Bonfim de Mendonça, P.S.; Svidzinski, T.I.E.; Kioshima, E.S. Promising antifungal activity of new oxadiazole against Candida krusei. PLoS ONE 2020, 15, e0227876. [Google Scholar] [CrossRef]
- Binder, J.; Shadkchan, Y.; Osherov, N.; Krappmann, S. The Essential Thioredoxin Reductase of the Human Pathogenic Mold Aspergillus fumigatus is a Promising Antifungal Target. Front. Microbiol. 2020, 11, 1383. [Google Scholar] [CrossRef]
- Missall, T.A.; Lodge, J.K. Thioredoxin reductase is essential for viability in the fungal pathogen Cryptococcus neoformans. Eukaryot Cell 2005, 4, 487–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bravo-Chaucanés, C.P.; Abadio, A.K.R.; Kioshima, É.S.; Felipe, M.S.S.; Barbosa, J.A.R.G. Crystal structure of thioredoxin 1 from Cryptococcus neoformans at 1.8 Å resolution shows unexpected plasticity of the loop preceding the catalytic site. Biochem. Biophys. Rep. 2020, 21, 100724. [Google Scholar] [CrossRef]
- Marshall, A.C.; Kidd, S.E.; Lamont-Friedrich, S.J.; Arentz, G.; Hoffmann, P.; Coad, B.R.; Bruning, J.B. Structure, Mechanism, and Inhibition of Aspergillus fumigatus Thioredoxin Reductase. Antimicrob. Agents Chemother. 2019, 63, e02281-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues-Vendramini, F.A.V.; Faria, D.R.; Arita, G.S.; Capoci, I.R.G.; Sakita, K.M.; Caparroz-Assef, S.M.; Becker, T.C.A.; Bonfim-Mendonça, P.S.; Felipe, M.S.; Svidzinski, T.I.E.; et al. Antifungal activity of two oxadiazole compounds for the paracoccidioidomycosis treatment. PLoS Negl. Trop. Dis. 2019, 13, e0007441. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Li, X.; Lin, X. A Review on Applications of Computational Methods in Drug Screening and Design. Molecules 2020, 25, 1375. [Google Scholar] [CrossRef] [Green Version]
- Franco, P.H.C.; Braga, S.F.P.; de Oliveira, R.B.; César, I.C. Purity determination of a new antifungal drug candidate using quantitative 1 H NMR spectroscopy: Method validation and comparison of calibration approaches. Magn. Reson. Chem. 2020, 58, 97–105. [Google Scholar] [CrossRef]
- Tighadouini, S.; Radi, S.; Abrigach, F.; Benabbes, R.; Eddike, D.; Tillard, M. Novel β-keto–enol Pyrazolic Compounds as Potent Antifungal Agents. Design, Synthesis, Crystal Structure, DFT, Homology Modeling, and Docking Studies. J. Chem. Inf. Model. 2019, 59, 1398–1409. [Google Scholar] [CrossRef]
- Coitinho, J.B.; Costa, M.A.; Melo, E.M.; Morais, E.A.; de Andrade, L.G.; da Rocha, A.M.; Magalhães, M.T.Q.; Favaro, D.C.; Bleicher, L.; Pedroso, E.R.P.; et al. Structural and immunological characterization of a new nucleotidyltransferase-like antigen from Paracoccidioides brasiliensis. Mol. Immunol. 2019, 112, 151–162. [Google Scholar] [CrossRef]
- Bagatin, M.C.; Pimentel, A.L.; Biavatti, D.C.; Basso, E.A.; Kioshima, E.S.; Seixas, F.A.V.; Gauze, G.F. Targeting the Homoserine Dehydrogenase of Paracoccidioides Species for Treatment of Systemic Fungal Infections. Antimicrob. Agents Chemother. 2017, 61, e00165-17. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues-Vendramini, F.A.V.; Marschalk, C.; Toplak, M.; Macheroux, P.; Bonfim-Mendonça, P.S.; Svidzinski, T.I.E.; Seixas, F.A.V.; Kioshima, E.S. Promising New Antifungal Treatment Targeting Chorismate Synthase from Paracoccidioides brasiliensis. Antimicrob. Agents Chemother. 2018, 63, e01097-18. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, L.S.; Barbosa, U.R.; Silva, L.D.C.; Soares, C.M.; Pereira, M.; da Silva, R.A. Identification of a new antifungal compound against isocitrate lyase of Paracoccidioides brasiliensis. Future Microbiol. 2019, 14, 1589–1606. [Google Scholar] [CrossRef] [PubMed]
- Beautrait, A.; Leroux, V.; Chavent, M.; Ghemtio, L.; Devignes, M.D.; Smaïl-Tabbone, M.; Cai, W.; Shao, X.; Moreau, G.; Bladon, P.; et al. Multiple-step virtual screening using VSM-G: Overview and validation of fast geometrical matching enrichment. J. Mol. Model. 2008, 14, 135–148. [Google Scholar] [CrossRef]
- Ghemtio, L.; Devignes, M.D.; Smail-Tabbone, M.; Souchet, M.; Leroux, V.; Maigret, B. Comparison of three preprocessing filters efficiency in virtual screening: Identification of new putative LXR beta regulators as a test case. J. Chem. Inf. Model. 2010, 50, 701–715. [Google Scholar] [CrossRef] [PubMed]
- Oum, Y.H.; Kell, S.A.; Yoon, Y.; Liang, Z.; Burger, P.; Shim, H. Discovery of novel aminopiperidinyl amide CXCR4 modulators through virtual screening and rational drug design. Eur. J. Med. Chem. 2020, 201, 112479. [Google Scholar] [CrossRef] [PubMed]
- Costa, F.G.; Neto, B.R.; Gonçalves, R.L.; da Silva, R.A.; de Oliveira, C.M.; Kato, L.; Freitas, C.S.; Giannini, M.J.; da Silva, J.F.; Soares, C.M.; et al. Alkaloids as inhibitors of malate synthase from Paracoccidioides spp.: Receptor-ligand interaction-based virtual screening and molecular docking studies, antifungal activity, and the adhesion process. Antimicrob. Agents Chemother. 2015, 59, 5581–5594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, H.; Liu, H.; Kuang, Y.; Chen, L.; Ye, M.; Lai, L. Discovery of Targeted Covalent Natural Products against PLK1 by Herb-Based Screening. J. Chem. Inf. Model. 2020, 60, 4350–4358. [Google Scholar] [CrossRef] [PubMed]
- Hughes, J.P.; Rees, S.; Kalindjian, S.B.; Philpott, K.L. Principles of early drug discovery. Br. J. Pharmacol. 2011, 162, 1239–1249. [Google Scholar] [CrossRef] [Green Version]
- DiMasi, J.A.; Grabowski, H.G.; Hansen, R.W. Innovation in the pharmaceutical industry: New estimates of R&D costs. J. Health Econ. 2016, 47, 20–33. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, M.A.; Discola, K.F.; Alves, S.V.; Barbosa, J.A.; Medrano, F.J.; Netto, L.E.; Guimarães, B.G. Crystallization and preliminary X-ray diffraction analysis of NADPH-dependent thioredoxin reductase I from Saccharomyces cerevisiae. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2005, 61, 387–390. [Google Scholar] [CrossRef] [Green Version]
- Lennon, B.W.; Williams, C.H.; Ludwig, M.L. Twists in catalysis: Alternating conformations of Escherichia coli thioredoxin reductase. Science 2000, 289, 1190–1194. [Google Scholar] [CrossRef]
- Kioshima, E.S.; Abadio, A.K.R.; Leurox, V.; Capoci, I.R.G.; Bonfim-Mendonça, P.S.; Felipe, M.S.; Svidzinski, T.I.E.; Maigret, B. Thioredoxin Systems as Target to New Antifungal Development. Status manuscript in preparation.
- Bueno, P.S.A.; Rodrigues, F.A.V.; Santos, J.L.; Canduri, F.; Biavatti, D.C.; Pimentel, A.L.; Bagatin, M.C.; Kioshima, É.S.; de Freitas Gauze, G.; Seixas, F.A.V. New inhibitors of homoserine dehydrogenase from Paracoccidioides brasiliensis presenting antifungal activity. J. Mol. Model. 2019, 25, 325. [Google Scholar] [CrossRef] [PubMed]
- Rozada, A.M.; Rodrigues, F.A.; Sampiron, E.G.; Seixas, F.A.; Basso, E.A.; Scodro, R.B.; Kioshima, É.S.; Gauze, G.F. Novel 4-methoxynaphthalene-N-acylhydrazones as potential for paracoccidioidomycosis and tuberculosis co-infection. Future Microbiol. 2019, 14, 587–598. [Google Scholar] [CrossRef] [PubMed]
- Rozada, A.M.F.; Rodrigues-Vendramini, F.A.V.; Gonçalves, D.S.; Rosa, F.A.; Basso, E.A.; Seixas, F.A.V.; Kioshima, É.S.; Gauze, G.F. Synthesis and antifungal activity of new hybrids pyrimido[4,5-d]pyridazinone-N-acylhydrazones. Bioorg. Med. Chem. Lett. 2020, 30, 127244. [Google Scholar] [CrossRef] [PubMed]
- Silva, P.A.; Abadio, A.K.R.; Kioshima, E.S.; Leroux, V.; Martins, N.F.; Nahum, V.H.; Mora-Montes, H.M.; Maigret, B.; Felipe, M.S.S. Inhibitors of α-1,2-mannosyltransferase as Promising Antifungal Activity against The Invasive Fungal Infections Agents. Status manuscript in preparation.
- Salci, T.P.; Negri, M.; Abadio, A.K.R.; Bonfim-Mendonça, P.S.; Capoci, I.R.G.; Caparroz-Assef, S.M.; Donati, L.; Felipe, M.S.S.; Kioshima, E.S.; Svidzinski, T.I.E. A new small molecule KRE2 inhibitor against invasive Candida parapsilosis infection. Future Microbiol. 2017, 12, 1283–1295. [Google Scholar] [CrossRef] [PubMed]
- Silva, L.C.; Neves, B.J.; Gomes, M.N.; Melo-Filho, C.C.; Soares, C.M.; Andrade, C.H.; Pereira, M. Computer-aided identification of novel anti-paracoccidioidomycosis compounds. Future Microbiol. 2018, 13, 1523–1535. [Google Scholar] [CrossRef] [PubMed]
- Ashburn, T.T.; Thor, K.B. Drug repositioning: Identifying and developing new uses for existing drugs. Nat. Rev. Drug Discov. 2004, 3, 673–683. [Google Scholar] [CrossRef]
- Butts, A.; Krysan, D.J. Antifungal drug discovery: Something old and something new. PLoS Pathog. 2012, 8, e1002870. [Google Scholar] [CrossRef]
- Katragkou, A.; Roilides, E.; Walsh, T.J. Can repurposing of existing drugs provide more effective therapies for invasive fungal infections? Expert Opin. Pharmacother. 2016, 17, 1179–1182. [Google Scholar] [CrossRef] [Green Version]
- FDA approvals for the first 6 months of 2014. Nat. Rev. Drug. Discov. 2014, 13, 565. [CrossRef]
- Zerbini, L.F.; Bhasin, M.K.; de Vasconcellos, J.F.; Paccez, J.D.; Gu, X.; Kung, A.L.; Libermann, T.A. Computational repositioning and preclinical validation of pentamidine for renal cell cancer. Mol. Cancer Ther. 2014, 13, 1929–1941. [Google Scholar] [CrossRef] [Green Version]
- Wiederhold, N.P.; Patterson, T.F.; Srinivasan, A.; Chaturvedi, A.K.; Fothergill, A.W.; Wormley, F.L.; Ramasubramanian, A.K.; Lopez-Ribot, J.L. Repurposing auranofin as an antifungal: In vitro activity against a variety of medically important fungi. Virulence 2017, 8, 138–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ekins, S.; Williams, A.J.; Krasowski, M.D.; Freundlich, J.S. In silico repositioning of approved drugs for rare and neglected diseases. Drug Discov. Today 2011, 16, 298–310. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Zilbermintz, L.; Martchenko, M. Repurposing FDA approved drugs against the human fungal pathogen, Candida albicans. Ann. Clin. Microbiol. Antimicrob. 2015, 14, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nixon, G.L.; McEntee, L.; Johnson, A.; Farrington, N.; Whalley, S.; Livermore, J.; Natal, C.; Washbourn, G.; Bibby, J.; Berry, N.; et al. Repurposing and Reformulation of the Antiparasitic Agent Flubendazole for Treatment of Cryptococcal Meningoencephalitis, a Neglected Fungal Disease. Antimicrob. Agents Chemother. 2018, 62, e01909-17. [Google Scholar] [CrossRef] [Green Version]
- Capoci, I.R.G.; Faria, D.R.; Sakita, K.M.; Rodrigues-Vendramini, F.A.V.; Bonfim-Mendonça, P.S.; Becker, T.C.A.; Kioshima, É.S.; Svidzinski, T.I.E.; Maigret, B. Repurposing approach identifies new treatment options for invasive fungal disease. Bioorganic Chem. 2019, 84, 87–97. [Google Scholar] [CrossRef]
- De Oliveira, A.A.; Neves, B.J.; do Silva, L.C.; de Soares, C.M.A.; Andrade, C.H.; Pereira, M. Drug Repurposing for Paracoccidioidomycosis Through a Computational Chemogenomics Framework. Front. Microbiol. 2019, 10, 1301. [Google Scholar] [CrossRef]
- Caffrey, C.R.; Rohwer, A.; Oellien, F.; Marhöfer, R.J.; Braschi, S.; Oliveira, G.; McKerrow, J.H.; Selzer, P.M. A comparative chemogenomics strategy to predict potential drug targets in the metazoan pathogen, Schistosoma mansoni. PLoS ONE 2009, 4, e4413. [Google Scholar] [CrossRef]
- Bredel, M.; Jacoby, E. Chemogenomics: An emerging strategy for rapid target and drug discovery. Nat. Rev. Genet. 2004, 5, 262–275. [Google Scholar] [CrossRef] [Green Version]
- Neves, B.J.; Braga, R.C.; Bezerra, J.C.; Cravo, P.V.; Andrade, C.H. In silico repositioning-chemogenomics strategy identifies new drugs with potential activity against multiple life stages of Schistosoma mansoni. PLoS Negl. Trop. Dis. 2015, 9, e3435. [Google Scholar] [CrossRef]
- Arif, T.; Bhosale, J.D.; Kumar, N.; Mandal, T.K.; Bendre, R.S.; Lavekar, G.S.; Dabur, R. Natural products-antifungal agents derived from plants. J. Asian Nat. Prod. Res. 2009, 11, 621–638. [Google Scholar] [CrossRef] [PubMed]
- San-Blas, G.; Mariño, L.; San-Blas, F.; Apitz-Castro, R. Effect of ajoene on dimorphism of Paracoccidioides brasiliensis. J. Med. Vet. Mycol. 1993, 31, 133–141. [Google Scholar] [CrossRef] [PubMed]
- San-Blas, G.; Urbina, J.A.; Marchán, E.; Contreras, L.M.; Sorais, F.; San-Blas, F. Inhibition of Paracoccidioides brasiliensis by ajoene is associated with blockade of phosphatidylcholine biosynthesis. Microbiology 1997, 143, 1583–1586. [Google Scholar] [CrossRef] [Green Version]
- Maluf, M.L.; Takahachi, G.; Svidzinski, T.I.; Xander, P.; Apitz-Castro, R.; Bersani-Amado, C.A.; Cuman, R.K. Antifungal activity of ajoene on experimental murine paracoccidioidomycosis. Rev. Iberoam. Micol. 2008, 25, 163–166. [Google Scholar] [CrossRef]
- Thomaz, L.; Apitz-Castro, R.; Marques, A.F.; Travassos, L.R.; Taborda, C.P. Experimental paracoccidioidomycosis: Alternative therapy with ajoene, compound from Allium sativum, associated with sulfamethoxazole/trimethoprim. Med. Mycol. 2008, 46, 113–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, G.D.; Ferri, P.H.; Santos, S.C.; Bao, S.N.; Soares, C.M.; Pereira, M. Oenothein B inhibits the expression of PbFKS1 transcript and induces morphological changes in Paracoccidioides brasiliensis. Med. Mycol. 2007, 45, 609–618. [Google Scholar] [CrossRef] [PubMed]
- Zambuzzi-Carvalho, P.F.; Tomazett, P.K.; Santos, S.C.; Ferri, P.H.; Borges, C.L.; Martins, W.S.; Soares, C.M.A.; Pereira, M. Transcriptional profile of Paracoccidioides induced by oenothein B, a potential antifungal agent from the Brazilian Cerrado plant Eugenia uniflora. BMC Microbiol. 2013, 13, 227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Freitas, C.S.; Kato, L.; de Oliveira, C.M.; Queiroz, L.H., Jr.; Santana, M.J.; Schuquel, I.T.; Delprete, P.G.; da Silva, R.A.; Quintino, G.O.; da Silva Neto, B.R.; et al. β-Carboline alkaloids from Galianthe ramosa inhibit malate synthase from Paracoccidioides spp. Planta Med. 2014, 80, 1746–1752. [Google Scholar] [CrossRef] [PubMed]
- Zambuzzi-Carvalho, P.F.; Cruz, A.H.; Santos-Silva, L.K.; Goes, A.M.; Soares, C.M.; Pereira, M. The malate synthase of Paracoccidioides brasiliensis Pb01 is required in the glyoxylate cycle and in the allantoin degradation pathway. Med. Mycol. 2009, 47, 734–744. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira, K.M.; da Silva Neto, B.R.; Parente, J.A.; da Silva, R.A.; Quintino, G.O.; Voltan., A.R.; Mendes-Giannini, M.J.S.; Soares, C.M.A.; Pereira, M. Intermolecular interactions of the malate synthase of Paracoccidioides spp. BMC Microbiol. 2013, 13, 107. [Google Scholar] [CrossRef] [Green Version]
- Prado, R.S.; Alves, R.J.; Oliveira, C.M.; Kato, L.; Silva, R.A.; Quintino, G.O.; do Desterro Cunha, S.; Soares, C.M.A.; Pereira, M. Inhibition of Paracoccidioides lutzii Pb01 isocitrate lyase by the natural compound argentilactone and its semi-synthetic derivatives. PLoS ONE 2014, 9, e94832. [Google Scholar] [CrossRef]
- Cruz, A.H.; Brock, M.; Zambuzzi-Carvalho, P.F.; Santos-Silva, L.K.; Troian, R.F.; Góes, A.M.; Soares, C.M.; Pereira, M. Phosphorylation is the major mechanism regulating isocitrate lyase activity in Paracoccidioides brasiliensis yeast cells. FEBS J. 2011, 278, 2318–2332. [Google Scholar] [CrossRef] [PubMed]
- Do Carmo Silva, L.; Tamayo Ossa, D.P.; Castro, S.V.; Bringel Pires, L.; Alves de Oliveira, C.M.; Conceição da Silva, C.; Coelho, N.P.; Bailão, A.M.; Parente-Rocha, J.A.; Soares, C.M.; et al. Transcriptome Profile of the Response of Paracoccidioides spp. to a Camphene Thiosemicarbazide Derivative. PLoS ONE 2015, 10, e0130703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borba, J.V.V.B.; Tauhata, S.B.F.; Oliveira, C.M.A.; Ferreira Marques, M.; Bailão, A.M.; Soares, C.M.A.; Pereira, M. Chemoproteomic identification of molecular targets of antifungal prototypes, thiosemicarbazide and a camphene derivative of thiosemicarbazide, in Paracoccidioides brasiliensis. PLoS ONE 2018, 13, e0201948. [Google Scholar] [CrossRef] [PubMed]
- Amaral, A.C.; Marques, A.F.; Muñoz, J.E.; Bocca, A.L.; Simioni, A.R.; Tedesco, A.C.; Morais, P.C.; Travassos, L.R.; Taborda, C.P.; Felipe, M.S. Poly(lactic acid-glycolic acid) nanoparticles markedly improve immunological protection provided by peptide P10 against murine paracoccidioidomycosis. Br. J. Pharmacol. 2010, 159, 1126–1132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos Junior, S.R.; Silva, F.K.L.; Dias, L.S.; Souza, A.C.O.; Araujo, M.V.; Silva, L.B.R.; Travassos, L.R.; Amaral, A.C.; Taborda, C.P. Intranasal Vaccine Using P10 Peptide Complexed within Chitosan Polymeric Nanoparticles as Experimental Therapy for Paracoccidioidomycosis in Murine Model. J. Fungi 2020, 6, 160. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, A.M.; Souza, A.C.; Amaral, A.C.; Vasconcelos, N.M.; Jeronimo, M.S.; Carneiro, F.P.; Faccioli, L.H.; Felipe, M.S.; Silva, C.L.; Bocca, A.L. Nanobiotechnological approaches to delivery of DNA vaccine against fungal infection. J. Biomed. Nanotechnol. 2013, 9, 221–230. [Google Scholar] [CrossRef]
- Jannuzzi, G.P.; Souza, N.A.; Françoso, K.S.; Pereira, R.H.; Santos, R.P.; Kaihami, G.H.; Almeida, J.R.F.; Batista, W.L.; Amaral, A.C.; Maranhão, A.Q.; et al. Therapeutic treatment with scFv-PLGA nanoparticles decreases pulmonary fungal load in a murine of paracoccidioidomycosis. Microbes Infect. 2018, 20, 48–56. [Google Scholar] [CrossRef]
- Amaral, A.C.; Bocca, A.L.; Ribeiro, A.M.; Nunes, J.; Peixoto, D.L.; Simioni, A.R.; Primo, F.L.; Lacava, Z.G.; Bentes, R.; Titze-de-Almeida, R.; et al. Amphotericin B in poly(lactic-co-glycolic acid) (PLGA) and dimercaptosuccinic acid (DMSA) nanoparticles against paracoccidioidomycosis. J. Antimicrob. Chemother. 2009, 63, 526–533. [Google Scholar] [CrossRef] [Green Version]
- Saldanha, C.A.; Garcia, M.P.; Iocca, D.C.; Rebelo, L.G.; Souza, A.C.; Bocca, A.L.; Almeida Santos, M.F.; Morais, P.C.; Azevedo, R.B. Antifungal Activity of Amphotericin B Conjugated to Nanosized Magnetite in the Treatment of Paracoccidioidomycosis. PLoS Negl. Trop. Dis. 2016, 10, e0004754. [Google Scholar] [CrossRef] [Green Version]
- Cunha-Azevedo, E.P.; Py-Daniel, K.R.; Siqueira-Moura, M.P.; Bocca, A.L.; Felipe, M.S.S.; Tedesco, A.C.; Junior, O.R.P.; Lucci, C.M.; Azevedo, R.B. In vivo evaluation of the efficacy, toxicity and biodistribution of PLGA-DMSA nanoparticles loaded with itraconazole for treatment of paracoccidioidomycosis. J. Drug Deliv. Sci. Technol. 2018, 45, 135–141. [Google Scholar] [CrossRef]
- Souza, A.C.; Amaral, A.C. Antifungal Therapy for Systemic Mycosis and the Nanobiotechnology Era: Improving Efficacy, Biodistribution and Toxicity. Front. Microbiol. 2017, 8, 336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farjadian, F.; Ghasemi, A.; Gohari, O.; Roointan, A.; Karimi, M.; Hamblin, M.R. Nanopharmaceuticals and nanomedicines currently on the market: Challenges and opportunities. Nanomed. Lond. 2019, 14, 93–126. [Google Scholar] [CrossRef] [PubMed]
- Amaral, A.C.; Felipe, M.S. Nanobiotechnology: An efficient approach to drug delivery of unstable biomolecules. Curr. Protein Pept. Sci. 2013, 14, 588–594. [Google Scholar] [CrossRef] [PubMed]
- Detsi, A.; Kavetsou, E.; Kostopoulou, I.; Pitterou, I.; Pontillo, A.R.N.; Tzani, A.; Christodoulou, P.; Siliachli, A.; Zoumpoulakis, P. Nanosystems for the Encapsulation of Natural Products: The Case of Chitosan Biopolymer as a Matrix. Pharmaceutics 2020, 12, 669. [Google Scholar] [CrossRef] [PubMed]
- Garcia, M.P.; Parca, R.M.; Chaves, S.B.; Silva, L.P.; Santos, A.D.; Lacava, Z.G.M.; Morais, P.C.; Azevedo, R.B. Morphological analysis of mice’s lung after treatment with magnetite-based magnetic fluid stabilized with DMSA. J. Magn. Magn. Mater. 2005, 293, 277–282. [Google Scholar] [CrossRef]
- Yilma, S.; Liu, N.; Samoylov, A.; Lo, T.; Brinker, C.J.; Vodyanoy, V. Amphotericin B channels in phospholipid membrane-coated nanoporous silicon surfaces: Implications for photovoltaic driving of ions across membranes. Biosens. Bioelectron. 2007, 22, 1605–1611. [Google Scholar] [CrossRef]
- Souza, A.C.; Nascimento, A.L.; de Vasconcelos, N.M.; Jerônimo, M.S.; Siqueira, I.M.; Santos, L.; Cintra, D.O.; Fuscaldi, L.L.; Pires Júnior, O.R.; Titze-de-Almeida, R.; et al. Activity and in vivo tracking of Amphotericin B loaded PLGA nanoparticles. Eur. J. Med. Chem. 2015, 95, 267–276. [Google Scholar] [CrossRef]
- Suneet, K.; Sridhar, S.; Agiwal, P.; Sridhar, M.S.; Sanyal, K.; Jain, S. Magnetic hyperthermia adjunctive therapy for fungi: In vitro studies against Candida albicans. Int. J. Hyperth. 2019, 36, 545–553. [Google Scholar] [CrossRef] [Green Version]
- Feng, Q.; Liu, Y.; Huang, J.; Chen, K.; Huang, J.; Xiao, K. Uptake, distribution, clearance, and toxicity of iron oxide nanoparticles with different sizes and coatings. Sci. Rep. 2018, 8, 2082. [Google Scholar] [CrossRef]
- Denning, D.W.; Bromley, M.J. Infectious Disease. How to bolster the antifungal pipeline. Science 2015, 347, 1414–1416. [Google Scholar] [CrossRef] [Green Version]
- Perfect, J.R. “Is there an emerging need for new antifungals?”. Expert. Opin. Emerg. Drugs 2016, 21, 129–131. [Google Scholar] [CrossRef] [PubMed]
- Gustafson, H.H.; Holt-Casper, D.; Grainger, D.W.; Ghandehari, H. Nanoparticle uptake: The phagocyte problem. Nano Today 2015, 10, 487–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bandeira, A.C.; de Oliveira Matos, A.; Evangelista, B.S.; da Silva, S.M.; Nagib, P.R.A.; de Moraes Crespo, A.; Amaral, A.C. Is it possible to track intracellular chitosan nanoparticles using magnetic nanoparticles as contrast agent? Bioorg. Med. Chem. 2019, 27, 2637–2643. [Google Scholar] [CrossRef] [PubMed]
- Puccia, R.; Schenkman, S.; Gorin, P.A.J.; Travassos, L.R. Exocellular components of Paracoccidoides brasiliensis: Identification of a specific antigen. Infect. Immun. 1986, 53, 199–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Nanostructures | Encapsulated Compound | Reference |
|---|---|---|
| PLGA + DMSA | Amphotericin B | Amaral et al. [191] |
| PLGA + DMSA | P10 peptide | Amaral et al. [187] |
| Liposome and PLGA | DNA Hsp65 vaccine | Ribeiro et al. [189] |
| Magnetic nanoparticles | Amphotericin B | Saldanha et al. [192] |
| PLGA | scFv | Jannuzzi et al. [190] |
| PLGA + DMSA | Itraconazole | Cunha-Azevedo et al. [193] |
| Chitosan | P10 peptide | Rodrigues et al. [188] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seki Kioshima, E.; de Souza Bonfim de Mendonça, P.; de Melo Teixeira, M.; Grenier Capoci, I.R.; Amaral, A.; Vilugron Rodrigues-Vendramini, F.A.; Lauton Simões, B.; Rodrigues Abadio, A.K.; Fernandes Matos, L.; Soares Felipe, M.S. One Century of Study: What We Learned about Paracoccidioides and How This Pathogen Contributed to Advances in Antifungal Therapy. J. Fungi 2021, 7, 106. https://doi.org/10.3390/jof7020106
Seki Kioshima E, de Souza Bonfim de Mendonça P, de Melo Teixeira M, Grenier Capoci IR, Amaral A, Vilugron Rodrigues-Vendramini FA, Lauton Simões B, Rodrigues Abadio AK, Fernandes Matos L, Soares Felipe MS. One Century of Study: What We Learned about Paracoccidioides and How This Pathogen Contributed to Advances in Antifungal Therapy. Journal of Fungi. 2021; 7(2):106. https://doi.org/10.3390/jof7020106
Chicago/Turabian StyleSeki Kioshima, Erika, Patrícia de Souza Bonfim de Mendonça, Marcus de Melo Teixeira, Isis Regina Grenier Capoci, André Amaral, Franciele Abigail Vilugron Rodrigues-Vendramini, Bruna Lauton Simões, Ana Karina Rodrigues Abadio, Larissa Fernandes Matos, and Maria Sueli Soares Felipe. 2021. "One Century of Study: What We Learned about Paracoccidioides and How This Pathogen Contributed to Advances in Antifungal Therapy" Journal of Fungi 7, no. 2: 106. https://doi.org/10.3390/jof7020106
APA StyleSeki Kioshima, E., de Souza Bonfim de Mendonça, P., de Melo Teixeira, M., Grenier Capoci, I. R., Amaral, A., Vilugron Rodrigues-Vendramini, F. A., Lauton Simões, B., Rodrigues Abadio, A. K., Fernandes Matos, L., & Soares Felipe, M. S. (2021). One Century of Study: What We Learned about Paracoccidioides and How This Pathogen Contributed to Advances in Antifungal Therapy. Journal of Fungi, 7(2), 106. https://doi.org/10.3390/jof7020106