
Review of T-2307, an Investigational Agent That Causes Collapse of Fungal Mitochondrial Membrane Potential


Abstract
:1. Introduction
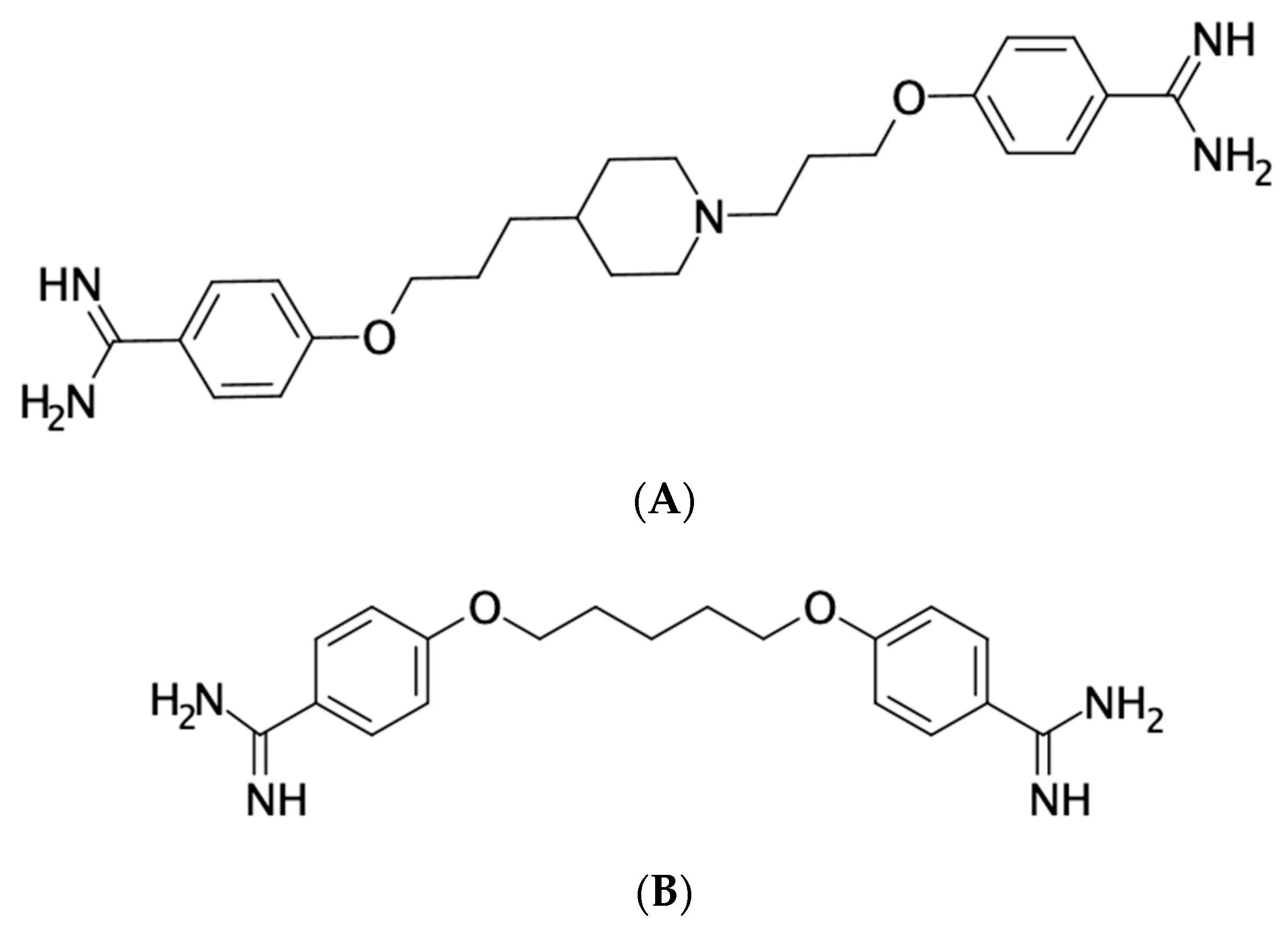
2. Structure and Mechanism of Action
3. In Vitro Spectrum of Activity
4. In Vivo Effectiveness
5. Tolerability, Safety, and Clinical Data
6. Conclusions
Funding
Conflicts of Interest
References
- WHO. Antimicrobial Resistance: Global Report on Surveillance. Available online: http://www.who.int/drugresistance/documents/surveillancereport/en/ (accessed on 21 June 2020).
- Alexander, B.D.; Johnson, M.D.; Pfeiffer, C.D.; Jiménez-Ortigosa, C.; Catania, J.; Booker, R.; Castanheira, M.; Messer, S.A.; Perlin, D.S.; Pfaller, M.A. Increasing Echinocandin Resistance in Candida glabrata: Clinical Failure Correlates With Presence of FKS Mutations and Elevated Minimum Inhibitory Concentrations. Clin. Infect. Dis. 2013, 56, 1724–1732. [Google Scholar] [CrossRef] [Green Version]
- Vallabhaneni, S.; Cleveland, A.A.; Farley, M.M.; Harrison, L.H.; Schaffner, W.; Beldavs, Z.G.; Derado, G.; Pham, C.D.; Lockhart, S.R.; Smith, R.M. Epidemiology and Risk Factors for Echinocandin Nonsusceptible Candida glabrata Bloodstream Infections: Data From a Large Multisite Population-Based Candidemia Surveillance Program, 2008–2014. Open Forum Infect. Dis. 2015, 2, ofv163. [Google Scholar] [CrossRef]
- Pfaller, M.A.; Castanheira, M.T.D.; Lockhart, S.R.; Ahlquist, A.M.; Messer, S.A.; Jones, R.N. Frequency of Decreased Susceptibility and Resistance to Echinocandins among Fluconazole-Resistant Bloodstream Isolates of Candida glabrata. J. Clin. Microbiol. 2012, 50, 1199–1203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lockhart, S.R.; Etienne, K.A.; Vallabhaneni, S.; Farooqi, J.; Chowdhary, A.; Govender, N.P.; Colombo, A.L.; Calvo, B.; Cuomo, C.A.; Desjardins, C.A.; et al. Simultaneous Emergence of Multidrug-Resistant Candida auris on 3 Continents Confirmed by Whole-Genome Sequencing and Epidemiological Analyses. Clin. Infect. Dis. 2017, 64, 134–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chow, N.A.; Gade, L.; Tsay, S.V.; Forsberg, K.; Greenko, J.A.; Southwick, K.L.; Barrett, P.M.; Kerins, J.L.; Lockhart, S.R.; Chiller, T.M.; et al. Multiple introductions and subsequent transmission of multidrug-resistant Candida auris in the USA: A molecular epidemiological survey. Lancet Infect. Dis. 2018, 18, 1377–1384. [Google Scholar] [CrossRef]
- Chowdhary, A.; Sharma, C.; Meis, J.F. Candida auris: A rapidly emerging cause of hospital-acquired multidrug-resistant fungal infections globally. PLOS Pathog. 2017, 13, e1006290. [Google Scholar] [CrossRef]
- Bruüggemann, R.J.M.; Alffenaar, J.C.; Blijlevens, N.M.A.; Billaud, E.M.; Kosterink, J.G.W.; Verweij, P.E.; Burger, D.M. Clinical Relevance of the Pharmacokinetic Interactions of Azole Antifungal Drugs with Other Coadministered Agents. Clin. Infect. Dis. 2009, 48, 1441–1458. [Google Scholar] [CrossRef] [Green Version]
- Lewis, R.E. Current Concepts in Antifungal Pharmacology. Mayo Clin. Proc. 2011, 86, 805–817. [Google Scholar] [CrossRef] [Green Version]
- Kontoyiannis, D.P.; Lewis, R.E. Treatment Principles for the Management of Mold Infections. Cold Spring Harb. Perspect. Med. 2015, 5, a019737. [Google Scholar] [CrossRef] [Green Version]
- Klatt, M.E.; Eschenauer, G.A. Review of Pharmacologic Considerations in the Use of Azole Antifungals in Lung Transplant Recipients. J. Fungi 2021, 7, 76. [Google Scholar] [CrossRef] [PubMed]
- Mitsuyama, J.; Nomura, N.; Hashimoto, K.; Yamada, E.; Nishikawa, H.; Kaeriyama, M.; Kimura, A.; Todo, Y.; Narita, H. In Vitro and In Vivo Antifungal Activities of T-2307, a Novel Arylamidine. Antimicrob. Agents Chemother. 2008, 52, 1318–1324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shibata, T.; Takahashi, T.; Yamada, E.; Kimura, A.; Nishikawa, H.; Hayakawa, H.; Nomura, N.; Mitsuyama, J. T-2307 Causes Collapse of Mitochondrial Membrane Potential in Yeast. Antimicrob. Agents Chemother. 2012, 56, 5892–5897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, E.; Nishikawa, H.; Nomura, N.; Mitsuyama, J. T-2307 Shows Efficacy in a Murine Model of Candida glabrata Infection despite In Vitro Trailing Growth Phenomena. Antimicrob. Agents Chemother. 2010, 54, 3630–3634. [Google Scholar] [CrossRef] [Green Version]
- Lanteri, C.A.; Trumpower, B.L.; Tidwell, R.R.; Meshnick, S.R. DB75, a Novel Trypanocidal Agent, Disrupts Mitochondrial Function in Saccharomyces cerevisiae. Antimicrob. Agents Chemother. 2004, 48, 3968–3974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishikawa, H.; Yamada, E.; Shibata, T.; Uchihashi, S.; Fan, H.; Hayakawa, H.; Nomura, N.; Mitsuyama, J. Uptake of T-2307, a novel arylamidine, in Candida albicans. J. Antimicrob. Chemother. 2010, 65, 1681–1687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishikawa, H.; Sakagami, T.; Yamada, E.; Fukuda, Y.; Hayakawa, H.; Nomura, N.; Mitsuyama, J.; Miyazaki, T.; Mukae, H.; Kohno, S. T-2307, a novel arylamidine, is transported into Candida albicans by a high-affinity spermine and spermidine carrier regulated by Agp2. J. Antimicrob. Chemother. 2016, 71, 1845–1855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toninello, A.; Via, L.D.; Siliprandi, D.; Garlid, K. Evidence that spermine, spermidine, and putrescine are transported electrophoretically in mitochondria by a specific polyamine uniporter. J. Biol. Chem. 1992, 267, 18393–18397. [Google Scholar] [CrossRef]
- Yamashita, K.; Miyazaki, T.; Fukuda, Y.; Mitsuyama, J.; Saijo, T.; Shimamura, S.; Yamamoto, K.; Imamura, Y.; Izumikawa, K.; Yanagihara, K.; et al. The Novel Arylamidine T-2307 Selectively Disrupts Yeast Mitochondrial Function by Inhibiting Respiratory Chain Complexes. Antimicrob. Agents Chemother. 2019, 63, e00374-19. [Google Scholar] [CrossRef] [Green Version]
- Wiederhold, N.P.; Najvar, L.K.; Fothergill, A.W.; Bocanegra, R.; Olivo, M.; McCarthy, D.I.; Fukuda, Y.; Mitsuyama, J.; Patterson, T.F. The novel arylamidine T-2307 demonstrates in vitro and in vivo activity against echinocandin-resistant Candida glabrata. J. Antimicrob. Chemother. 2015, 71, 692–695. [Google Scholar] [CrossRef] [Green Version]
- Wiederhold, N.P.; Najvar, L.K.; Fothergill, A.W.; Bocanegra, R.; Olivo, M.; McCarthy, D.I.; Kirkpatrick, W.R.; Fukuda, Y.; Mitsuyama, J.; Patterson, T.F. The Novel Arylamidine T-2307 MaintainsIn Vitroand In Vivo Activity against Echinocandin-Resistant Candida albicans. Antimicrob. Agents Chemother. 2014, 59, 1341–1343. [Google Scholar] [CrossRef] [Green Version]
- Wiederhold, N.P.; Najvar, L.K.; Jaramillo, R.; Olivo, M.; Patterson, H.; Connell, A.; Fukuda, Y.; Mitsuyama, J.; Catano, G.; Patterson, T.F. The Novel Arylamidine T-2307 Demonstrates In Vitro and In Vivo Activity against Candida auris. Antimicrob. Agents Chemother. 2019, 64. [Google Scholar] [CrossRef]
- Nishikawa, H.; Fukuda, Y.; Mitsuyama, J.; Tashiro, M.; Tanaka, A.; Takazono, T.; Saijo, T.; Yamamoto, K.; Nakamura, S.; Imamura, Y.; et al. In vitro and in vivo antifungal activities of T-2307, a novel arylamidine, against Cryptococcus gattii: An emerging fungal pathogen. J. Antimicrob. Chemother. 2017, 72, 1709–1713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Filamentous Fungi, Document M38, 3rd ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017. [Google Scholar]
- CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts, Document M27, 4th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017. [Google Scholar]
- Arendrup, M.C.; Cuenca-Estrella, M.; Lass-Flörl, C.; Hope, W. EUCAST technical note on the EUCAST definitive document EDef 7.2: Method for the determination of broth dilution minimum inhibitory concentrations of antifungal agents for yeasts EDef 7.2 (EUCAST-AFST)*. Clin. Microbiol. Infect. 2012, 18, E246–E247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiederhold, N.P.; Najvar, L.; Bocanegra, R.; Kirkpatrick, W.; Patterson, T. Comparison of anidulafungin’s and fluconazole’s in vivo activity in neutropenic and non-neutropenic models of invasive candidiasis. Clin. Microbiol. Infect. 2012, 18, E20–E23. [Google Scholar] [CrossRef] [Green Version]
- Abe, M.; Nakamura, S.; Kinjo, Y.; Masuyama, Y.; Mitsuyama, J.; Kaku, M.; Miyazaki, Y. Efficacy of T-2307, a novel arylamidine, against ocular complications of disseminated candidiasis in mice. J. Antimicrob. Chemother. 2019, 74, 1327–1332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coyle, P.; Carr, A.D.; Depczynski, B.B.; Chisholm, D.J. Diabetes mellitus associated with pentamidine use in HIV-infected patients. Med. J. Aust. 1996, 165, 587–588. [Google Scholar] [CrossRef] [PubMed]
- Lionakis, M.S.; Chamilos, G.; Lewis, R.E.; Wiederhold, N.P.; Raad, I.I.; Samonis, G.; Kontoyiannis, D.P. Pentamidine Is Active in a Neutropenic Murine Model of Acute Invasive Pulmonary Fusariosis. Antimicrob. Agents Chemother. 2006, 50, 294–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Brien, J.G.; Dong, B.J.; Coleman, R.L.; Gee, L.; Balano, K.B. A 5-Year Retrospective Review of Adverse Drug Reactions and Their Risk Factors in Human Immunodeficiency Virus—Infected Patients Who Were Receiving Intravenous Pentamidine Therapy for Pneumocystis carinii Pneumonia. Clin. Infect. Dis. 1997, 24, 854–859. [Google Scholar] [CrossRef]
- Helmick, C.G.; Green, J.K. Pentamidine-Associated Hypotension and Route of Administration. Ann. Intern. Med. 1985, 103, 480. [Google Scholar] [CrossRef]


{kind=link}
| Mechanism of Action | Causes the collapse of the mitochondrial membrane potential by targeting respiratory chain enzymatic complexes III and IV in the inner mitochondrial membrane of yeasts. |
| Spectrum of Activity | Candida species, including azole- and echinocandin-resistant isolates, Cryptococcus neoformans & C. gattii. Variable activity against Aspergillus species. Limited in vitro data against fungi suggests limited to no activity against Fusarium, Scedosporium, and the Mucorales. |
| In vivo activity | Efficacy in murine models of invasive candidiasis caused by C. albicans, C. glabrata, and C. auris; reductions in the ocular fungal burden caused by C. albicans. Efficacy in murine models of systemic aspergillosis due to Aspergillus fumigatus and cryptococcosis caused by C. neoformans and C. gattii. |
| Pharmacokinetic and clinical data | Not currently available in the literature. |
| Species | MIC Range | MIC50 | MIC90 |
|---|---|---|---|
| Candida albicans | 0.00025–0.008 | ≤0.008 | ≤0.008 |
| Candida auris | ≤0.008–0.015 | 0.015 | 0.015 |
| Candida glabrata | 0.004–>4 | ≤0.008 | ≤0.008 |
| Candida guilliermondii | 0.001–0.004 | 0.002 | 0.004 |
| Candida krusei | 0.0005–0.002 | 0.001 | 0.002 |
| Candida tropicalis | 0.00025–0.0005 | 0.0005 | 0.0005 |
| Cryptococcus neoformans | 0.0039–0.06 | 0.015 | 0.03 |
| Cryptococcus gattii | 0.008–0.06 | 0.03 | 0.06 |
| Aspergillus fumigatus | 0.125–4 | 1 | 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wiederhold, N.P. Review of T-2307, an Investigational Agent That Causes Collapse of Fungal Mitochondrial Membrane Potential. J. Fungi 2021, 7, 130. https://doi.org/10.3390/jof7020130
Wiederhold NP. Review of T-2307, an Investigational Agent That Causes Collapse of Fungal Mitochondrial Membrane Potential. Journal of Fungi. 2021; 7(2):130. https://doi.org/10.3390/jof7020130
Chicago/Turabian StyleWiederhold, Nathan P. 2021. "Review of T-2307, an Investigational Agent That Causes Collapse of Fungal Mitochondrial Membrane Potential" Journal of Fungi 7, no. 2: 130. https://doi.org/10.3390/jof7020130
APA StyleWiederhold, N. P. (2021). Review of T-2307, an Investigational Agent That Causes Collapse of Fungal Mitochondrial Membrane Potential. Journal of Fungi, 7(2), 130. https://doi.org/10.3390/jof7020130