Root Endophytic Fungal Community and Carbon and Nitrogen Stable Isotope Patterns Differ among Bletilla Species (Orchidaceae)

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Plant Species
2.2. Sampling
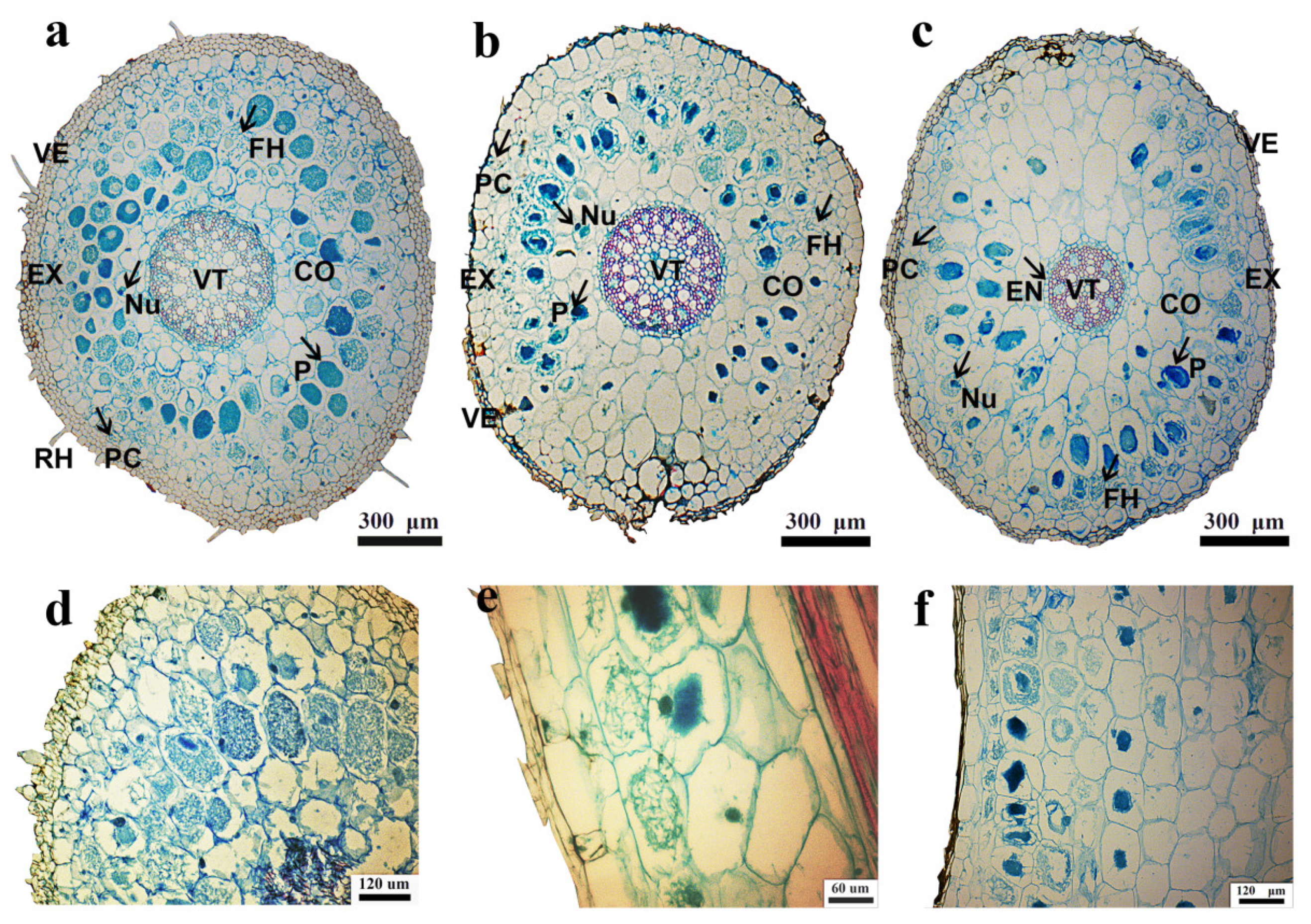
2.3. Observation of Mycelium Colonization
2.4. Molecular Identification of Root Endophytic Fungi
2.5. Analysis of Stable Isotope Abundance and N Concentration
2.6. Statistical Analyses
3. Results
3.1. Mycelium Colonization of Three Bletilla Species
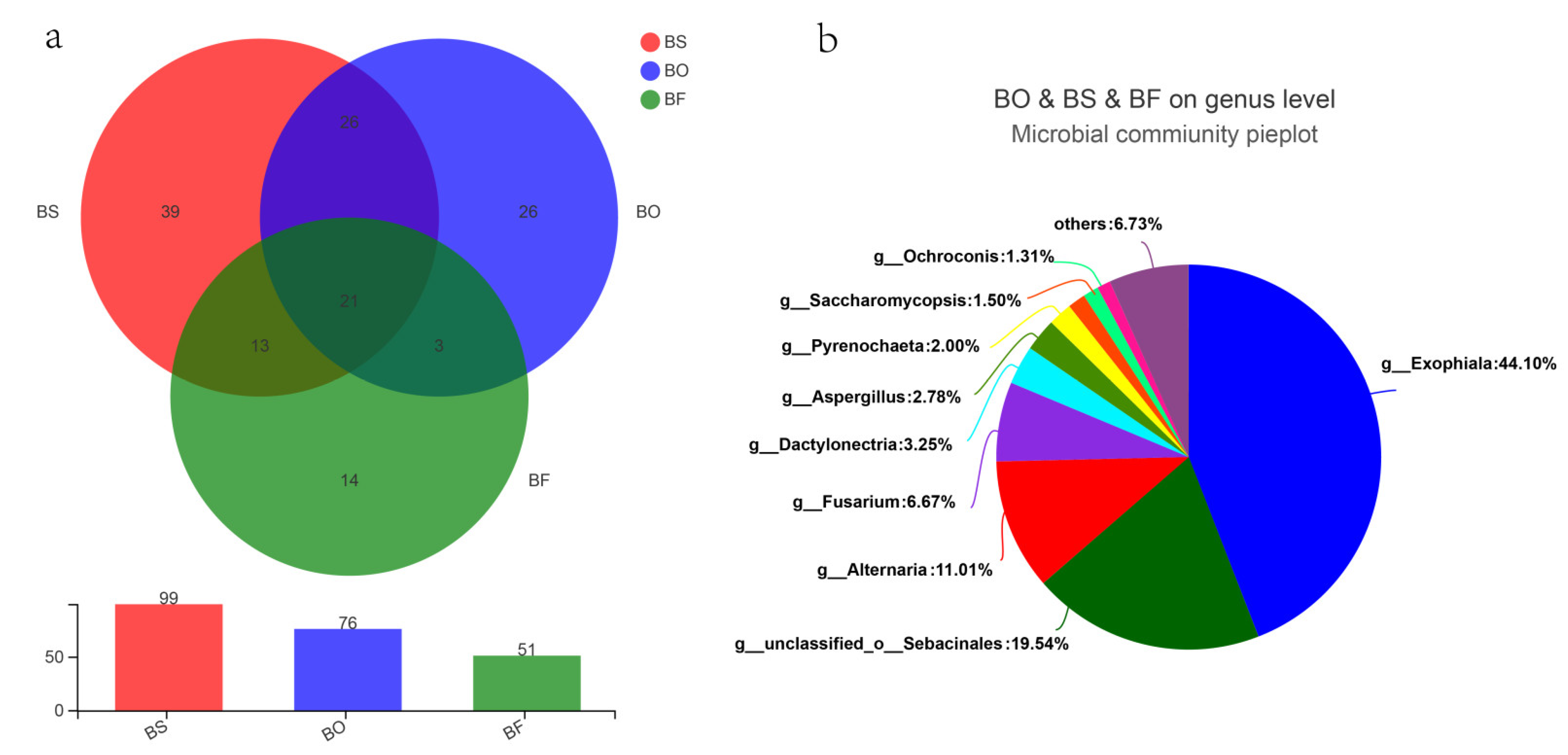
3.2. Fungal Community Found in Bletilla Roots
3.3. Diversity of Root Endophytic Fungi for Three Bletilla Species
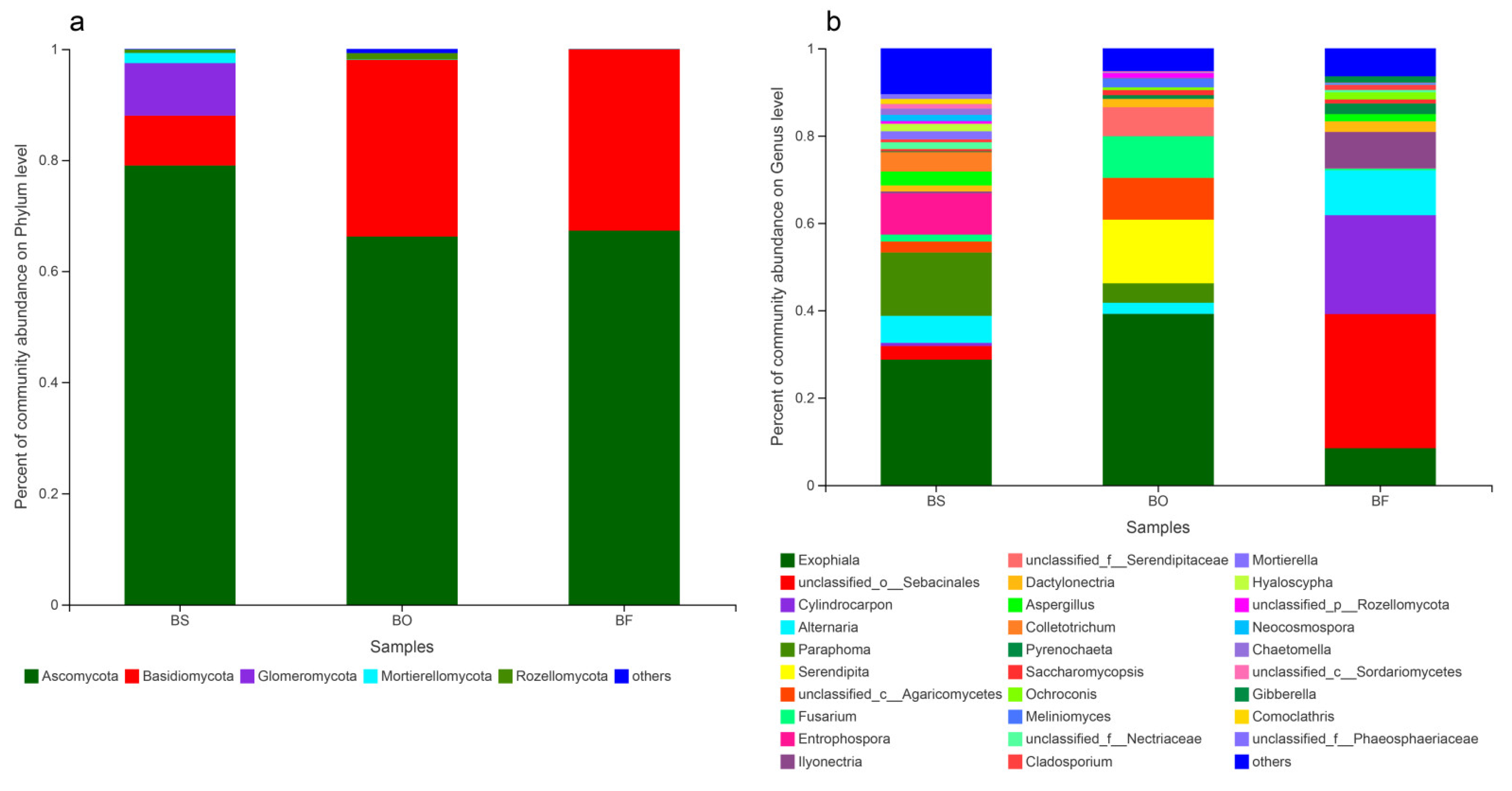
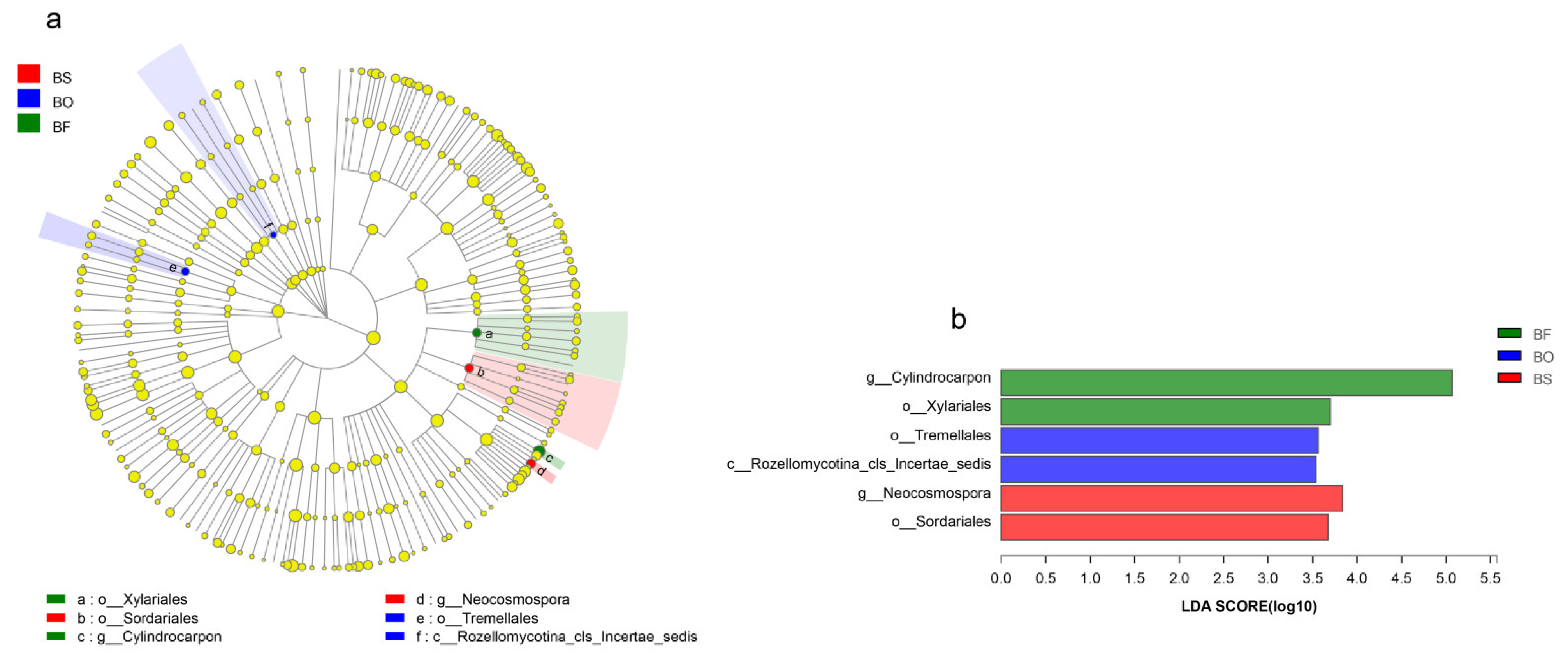
3.4. Community Composition of Root Endophytic Fungi among Bletilla Species
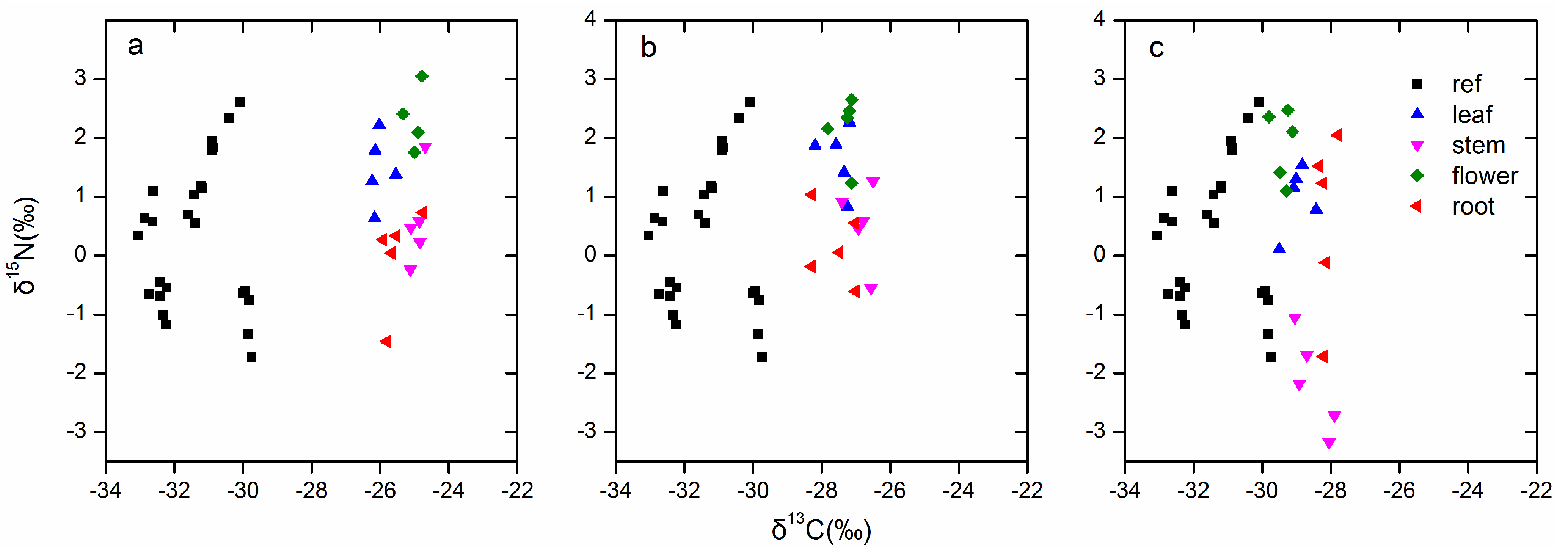
3.5. Stable Isotope Natural Abundance of Different Organs for Three Bletilla Species
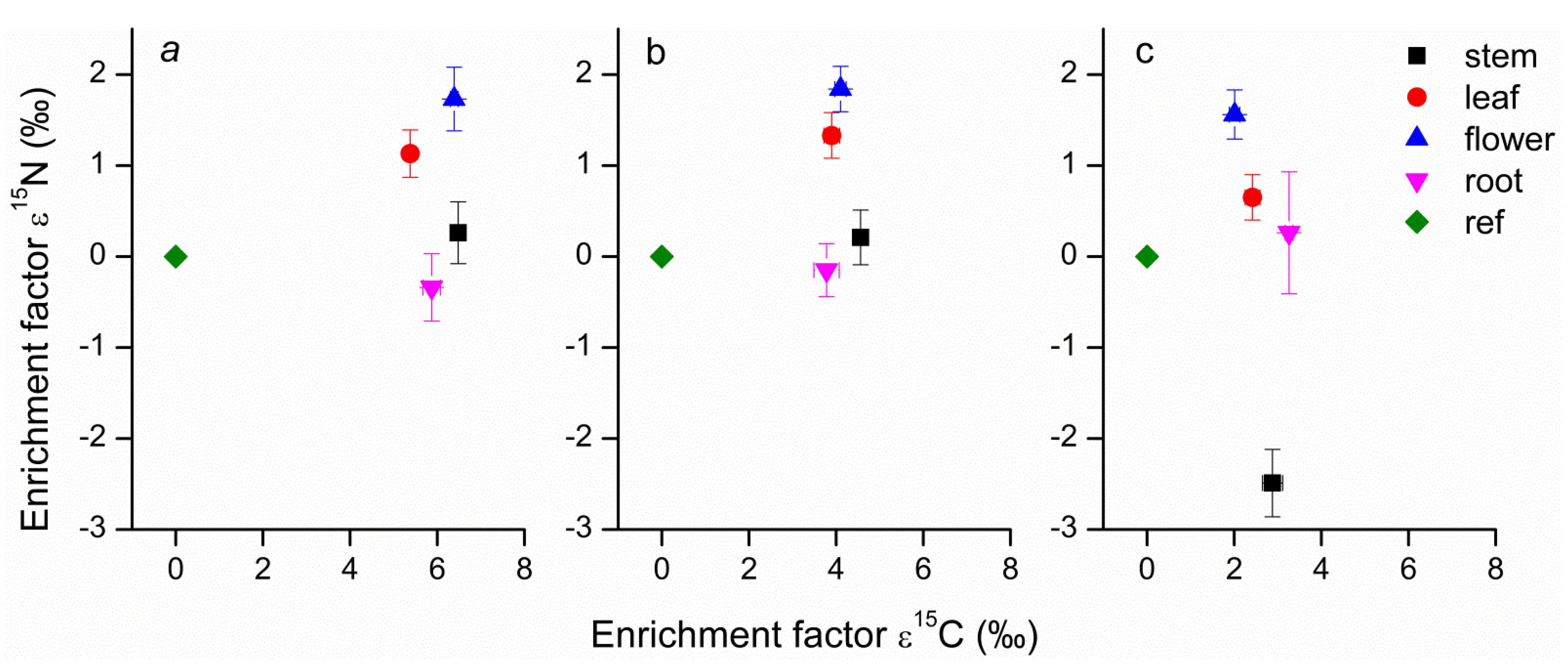
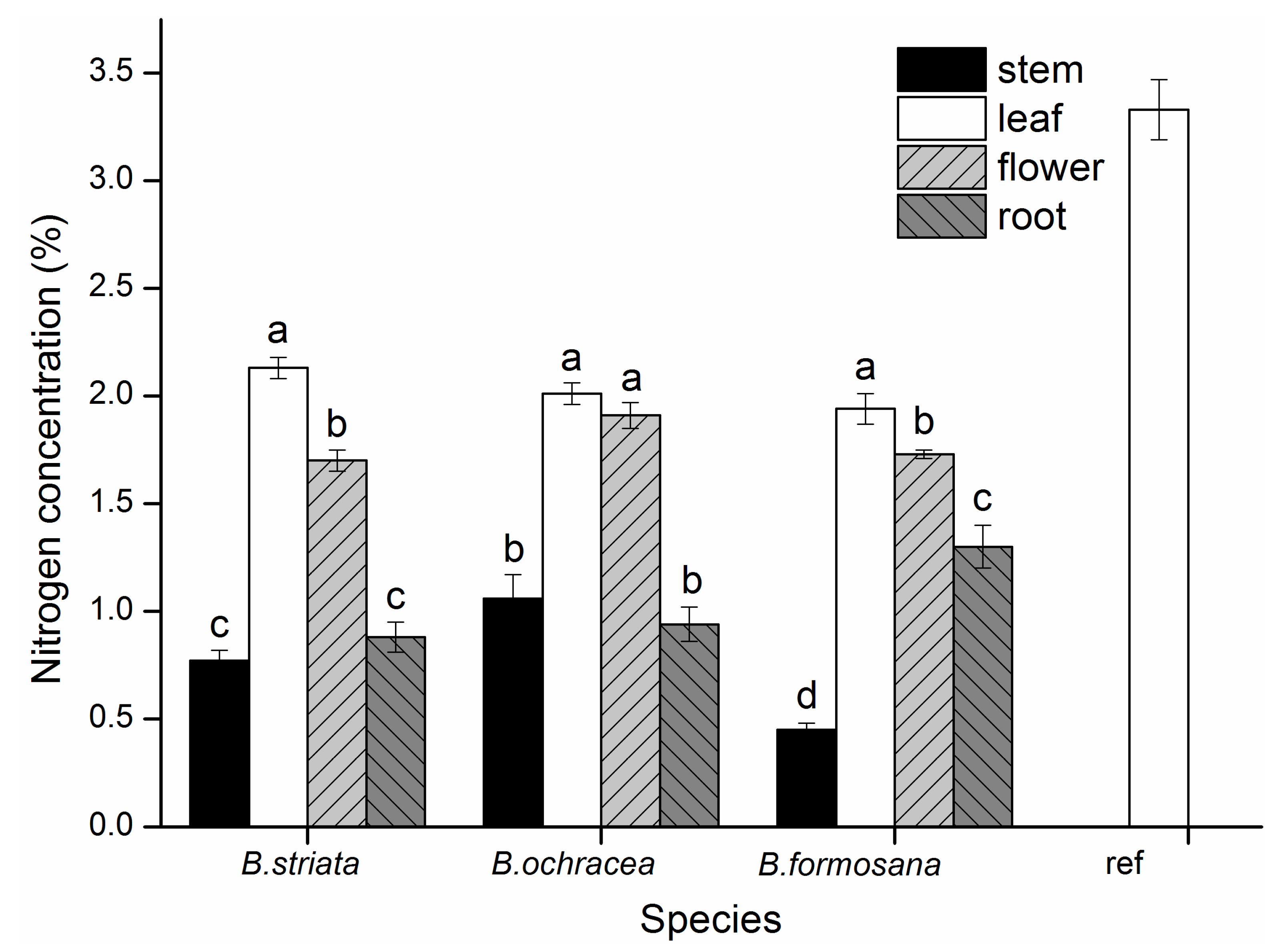
3.6. Enrichment Factors and N Concentrations among Different Species and Different Organs of Bletilla
4. Discussion
4.1. Composition and Diversity of Root Endophytic Fungi
4.2. Isotope Signature and Nutrient Gain from Fungal Partners
4.3. The Functional Roles of Root-Associated Fungal Endophytes
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ogura-Tsujita, Y.; Gebauer, G.; Xu, H.; Fukasawa, Y.; Umata, H.; Tetsuka, K.; Kubota, M.; Schweiger, J.M.I.; Yamashita, S.; Maekawa, N. The giant mycoheterotrophic orchid Erythrorchis altissima is associated mainly with a divergent set of wood-decaying fungi. Mol. Ecol. 2018, 27, 1324–1337. [Google Scholar] [CrossRef] [PubMed]
- Arditti, J.; Ghani, A.K.A. Numerical and physical properties of orchid seeds and their biological implications. New Phytol. 2002, 145, 367–421. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: New York, NY, USA, 2008. [Google Scholar]
- Schiebold, J.M.I.; Bidartondo, M.I.; Karasch, P.; Gravendeel, B.; Gebauer, G. You are what you get from your fungi: Nitrogen stable isotope patterns in Epipactis species. Ann. Bot. 2017, 119, 1085–1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, S.E. Physiology and ecology of orchid mycorrhizal fungi with reference to seedling nutrition. New Phytol. 1967, 66, 371–378. [Google Scholar] [CrossRef]
- Yan, R. Taxonomy of mycorrhizal fungi of orchid plants and symbiosis nutritional relationship between fungi and plant. J. West China For. Sci. 2004, 33, 50–53. (In Chinese) [Google Scholar]
- Chen, Y.H.; Xing, X.K.; Guo, S.X. Nutritional relationships between orchids and mycorrhizal fungi: A review. Mycosystema 2017, 36, 807–819. (In Chinese) [Google Scholar]
- Liu, S.S.; Chen, J.; Li, S.C.; Zeng, X.; Meng, Z.X.; Guo, S.X. Comparative transcriptome analysis of genes involved in GA-GID1-DELLA regulatory module in symbiotic and asymbiotic seed germination of Anoectochilus roxburghii (Wall.) Lindl. (Orchidaceae). Int. J. Mol. Sci. 2015, 16, 30190–30203. [Google Scholar] [CrossRef] [Green Version]
- Herrera, H.; Valadares, R.; Oliveira, G.; Fuentes, A.; Almonacis, L.; Bashan, Y.; Arriagada, C. Adaptation and tolerance mechanisms developed by mycorrhizal Bipinnula fimbriata plantlets (Orchidaceae) in a heavy metal-polluted ecosystem. Mycorrhiza 2018, 28, 651–663. [Google Scholar] [CrossRef]
- Gebauer, G.; Meyer, M. 15N and 13C natural abundance of autotrophic and mycoheterotrophic orchids provides insight into nitrogen and carbon gain from fungal association. New Phytol. 2003, 160, 209–223. [Google Scholar] [CrossRef]
- Gonneau, C.; Jersakova, J.; de Tredern, E.; Till-Bottraud, I.; Saarinen, K.; Sauve, M.; Roy, M.; Hajek, T.; Selosse, M.A. Photosynthesis in perennial mixotrophic Epipactis spp. (Orchidaceae) contributes more to shoot and fruit biomass than to hypogeous survival. J. Ecol. 2014, 102, 1183–1194. [Google Scholar] [CrossRef]
- Liebel, H.T.; Bidartondo, M.I.; Gebauer, G. Are carbon and nitrogen exchange between fungi and the orchid Goodyera repens affected by irradiance? Ann. Bot. 2015, 115, 251–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burke, R.M.; Cairney, J.W.G. Carbohydrate oxidases in ericoid and ectomycorrhizal fungi: A possible source of Fenton radicals during the degradation of lignocelluloses. New Photol. 1998, 139, 637–645. [Google Scholar] [CrossRef]
- Slezack, S.; Dumas-Gaudot, E.; Rosendahl, S.; Kjoller, R. Endoproteolytic activities in pea roots inoculated with the arbuscular mycorrhizal fungus Glomus mosseae and/or Aphanomyces euteiches in relation to bioprotection. New Photol. 1999, 142, 517–529. [Google Scholar] [CrossRef]
- Stockel, M.; Tesitelova, T.; Jersakova, J.; Bidartondo, M.I.; Gebauer, G. Carbon and nitrogen gain during the growth of orchid seedlings in nature. New Phytol. 2014, 202, 606–615. [Google Scholar] [CrossRef] [PubMed]
- Merckx, V.S.F.T. Mycoheterotrophy: An introduction. In Mycoheterotrophy: The Biology of Plants Living on Fungi; Merckx, V., Ed.; Springer: New York, NY, USA, 2013; pp. 1–17. [Google Scholar]
- Roy, M.; Gonneau, C.; Rocheteau, A.; Berveiller, D.; Thomas, J.C.; Damesin, C.; Selosse, M.A. Why do mixotrophic plants stay green? A comparison between green and achlorophyllous orchid individuals in situ. Ecol. Monogr. 2013, 83, 95–117. [Google Scholar] [CrossRef] [Green Version]
- Preiss, K.; Adam, I.K.U.; Gebauer, G. Irradiance governs exploitation of fungi: Fine-tuning of carbon gain by two partially myco-heterotrophic orchids. Proc. R. Soc. B Biol. Sci. 2010, 277, 1333–1336. [Google Scholar] [CrossRef] [Green Version]
- Bellino, A.; Alfani, A.; Selosse, M.A.; Guerrieri, R.; Borghetti, M.; Baldantoni, D. Nutritional regulation in mixotrophic plants: New insights from Limodorum abortivum. Oecologia 2014, 175, 875–885. [Google Scholar] [CrossRef]
- Hynson, N.A.; Madsen, T.P.; Selosse, M.A.; Adam, I.K.U.; Ogura-Tsujita, Y.; Roy, M.; Gebauer, G. The physiological ecology of mycoheterotrophy. In Mycoheterotrophy. The Biology of Plants Living on Fungi; Merckx, V.S.F.T., Ed.; Springer: New York, NY, USA, 2013; pp. 297–342. [Google Scholar]
- Dawson, T.E.; Mambelli, S.; Plamboeck, A.H.; Templer, P.H.; Tu, K.P. Stable isotopes in plant ecology. Annu. Rev. Ecol. Syst. 2002, 33, 507–559. [Google Scholar] [CrossRef]
- Ercole, E.; Adamo, M.; Rodda, M.; Gebauer, G.; Girlanda, M.; Perotto, S. Temporal variation in mycorrhizal diversity and carbon and nitrogen stable isotope abundance in the wintergreen meadow orchid Anacamptis morio. New Phytol. 2015, 205, 1308–1319. [Google Scholar] [CrossRef]
- Gebauer, G.; Dietrich, P. Nitrogen isotope ratios in different compartments of a mixed stand of spruce, larch and beech trees and of understory vegetation including fungi. Isot. Isot. Environ. Health Stud. 1993, 29, 35–44. [Google Scholar] [CrossRef]
- Gleixner, G.; Danier, H.J.; Werner, R.A.; Schmidt, H.L. Correlations between the 13C content of primary and secondary plant products in different cell compartments and that in decomposing basidiomycetes. Plant Physiol. 1993, 102, 1287–1290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, D.L.; Bruns, T.D.; Szaro, T.M.; Hodges, S.A. Divergence in mycorrhizal specialization within Hexalectris spicata (Orchidaceae), a nonphotosynthetic desert orchid. Am. J. Bot. 2003, 90, 1168–1179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trudell, S.A.; Rygiewicz, P.T.; Edmonds, R.L. Nitrogen and carbon stable isotope abundances support the myco-heterotrophic nature and host-specificity of certain achlorophyllous plants. New Phytol. 2003, 160, 391–401. [Google Scholar] [CrossRef] [Green Version]
- Roy, M.; Watthana, S.; Richard, F.; Vessabutr, S.; Selosse, M.A. Mycoheterotrophic orchids from Thailand tropical dipterocarpacean forests associate with a broad diversity of ectomycorrhizal fungi. BMC Biol. 2009, 7, 51. [Google Scholar] [CrossRef] [Green Version]
- Yagame, T.; Orihara, T.; Selosse, M.; Yamato, M.; Iwase, K. Mixotrophy of Platanthera minor, an orchid associated with ectomycorrhiza-forming Ceratobasidiaceae fungi. New Phytol. 2012, 193, 178–187. [Google Scholar] [CrossRef]
- Martos, F.; Dulormne, M.; Pailler, T.; Bonfante, P.; Faccio, A.; Fournel, J.; Dubois, M.P.; Selosse, M.A. Independent recruitment of saprotrophic fungi as mycorrhizal partners by tropical achlorophyllous orchids. New Phytol. 2009, 184, 668–681. [Google Scholar] [CrossRef]
- Ogura-Tsujita, Y.; Gebauer, G.; Hashimoto, T.; Umata, H.; Yukawa, T. Evidence for novel and specialised mycorrhizal parasitism: The orchid Gastrodia confusa gains carbon from saprotrophic Mycena. Proc. R. Soc. Lond. B Biol. Sci. 2009, 276, 761–767. [Google Scholar]
- Girlanda, M.; Segreto, R.; Cafasso, D.; Liebel, H.T.; Rodda, M.; Ercole, E.; Salvatore, C.; Gebauer, G.; Perotto, S. Photosynthetic Mediterranean meadow orchids feature partial mycoheterotrophy and specific mycorrhizal associations. Am. J. Bot. 2011, 98, 1148–1163. [Google Scholar] [CrossRef] [Green Version]
- Hynson, N.A.; Bruns, T.D. Evidence of a myco-heterotroph in the plant family Ericaceae that lacksmycorrhizal specificity. Proc. R. Soc. B Biol. Sci. 2009, 276, 4053–4059. [Google Scholar] [CrossRef] [Green Version]
- Liebel, H.T.; Bidartondo, M.I.; Preiss, K.; Segreto, R.; Stockel, M.; Rodda, M.; Gebauer, G. C and N stable isotope signatures reveal constraints to nutritional modes in orchids from the Mediterranean and macaronesia. Am. J. Bot. 2010, 97, 903–912. [Google Scholar] [CrossRef]
- Flora of China Editorial Committee. Flora of China; Science Press: Beijing, China, 1999. [Google Scholar]
- Qian, C.D.; Jiang, F.S.; Yu, H.S.; Fu, Y.H.; Cheng, D.Q.; Gan, L.S.; Ding, Z.S. Antibacterial biphenanthrenes from the fibrous roots of Bletilla striata. J. Nat. Prod. 2015, 78, 939–943. [Google Scholar] [CrossRef] [PubMed]
- Li, W.P.; He, L.Y.; Ding, Z.S. Application and resources current situation of common Bletilla Tuber. Chin. Arch. Tradit. Chin. Med. 2012, 30, 158–160. (In Chinese) [Google Scholar]
- Ren, F.M.; Liu, Y.; Li, Y.; Yang, T.J.; Han, F.; Song, J.Y. Resource distribution and breeding of medicinal plants in genus Bletilla Reichb. f. Chin. Tradit. Herb. Drugs 2016, 47, 4478–4487. (In Chinese) [Google Scholar]
- Bidartondo, M.I.; Burghardt, B.; Gebauer, G.; Bruns, T.D.; Read, D.J. Changing partners in the dark: Isotopic and molecular evidence of ectomycorrhizal liaisons between forest orchids and trees. Proc. R. Soc. B Biol. Sci. 2004, 271, 1799–1806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gebauer, G.; Schulze, E.D. Carbon and nitrogen isotope ratios in different compartments of a healthy and a declining Picea abies forest in the Fichtelgebirge, NE Bavaria. Oecologia 1991, 87, 198–207. [Google Scholar] [CrossRef]
- Hynson, N.A.; Preiss, K.; Gebauer, G. Is it better to give than receive? A stable isotope perspective to orchid–fungal carbon transport in the green orchid species Goodyera repens and G. oblongifolia. New Phytol. 2009, 182, 8–11. [Google Scholar] [CrossRef]
- Qin, J.; Zhang, W.; Ge, Z.W.; Zhang, S.B. Molecular identifications uncover diverse fungal symbionts of Pleione (Orchidaceae). Fungal Ecol. 2019, 37, 19–29. [Google Scholar] [CrossRef]
- Bronstein, J.L. The evolution of facilitation and mutualism. J. Ecol. 2009, 97, 1160–1170. [Google Scholar] [CrossRef]
- Taylor, D.L.; Bruns, T.D.; Hodges, S.A. Evidence for mycorrhizal races in a cheating orchid. Proc. R. Soc. B Biol. Sci. 2004, 271, 35–43. [Google Scholar] [CrossRef] [Green Version]
- Suetsugu, K.; Yamato, M.; Miura, C.; Yamaguchi, K.; Takahashi, K.; Ida, Y.; Shigenobu, S.; Kaminaka, H. Comparison of green and albino individuals of the partially mycoheterotrophic orchid Epipactis helleborine on molecular identities of mycorrhizal fungi, nutritional modes and gene expression in mycorrhizal roots. Mol. Ecol. 2017, 26, 1652–1669. [Google Scholar] [CrossRef]
- Tao, G.; Liu, Z.Y.; Sun, B.D.; Zhu, B.D.; Zhu, Y.; Cai, L.; Liu, X.Z. Occurrence and diversity of endophytic fungi in Bletilla ochracea (Orchidaceae) in Guizhou, China. Afr. J. Microbiol. Res. 2012, 6, 2859–2868. [Google Scholar]
- Liu, Z.; Tao, G.; Liu, Z.Y.; Zhu, Y. Diversity analysis of endophytic fungi associated with Bletilla ochracea (Orchidaceae). Mycosystema 2013, 32, 812–818. (In Chinese) [Google Scholar]
- Vallius, E. Factors affecting fruit and seed production in Dactylorhiza maculata (Orchidaceae). Bot. J. Linn. Soc. 2001, 135, 89–95. [Google Scholar] [CrossRef]
- Cernusak, L.A.; Tcherkez, G.; Keitel, C.; Cornwell, W.K.; Santiago, L.S.; Knohl, A.; Barbour, M.M.; Williams, D.G.; Reich, P.B.; Ellsworth, D.S.; et al. Why are non-photosynthetic tissues generally 13C enriched compared to leaves in C3 plants? Review and synthesis of current hypotheses. Funct. Plant Biol. 2009, 36, 199–213. [Google Scholar] [CrossRef] [PubMed]
- Johansson, V.A.; Mikusinska, A.; Ekblad, A.; Eriksson, O. Partial mycoheterotrophy in Pyroleae: Nitrogen and carbon stable isotope signatures during development from seedling to adult. Oecologia 2015, 177, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Cevallos, S.; Herrera, P.; Sanchez-Rodriguez, A.; Declerck, S.; Suarez, J.P. Untangling factors that drive community composition of root associated fungal endophytes of Neotropical epiphytic orchids. Fungal Ecol. 2018, 34, 67–75. [Google Scholar] [CrossRef]
- Selosse, M.A.; Roy, M. Green plants that feed on fungi: Facts and questions about mixotrophy. Trends Plant Sci. 2009, 14, 64–70. [Google Scholar] [CrossRef]
- Oliveira, S.F.; Bocayuva, M.F.; Veloso, T.G.R.; Bazzolli, D.M.S.; da Silva, C.C.; Pereira, O.L.; Kasuya, M.C.M. Endophytic and mycorrhizal fungi associated with roots of endangered native orchids from the Atlantic Forest, Brazil. Mycorrhiza 2014, 24, 55–64. [Google Scholar] [CrossRef]
- Koyyappurath, S.; Atuahiva, T.; Le Guen, R.; Batina, H.; Gautheron, N.; Edel Hermann, V.; Peribe, J.; Jahiel, M.; Liew, E.C.Y.; Alabouvette, C.; et al. Fusarium oxysporum f. sp. radicis-vanillae is the causal agent of root and stem rot of vanilla. Plant Pathol. 2016, 6, 12–25. [Google Scholar]
- Jiang, J.W.; Zhang, K.; Cheng, S.; Nie, Q.W.; Zhou, S.X.; Chen, Q.Q.; Zhou, J.L.; Zhen, X.; Li, X.T.; Zhen, T.W.; et al. Fusarium oxysporum KB-3 from Bletilla striata: An orchid mycorrhizal fungus. Mycorrhiza 2019, 29, 531–540. [Google Scholar] [CrossRef]
- Bayman, P.; Otero, J.T. Microbial Endophytes of Orchid Roots. In Microbial Root Endophytes; Schulz, B.J.E., Boyle, C.J.C., Sieber, T.N., Eds.; Springer: Berlin/Heidelberg, Germany, 2006. [Google Scholar]
- Guo, S.X. Biology of Endophytic Fungi in Medicinal Plants; Science Press: Beijing, China, 2016. [Google Scholar]
- Farh, M.E.A.; Kim, Y.J.; Singh, P.; Yang, D.K. Cross interaction between llyonectriamors-panacis isolates infecting korean ginseng and ginseng saponins in correlation with their pathogenicity. Phytopathology 2017, 107, 561–569. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.Y.; Kang, J.C.; Nontachaiyapoom, S.; Wen, T.; Hyde, K.D. Non-mycorrhizal endophytic fungi from orchids. Curr. Sci. 2015, 109, 72–87. [Google Scholar]
- Zhang, L.C.; Chen, J.; Lv, Y.L.; Gao, C.; Guo, S.X. Mycena sp., a mycorrhizal fungus of the orchid Dendrobium oficinale. Mycol. Prog. 2012, 11, 395–401. [Google Scholar] [CrossRef]
- Abarenkov, K.; Henrik Nilsson, R.; Larsson, K.H.; Alexander, I.J.; Eberhardt, U.; Erland, S.; Hoiland, K.; Kjoller, R.; Larsson, E.; Pennanen, T.; et al. The UNITE database for molecular identification of fungi recent updates and future perspectives. New Phytol. 2010, 186, 281–285. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sobs | Shannon | Simpson | Coverage (%) | |
|---|---|---|---|---|
| B. striata | 94 ± 15 a | 2.35 ± 0.48 a | 0.26 ± 0.12 a | 99.97 a |
| B. ochracea | 73 ±9 ab | 1.76 ± 0.31 a | 0.32 ± 0.06 a | 99.98 a |
| B. formosana | 39 ± 5 b | 1.89 ± 0.45 a | 0.30 ± 0.14 a | 99.99 a |
| Autotrophic References | B. striata | B. ochracea | B. formosana | |
|---|---|---|---|---|
| δ13C | −31.40 ± 0.22 d | −25.37 ± 1.02 a | −27.32 ± 1.14 b | −28.76 ± 0.33 c |
| δ15N | 0.33 ± 0.25 a | 1.02 ± 0.24 a | 1.14 ± 0.22 a | 0.33 ± 0.39 a |
| δ13C | δ15N | |||||||
|---|---|---|---|---|---|---|---|---|
| Stem | Leaf | Flower | Root | Stem | Leaf | Flower | Root | |
| B. striata | −24.92 ± 0.08 a | −26.02 ± 0.12 c | −25.01 ± 0.09 a | −25.53 ± 0.20 b | 0.59 ± 0.35 bc | 1.46 ± 0.26 ab | 2.06 ± 0.35 a | −0.01 ± 0.37 c |
| B. ochracea | −26.84 ± 0.16 a | −27.5 ± 0.18 b | −27.3 ± 0.13 a | −27.62 ± 0.29 b | 0.54 ± 0.3 b | 1.66 ± 0.25 a | 2.17 ± 0.25 a | 0.18 ± 0.28b |
| B. formosana | −28.52 ± 0.23 ab | −28.98 ± 0.17 bc | −29.39 ± 0.12 c | −28.14 ± 0.09 a | −2.16 ± 0.37 b | 0.98 ± 0.25 a | 1.89 ± 0.27 a | 0.59 ± 0.68 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, X.; Ni, Z.; Diao, H.; Jiang, K.; Hu, C.; Shao, L.; Huang, W. Root Endophytic Fungal Community and Carbon and Nitrogen Stable Isotope Patterns Differ among Bletilla Species (Orchidaceae). J. Fungi 2021, 7, 69. https://doi.org/10.3390/jof7020069
Zeng X, Ni Z, Diao H, Jiang K, Hu C, Shao L, Huang W. Root Endophytic Fungal Community and Carbon and Nitrogen Stable Isotope Patterns Differ among Bletilla Species (Orchidaceae). Journal of Fungi. 2021; 7(2):69. https://doi.org/10.3390/jof7020069
Chicago/Turabian StyleZeng, Xinhua, Ziyi Ni, Haixin Diao, Kai Jiang, Chao Hu, Li Shao, and Weichang Huang. 2021. "Root Endophytic Fungal Community and Carbon and Nitrogen Stable Isotope Patterns Differ among Bletilla Species (Orchidaceae)" Journal of Fungi 7, no. 2: 69. https://doi.org/10.3390/jof7020069
APA StyleZeng, X., Ni, Z., Diao, H., Jiang, K., Hu, C., Shao, L., & Huang, W. (2021). Root Endophytic Fungal Community and Carbon and Nitrogen Stable Isotope Patterns Differ among Bletilla Species (Orchidaceae). Journal of Fungi, 7(2), 69. https://doi.org/10.3390/jof7020069