
Compatibility of Beauveria bassiana and a Plant Secondary Metabolite: A Novel Modeling Approach to Invade Host Defense for Effective Control of Oligonychus afrasiaticus (McGregor) on Date Palms


Abstract
:1. Introduction
2. Materials and Methods
2.1. Microbial Culture
2.2. Oligonychus afrasiaticus
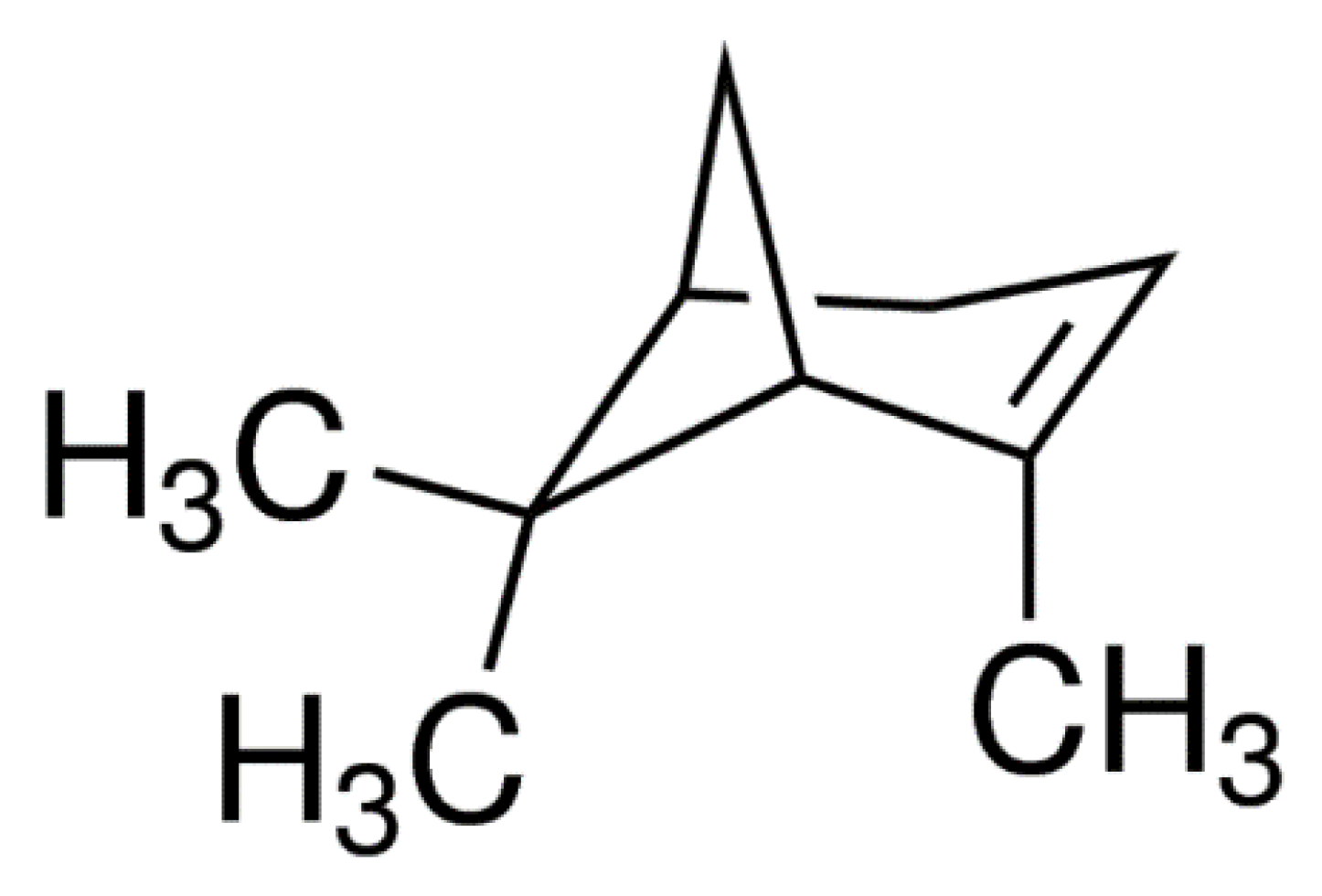
2.3. (+)-α-Pinene
2.4. Fungus-Toxin Compatibility Assays
2.4.1. Germination (GR)
2.4.2. Vegetative Growth (VG)
2.4.3. Conidiation (SP)
2.5. Fungus-Toxin Synergism Bioassays
2.6. Response of Fungus-Toxin Interactions on the Target Host Defense-Related Enzymatic Regulation
3. Results
3.1. Pathogen-Toxin Compatibility Response Analysis
3.2. Date Palm Dust Mites Mortality Response against Pathogen-Toxin Interaction
3.3. Defense-Related Enzymatic Activities Analysis
4. Discussion
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT. Food and Agricultural Commodities Production. Available online: http://www.fao.org/faostat/en/#data/QC/visualize (accessed on 21 February 2021).
- Hussain, A.; Rizwan-ul-haq, M.; AlJabr, A.M.; Al-Ayedh, H. Lethality of Sesquiterpenes Reprogramming Red Palm Weevil Detoxification Mechanism for Natural Novel Biopesticide Development. Molecules 2019, 24, 1648. [Google Scholar] [CrossRef] [Green Version]
- Hussain, A.; AlJabr, A.M.; Al-Ayedh, H. Development-Disrupting Chitin Synthesis Inhibitor, Novaluron, Reprogramming the Chitin Degradation Mechanism of Red Palm Weevils. Molecules 2019, 24, 4304. [Google Scholar] [CrossRef] [Green Version]
- Ehsine, M.; Belkadhi, M.S.; Chaieb, M. Seasonal and nocturnal activities of the Rhinoceros Borer (Coleoptera: Scarabaeidae) in the North Saharan Oases Ecosystems. J. Insect Sci. 2014, 14, 256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, A.; Rizwan-ul-haq, M.; AlJabr, A.M.; Al-Ayedh, H. Host-pathogen interaction for screening potential of Metarhizium anisopliae isolates against the date-palm dust mite, Oligonychus afrasiaticus (McGregor) (Acari: Tetranychidae). Egypt. J. Biol. Pest Control 2019, 29, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Hussain, A.; AlJabr, A.M. Potential synergy between spores of Metarhizium anisopliae and plant secondary metabolite, 1-chlorooctadecane for effective natural acaricide development. Molecules 2020, 25, 1900. [Google Scholar] [CrossRef] [Green Version]
- Coudin, R.; Galves, F. Biologie de l’acarien du palmier dattier Oligonychus afrasiaticus (MacGregor) en Mauritanie. Fruits 1976, 31, 543–550. [Google Scholar]
- Ba-Angood, S.A.; Bass’haih, G.S. A study on the effect of date palm dust mite Oligonychus afrasiaticus (McGregor) (Acarina: Tetranychidae) on the physiochemical characters of three different date varieties in Wadi Hadhramout, Yemen. Arab J. Plant Prot. 2000, 18, 82–85. [Google Scholar]
- Elwan, A.A. Survey of the insect and mite pests associated with date palm trees in Al-Dakhliya region, Sultanate of Oman. Egypt. J. Agric. Res. 2000, 78, 653–664. [Google Scholar]
- Palevsky, E.; Ucko, O.; Peles, S.; Yablonski, S.; Gerson, U. Evaluation of control measures for Oligonychus afrasiaticus infesting date palm cultivars in the Southern Arava Valley of Israel. Crop. Prot. 2004, 23, 387–392. [Google Scholar] [CrossRef]
- Ben Chaabane, S.; Chermiti, B. Characteristics of date fruit and its influence on population dynamics of Oligonychus afrasiaticus in the southern of Tunisia. Acarologia 2009, 49, 29–37. [Google Scholar]
- Chaaban, S.B.; Brahim, C.; Serge, K. Effects of host plants on distribution, abundance, developmental time and life table parameters of Oligonychus afrasiaticus (McGregor) (Acari: Tetranychidae). Papéis Avulsos Zool. 2012, 52, 121–133. [Google Scholar] [CrossRef] [Green Version]
- Yousof, D.E.; Mahmoud, M.E.E. Distribution of date palm dust mite Oligonychus afrasiaticus Meg., (Acari: Tetranychidae) in northern state in Sudan and its impact on productivity of fruits of date. Persian Gulf Crop Prot. 2013, 2, 54–59. [Google Scholar]
- Latifian, M. Integrated Pest Management of Date Palm Fruit Pests: A Review. J. Entomol. 2017, 14, 112–121. [Google Scholar] [CrossRef]
- Edongali, E.; Kerra, H.; Gashira, B. Distribution and control of date mite (Oligonychus afrasiaticus) McGregor in Libya. Arab Near East Plant Prot. Newsl. 1988, 7, 1–25. [Google Scholar]
- Talhouk, A.S. On the management of the date palm and its arthropod enemies in the Arabian Peninsula. J. Appl. Entomol. 1991, 111, 514–520. [Google Scholar] [CrossRef]
- Al-Doghairi, M.A. Effect of eight acaricides against the Date Dust Mite, Oligonychus afrasiaticus (Mcgregor) (Acari: Tetranychidae). Pak. J. Biol. Sci. 2004, 7, 1168–1171. [Google Scholar] [CrossRef] [Green Version]
- Blumberg, D. Review: Date palm arthropod pests and their management in Israel. Phytoparasitica 2008, 36, 411–448. [Google Scholar] [CrossRef]
- Chaaban, S.B.; Chermiti, B.; Kreiter, S. Comparative demography of the spider mite, Oligonychus afrasiaticus, on four date palm varieties in Southwestern Tunisia. J. Insect Sci. 2011, 11, 136. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.J.; Jeong, G.; Han, J.H.; Lee, S. Biological control of aphid using fungal culture and culture filtrates of Beauveria bassiana. Mycobiology 2013, 41, 221–224. [Google Scholar] [CrossRef] [Green Version]
- Doberski, J.W. Comparative laboratory studies on three fungal pathogens of the elm bark beetle Scolytus scolytus: Pathogenicity of Beauveria bassiana, Metarhizium anisopliae, and Paecilomyces farinosus to larvae and adults of S. scolytus. J. Invertebr. Pathol. 1981, 37, 188–194. [Google Scholar] [CrossRef]
- Lord, J.C. Enhanced efficacy of Beauveria bassiana for red flour beetle with reduced moisture. J. Econ. Entomol. 2007, 100, 1071–1074. [Google Scholar] [CrossRef]
- C, M.M.; Lakshmi, K.A.; Devi, K.U. Laboratory evaluation of the pathogenicity of three isolates of the entomopathogenic fungus Beauveria bassiana (Bals.) Vuillemin on the American cockroach (Periplaneta americana). Biocontrol Sci. Technol. 1999, 9, 29–33. [Google Scholar]
- Inglis, G.; Johnson, D.; Goettel, M. Effects of temperature and sunlight on mycosis (Beauveria bassiana) (Hyphomycetes: Sympodulosporae) of Grasshoppers under field conditions. Environ. Entomol. 1997, 26, 400–409. [Google Scholar] [CrossRef] [Green Version]
- Hussain, A.; Tian, M.Y.; He, Y.R.; Ahmed, S. Entomopathogenic fungi disturbed the larval growth and feeding performance of Ocinara varians (Lepidoptera: Bombycidae) larvae. Insect Sci. 2009, 16, 511–517. [Google Scholar] [CrossRef]
- Vandenberg, J.D.; Shelton, A.M.; Wilsey, W.T.; Ramos, M. Assessment of Beauveria bassiana sprays for control of Diamondback moth (Lepidoptera: Plutellidae) on crucifers. J. Econ. Entomol. 1998, 91, 624–630. [Google Scholar] [CrossRef] [Green Version]
- Hussain, A.; Tian, M.-Y.; Wen, S.-Y. Exploring the caste-specific multi-layer defense mechanism of Formosan Subterranean Termites, Coptotermes formosanus Shiraki. Int. J. Mol. Sci. 2017, 18, 2694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, B.C.; Morisawa, T.A.; Newman, J.P.; Tjosvold, S.A.; Parrella, M.P. Fungal pathogen controls thrips in greenhouse flowers. Calif. Agric. 1998, 52, 32–36. [Google Scholar] [CrossRef] [Green Version]
- Hussain, A.; Rizwan-ul-Haq, M.; Al-Ayedh, H.; Ahmed, S.; Al-Jabr, A.M. Effect of Beauveria bassiana infection on the feeding performance and antioxidant defence of red palm weevil, Rhynchophorus ferrugineus. BioControl 2015, 60, 849–859. [Google Scholar] [CrossRef]
- Kim, C.-S.; Lee, J.-B.; Kim, B.-S.; Nam, Y.-H.; Shin, K.-S.; Kim, J.-W.; Kim, J.-E.; Kwon, G.-S. A Technique for the prevention of Greenhouse Whitefly (Trialeurodes vaporariorum) using the entomopathogenic fungus Beauveria bassiana M130. J. Microbiol. Biotechnol. 2014, 24, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dambolena, J.S.; Zunino, M.P.; Herrera, J.M.; Pizzolitto, R.P.; Areco, V.A.; Zygadlo, J.A. Terpenes: Natural Products for Controlling Insects of Importance to Human Health—A Structure-Activity Relationship Study. Psyche 2016, 2016, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Langsi, J.D.; Nukenine, E.N.; Oumarou, K.M.; Moktar, H.; Fokunang, C.N.; Mbata, G.N. Evaluation of the insecticidal activities of α-Pinene and 3-Carene on Sitophilus zeamais Motschulsky (Coleoptera: Curculionidae). Insects 2020, 11, 540. [Google Scholar] [CrossRef]
- Usha Pathipati, R.; Madhusudhana Murthy, J.; Sreedhar, B. Dynamic adsorption of α-pinene and linalool on silica nanoparticles for enhanced antifeedant activity against agricultural pests. J. Pest. Sci. 2014, 87, 191–200. [Google Scholar] [CrossRef]
- Gonçalves Diniz, A.; Barbosa, L.F.S.; Santos, A.C.S.; Oliveira, N.T.; da Costa, A.F.; Carneiro-Leão, M.P.; Tiago, P.V. Bio-insecticide effect of isolates of Fusarium caatingaense (Sordariomycetes: Hypocreales) combined to botanical extracts against Dactylopius opuntiae (Hemiptera: Dactylopiidae). Biocontrol Sci. Technol. 2020, 384–395. [Google Scholar] [CrossRef]
- Hernández, M.M.; Martínez-Villar, E.; Peace, C.; Pérez-Moreno, I.; Marco, V. Compatibility of the entomopathogenic fungus Beauveria bassiana with flufenoxuron and azadirachtin against Tetranychus urticae. Exp. Appl. Acarol. 2012, 58, 395–405. [Google Scholar] [CrossRef]
- Akbar, W.; Lord, J.C.; Nechols, J.R.; Loughin, T.M. Efficacy of Beauveria bassiana for red flour beetle when applied with plant essential oils or in mineral oil and organosilicone carriers. J. Econ. Entomol. 2005, 98, 683–688. [Google Scholar] [CrossRef] [PubMed]
- AlJabr, A.; Hussain, A.; Rizwan-ul-haq, M. Toxin-Pathogen synergy reshaping detoxification and antioxidant defense mechanism of Oligonychus afrasiaticus (McGregor). Molecules 2018, 23, 1978. [Google Scholar] [CrossRef] [Green Version]
- Schumacher, V.; Poehling, H.-M. In vitro effect of pesticides on the germination, vegetative growth, and conidial production of two strains of Metarhizium anisopliae. Fungal Biol. 2012, 116, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Statistix Statistix 8.1 Tallahassee, FL: Analytical Software 2003. Available online: https://www.amazon.com/Statistix-Analytical-Software-Users-Manual/dp/1881789063 (accessed on 25 April 2021).
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- StatsDirect StatsDirect Statistical Software; England StatsDirect Ltd.: Merseyside, UK, 2013; Available online: http://www.statsdirect.com (accessed on 25 April 2021).
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Hussain, A.; Rizwan-ul-Haq, M.; AlJabr, A.M.; Al-Ayedh, H. Evaluation of host–pathogen interactions for selection of entomopathogenic fungal isolates against Oligonychus afrasiaticus (McGregor). BioControl 2020, 65, 185–195. [Google Scholar] [CrossRef]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S-transferases. The first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef]
- Hussain, A.; Rizwan-ul-Haq, M.; Al-Ayedh, H.; AlJabr, A. Susceptibility and immune defence mechanisms of Rhynchophorus ferrugineus (Olivier) (Coleoptera: Curculionidae) against entomopathogenic fungal infections. Int. J. Mol. Sci. 2016, 17, 1518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, Y.; Hussain, A.; Guan, Z.; Wang, D.; Jaleel, W.; Lyu, L.; He, Y. Unraveling the mode of action of Cordyceps fumosorosea: Potential biocontrol agent against Plutella xylostella (Lepidoptera: Plutellidae). Insects 2021, 12, 179. [Google Scholar] [CrossRef]
- Hussain, A.; Tian, M.-Y.; Wen, S.-Y. Proteomic analysis of Formosan Subterranean Termites during exposure to entomopathogenic fungi. Curr. Proteom. 2018, 15, 229–240. [Google Scholar] [CrossRef]
- Hussain, A. Reprogramming the virulence: Insect defense molecules navigating the epigenetic landscape of Metarhizium robertsii. Virulence 2018, 9, 447–449. [Google Scholar] [CrossRef] [Green Version]
- Wraight, S.P.; Ramos, M.E. Synergistic interaction between Beauveria bassiana and Bacillus thuringiensis tenebrionis-based biopesticides applied against field populations of Colorado potato beetle larvae. J. Invertebr. Pathol. 2005, 90, 139–150. [Google Scholar] [CrossRef]
- Hussain, A.; Ali, M.W.; AlJabr, A.M.; Al-Kahtani, S.N. Insights into the Gryllus bimaculatus immune-related transcriptomic profiling to combat naturally invading pathogens. J. Fungi 2020, 6, 232. [Google Scholar] [CrossRef]
- Dannon, H.F.; Dannon, A.E.; Douro-Kpindou, O.K.; Zinsou, A.V.; Houndete, A.T.; Toffa-Mehinto, J.; Elegbede, I.A.T.M.; Olou, B.D.; Tamo, M. Toward the efficient use of Beauveria bassiana in integrated cotton insect pest management. J. Cott. Res. 2020, 3, 1–21. [Google Scholar]
- Al-mazra’awi, M.S.; Al-Abbadi, A.; Shatnawi, M.A.; Ateyyat, M. Effect of application method on the interaction between Beauveria bassiana and neem tree extract when combined for Thrips tabaci(Thysanoptera: Thripidae) control. J. Food Agric. Environ. 2009, 7, 869–873. [Google Scholar]
- Dampc, J.; Kula-Maximenko, M.; Molon, M.; Durak, R. Enzymatic defense response of apple aphid Aphis pomi to increased temperature. Insects 2020, 11, 436. [Google Scholar] [CrossRef]
- AlJabr, A.M.; Hussain, A.; Rizwan-ul-Haq, M.; Al-Ayedh, H. Toxicity of Plant Secondary Metabolites Modulating Detoxification Genes Expression for Natural Red Palm Weevil Pesticide Development. Molecules 2017, 22, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, A.; Rizwan-ul-Haq, M.; Al-Ayedh, H.; Aljabr, A.M. Toxicity and detoxification mechanism of black pepper and its major constituent in controlling Rhynchophorus ferrugineus Olivier (Curculionidae: Coleoptera). Neotrop. Entomol. 2017, 46, 685–693. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Ma, C.; Luo, J.; Niu, L.; Hua, H.; Zhang, S.; Cui, J. Potential of Cucurbitacin B and Epigallocatechin Gallate as Biopesticides against Aphis gossypii. Insects 2021, 12, 32. [Google Scholar] [CrossRef] [PubMed]
- Badawy, M.E.I.; Nasr, H.M.; Rabea, E.I. Toxicity and biochemical changes in the honey bee Apis mellifera exposed to four insecticides under laboratory conditions. Apidologie 2015, 46, 177–193. [Google Scholar] [CrossRef] [Green Version]
- Zibaee, A.; Bandani, A.R. A study on the toxicity of a medicinal plant, Artemisia annua L. (Asteracea) extracts to the Sunn pest, Eurygaster integriceps Puton (Hemiptera: Scutelleridae). J. Plant Prot. Res. 2010, 50, 79–85. [Google Scholar] [CrossRef]


{kind=link}
| Biological Index 1 | Criterion | Classification |
|---|---|---|
| Conidia vs. (+)-α-Pinene | >66 | Compatible |
| 42–66 | Moderately Toxic | |
| <42 | Toxic |
| (+)-α-Pinene (mg/mL). | Scheme I | Scheme II | Scheme III | Scheme IV | Conidia (mg/mL) | ||||
|---|---|---|---|---|---|---|---|---|---|
| Share of Category Weights | Share of Category Weights | Share of Category Weights | Share of Category Weights | ||||||
| 20% (+)-α-Pinene: 80% Conidia | 40% (+)-α-Pinene: 60% Conidia | 60% (+)-α-Pinene: 40% Conidia | 80% (+)-α-Pinene: 20% Conidia | ||||||
| (+)-α-Pinene (mg/mL) | Conidia mg/mL | (+)-α-Pinene (mg/mL) | Conidia mg/mL | (+)-α-Pinene (mg/mL) | Conidia mg/mL | (+)-α-Pinene (mg/mL) | Conidia mg/mL | ||
| 0.7 | 0.14 | 3.2 | 0.28 | 2.4 | 0.42 | 1.6 | 0.56 | 0.8 | 4 |
| 1.4 | 0.28 | 6.4 | 0.56 | 4.8 | 0.84 | 3.2 | 1.12 | 1.6 | 8 |
| 2.1 | 0.42 | 9.6 | 0.84 | 7.2 | 1.26 | 4.8 | 1.68 | 2.4 | 12 |
| 2.8 | 0.56 | 12.8 | 1.12 | 9.6 | 1.68 | 6.4 | 2.24 | 3.2 | 16 |
| 3.5 | 0.70 | 16.0 | 1.40 | 12.0 | 2.10 | 8.0 | 2.80 | 4.0 | 20 |
| Treatments (mg/mL) | Germination (%) * | Vegetative Growth (mm) * | Conidiation (×107 Conidia/mL) * | Biological Index | Classification |
|---|---|---|---|---|---|
| Control | 98.90 ± 0.50 a | 86.60 ± 1.98 a | 7.90 ± 0.66 a | - | - |
| 0.7 | 98.00 ± 0.96 ab | 85.40 ± 2.32 ab | 7.20 ± 0.59 ab | 95 | Compatibility |
| 1.4 | 97.60 ± 0.76 ab | 83.10 ± 1.69 ab | 6.70 ± 0.54 abc | 91 | Compatibility |
| 2.1 | 96.40 ± 0.80 bc | 80.40 ± 1.83 bc | 6.40 ± 0.56 abc | 88 | Compatibility |
| 2.8 | 95.90 ± 0.98 bc | 76.80 ± 2.06 cd | 5.80 ± 0.47 bc | 83 | Compatibility |
| 3.5 | 95.20 ± 1.19 c | 74.20 ±1.69 d | 5.30 ± 0.30 c | 79 | Compatibility |
| Treatments | Post-Exposure Duration | ||
|---|---|---|---|
| 2 d | 4 d | 6 d | |
| (+)-α-Pinene | (%) | (%) | (%) |
| 0.7 mg/mL | 08.40 ± 0.75 i | 12.40 ± 0.75 h | 24.40 ± 1.33 fg |
| 1.4 mg/mL | 12.40 ± 0.98 h | 21.60 ± 1.33 g | 32.40 ± 2.14 e |
| 2.1 mg/mL | 23.60 ± 1.72 g | 28.80 ± 2.06 ef | 49.20 ± 2.87 c |
| 2.8 mg/mL | 27.60 ± 1.72 ef | 42.40 ± 1.83 d | 63.20 ± 2.33 b |
| 3.5 mg/mL | 31.60 ± 1.94 e | 56.80 ± 2.15 b | 79.60 ± 2.48 a |
| Scheme I: 20% (+)-α-Pinene: 80% Conidia | |||
| 0.14 mg/mL (+)-α-Pinene + 3.2 mg/mL conidia | 19.60 ± 1.17 k | 36.40 ± 1.94 h | 66.40 ± 1.72 e |
| 0.28 mg/mL (+)-α-Pinene + 6.4 mg/mL conidia | 24.40 ± 1.17 j | 43.20 ± 2.06 g | 72.40 ± 2.99 d |
| 0.42 mg/mL (+)-α-Pinene + 9.6 mg/mL conidia | 28.00 ± 1.10 i | 54.40 ± 1.60 f | 77.60 ± 2.86 c |
| 0.56 mg/mL (+)-α-Pinene + 12.8 mg/mL conidia | 29.20 ± 1.74 i | 65.60 ± 3.06 e | 83.60 ± 2.79 b |
| 0.70 mg/mL (+)-α-Pinene + 16.0 mg/mL conidia | 32.80 ± 2.15 h | 76.40 ± 2.48 cd | 87.60 ± 2.71 a |
| Scheme II: 40% (+)-α-Pinene: 60% Conidia | |||
| 0.28 mg/mL (+)-α-Pinene + 2.4 mg/mL conidia | 21.60 ± 1.17 l | 41.60 ± 1.72 h | 70.40 ± 2.14 f |
| 0.56 mg/mL (+)-α-Pinene + 4.8 mg/mL conidia | 23.60 ± 1.60 l | 43.20 ± 2.06 h | 80.80 ± 2.42 de |
| 0.84 mg/mL (+)-α-Pinene + 7.2 mg/mL conidia | 28.40 ± 1.47 k | 61.20 ± 2.15 g | 84.40 ± 2.14 c |
| 1.12 mg/mL (+)-α-Pinene + 9.6 mg/mL conidia | 31.60 ± 1.72 j | 77.60 ± 2.48 e | 89.60 ± 2.14 b |
| 1.40 mg/mL (+)-α-Pinene + 12.0 mg/mL conidia | 34.80 ± 1.62 i | 81.60 ± 2.56 cd | 92.80 ± 1.85 a |
| Scheme III: 60% (+)-α-Pinene: 40% Conidia | |||
| 0.42 mg/mL (+)-α-Pinene + 1.6 mg/mL conidia | 23.60 ± 1.17 j | 46.60 ± 2.14 e | 77.20 ± 1.85 c |
| 0.84 mg/mL (+)-α-Pinene + 3.2 mg/mL conidia | 27.60 ± 1.47 i | 71.20 ± 2.58 d | 87.60 ± 2.32 b |
| 1.26 mg/mL (+)-α-Pinene + 4.8 mg/mL conidia | 34.40 ± 1.60 h | 87.20 ± 2.58 b | 96.40 ± 0.98 a |
| 1.68 mg/mL (+)-α-Pinene + 6.4 mg/mL conidia | 38.40 ± 1.47 g | 97.60 ± 0.75 a | 98.80 ± 0.49 a |
| 2.10 mg/mL (+)-α-Pinene + 8.0 mg/mL conidia | 40.40 ± 1.60 f | 98.80 ± 0.49 a | 99.20 ± 0.49 a |
| Scheme IV: 80% (+)-α-Pinene: 20% Conidia | |||
| 0.56 mg/mL (+)-α-Pinene + 0.8 mg/mL conidia | 24.40 ± 1.60 h | 51.60 ± 1.72 d | 80.80 ± 1.85 c |
| 1.12 mg/mL (+)-α-Pinene + 1.6 mg/mL conidia | 29.60 ± 1.60 g | 88.00 ± 3.46 b | 96.80 ± 0.80 a |
| 1.68 mg/mL (+)-α-Pinene + 2.4 mg/mL conidia | 36.00 ± 1.67 f | 94.80 ± 2.15 a | 97.20 ± 1.02 a |
| 2.24 mg/mL (+)-α-Pinene + 3.2 mg/mL conidia | 42.80 ± 1.32 e | 98.80 ± 0.80 a | 98.80 ± 0.80 a |
| 2.80 mg/mL (+)-α-Pinene + 4.0 mg/mL conidia | 43.20 ± 1.85 e | 99.20 ± 0.49 a | 99.60 ± 0.40 a |
| B. bassiana ARSEF 8465 Conidia | |||
| 4 mg/mL | 6.40 ± 0.75 j | 15.60 ± 1.17 hi | 24.40 ± 1.72 ef |
| 8 mg/mL | 12.80 ± 1.02 i | 20.80 ± 1.02 fg | 31.60 ± 1.17 d |
| 12 mg/mL | 19.20 ± 1.02 gh | 32.40 ± 3.97 d | 42.40 ± 1.72 c |
| 16 mg/mL | 22.40 ± 1.47 f | 44.40 ± 1.72 c | 52.80 ± 2.33 b |
| 20 mg/mL | 26.40 ± 1.33 de | 56.40 ± 2.79 b | 69.60 ± 2.79 a |
| Schemes | * LC50 (mg/mL) | Joint Toxicity | Interaction ** |
|---|---|---|---|
| Scheme IV: 80% (+)-α-Pinene: 20% B. bassiana ARSEF 8465 Conidia | |||
| 0.56 mg/mL (+)-α-Pinene + 0.8 mg/mL conidia | 1.32 (1.09 to 1.51) | 755 | Synergistic |
| 1.12 mg/mL (+)-α-Pinene + 1.6 mg/mL conidia | |||
| 1.68 mg/mL (+)-α-Pinene + 2.4 mg/mL conidia | |||
| 2.24 mg/mL (+)-α-Pinene + 3.2 mg/mL conidia | |||
| 2.80 mg/mL (+)-α-Pinene + 4.0 mg/mL conidia | |||
| Scheme III: 60% (+)-α-Pinene: 40% B. bassiana ARSEF 8465 Conidia | |||
| 0.42 mg/mL (+)-α-Pinene + 1.6 mg/mL conidia | 2.40 (2.02 to 2.74) | 280 | Synergistic |
| 0.84 mg/mL (+)-α-Pinene + 3.2 mg/mL conidia | |||
| 1.26 mg/mL (+)-α-Pinene + 4.8 mg/mL conidia | |||
| 1.68 mg/mL (+)-α-Pinene + 6.4 mg/mL conidia | |||
| 2.10 mg/mL (+)-α-Pinene + 8.0 mg/mL conidia | |||
| Scheme II: 40% (+)-α-Pinene: 60% B. bassiana ARSEF 8465 Conidia | |||
| 0.28 mg/mL (+)-α-Pinene + 2.4 mg/mL conidia | 4.20 (3.16 to 5.07) | 121 | Synergistic |
| 0.56 mg/mL (+)-α-Pinene + 4.8 mg/mL conidia | |||
| 0.84 mg/mL (+)-α-Pinene + 7.2 mg/mL conidia | |||
| 1.12 mg/mL (+)-α-Pinene + 9.6 mg/mL conidia | |||
| 1.40 mg/mL (+)-α-Pinene + 12.0 mg/mL conidia | |||
| Scheme I: 20% (+)-α-Pinene: 80% B. bassiana ARSEF 8465 Conidia | |||
| 0.14 mg/mL (+)-α-Pinene + 3.2 mg/mL conidia | 7.06 (5.63 to 8.44) | 58 | Antagonistic |
| 0.28 mg/mL (+)-α-Pinene + 6.4 mg/mL conidia | |||
| 0.42 mg/mL (+)-α-Pinene + 9.6 mg/mL conidia | |||
| 0.56 mg/mL (+)-α-Pinene + 12.8 mg/mL conidia | |||
| 0.70 mg/mL (+)-α-Pinene + 16.0 mg/mL conidia | |||
| Treatments | Relative Enzymatic Activities (%) | |||
|---|---|---|---|---|
| CAT | SOD | GST | AchE | |
| B. bassiana ARSEF 8465 Conidia | ||||
| 4 mg/mL | 2.67 ± 0.23 n | 9.67 ± 0.25 opq | 7.69 ± 0.74 no | 1.18 ± 0.29 m |
| 8 mg/mL | 4.45 ± 0.28 n | 11.53 ± 0.21 no | 11.66 ± 0.93 klm | 2.62 ± 0.37 jklm |
| 12 mg/mL | 8.33 ± 0.98 m | 15.20 ± 0.20 lm | 13.49 ± 1.24 ijk | 3.92 ± 0.48 hij |
| 16 mg/mL | 9.06 ± 0.71 m | 17.87 ± 1.42 k | 15.38 ± 1.97 hij | 5.44 ± 0.65 fgh |
| 20 mg/mL | 10.44 ± 0.69 m | 19.13 ± 0.60 k | 21.87 ± 0.34 g | 12.98 ± 1.52 d |
| (+)-α-Pinene | ||||
| 0.7 mg/mL | 19.55 ± 0.83 k | 1.24 ± 0.13 u | 9.04 ± 0.45 mn | 2.14 ± 0.35 klm |
| 1.4 mg/mL | 23.27 ± 0.44 j | 1.79 ± 0.15 tu | 16.02 ± 1.59 hi | 3.58 ± 0.30 ijk |
| 2.1 mg/mL | 28.07 ± 0.62 i | 7.09 ± 1.22 qr | 21.87 ± 0.34 g | 5.92 ± 0.78 fg |
| 2.8 mg/mL | 35.71 ± 1.04 h | 9.67 ± 0.25 opq | 26.78 ± 0.32 f | 9.11 ± 0.48 e |
| 3.5 mg/mL | 36.46 ± 1.04 h | 14.17 ± 1.62 mn | 39.24 ± 1.20 d | 14.62 ± 1.08 d |
| Scheme I: 20% (+)-α-Pinene: 80% Conidia | ||||
| 0.14 mg/mL (+)-α-Pinene + 3.2 mg/mL conidia | 10.85 ± 1.99 m | 3.33 ± 0.55 stu | 4.59 ± 0.88 o | 1.59 ± 0.36 lm |
| 0.28 mg/mL (+)-α-Pinene + 6.4 mg/mL conidia | 14.80 ± 1.08 l | 4.37 ± 0.96 st | 7.40 ± 0.96 no | 2.18 ± 0.27 jklm |
| 0.42 mg/mL (+)-α-Pinene + 9.6 mg/mL conidia | 23.81 ± 0.91 j | 5.45 ± 0.57 rs | 10.08 ± 0.85 lmn | 3.56 ± 0.47 ijk |
| 0.56 mg/mL (+)-α-Pinene + 12.8 mg/mL conidia | 27.79 ± 0.73 i | 7.64 ± 1.19 pqr | 13.24 ± 0.87 ijk | 4.86 ± 0.59 ghi |
| 0.70 mg/mL (+)-α-Pinene + 16.0 mg/mL conidia | 29.12 ± 0.50 i | 9.32 ± 0.31 opq | 17.59 ± 1.01 h | 6.03 ± 0.31 fg |
| Scheme II: 40% (+)-α-Pinene: 60% Conidia | ||||
| 0.28 mg/mL (+)-α-Pinene + 2.4 mg/mL conidia | 43.08 ± 0.82 g | 9.79 ± 0.26 op | 13.05 ± 0.98 ijkl | 3.16 ± 0.63 ijkl |
| 0.56 mg/mL (+)-α-Pinene + 4.8 mg/mL conidia | 44.12 ± 0.96 g | 20.10 ± 0.24 k | 15.59 ± 1.28 hij | 7.13 ± 0.46 f |
| 0.84 mg/mL (+)-α-Pinene + 7.2 mg/mL conidia | 49.70 ± 0.63 f | 24.09 ± 0.50 j | 24.73 ± 1.12 fg | 9.04 ± 0.53 e |
| 1.12 mg/mL (+)-α-Pinene + 9.6 mg/mL conidia | 58.19 ± 0.81 d | 27.30 ± 0.69 i | 35.61 ± 2.84 e | 13.41 ± 0.83 d |
| 1.40 mg/mL (+)-α-Pinene + 12.0 mg/mL conidia | 63.86 ± 1.89 c | 39.06 ± 0.56 g | 44.30 ± 2.42 c | 19.61 ± 1.10 c |
| Scheme III: 60% (+)-α-Pinene: 40% Conidia | ||||
| 0.42 mg/mL (+)-α-Pinene + 1.6 mg/mL conidia | 44.12 ± 0.96 g | 17.44 ± 0.32 kl | 12.67 ± 0.81 jkl | 6.49 ± 0.36 fg |
| 0.84 mg/mL (+)-α-Pinene + 3.2 mg/mL conidia | 49.89 ± 1.62 f | 26.54 ± 0.72 ij | 18.31 ± 0.86 h | 7.15 ± 0.44 f |
| 1.26 mg/mL (+)-α-Pinene + 4.8 mg/mL conidia | 57.30 ± 1.11 d | 35.71 ± 1.04 h | 27.93 ± 1.40 f | 9.90 ± 0.70 e |
| 1.68 mg/mL (+)-α-Pinene + 6.4 mg/mL conidia | 68.07 ± 0.98 b | 49.70 ± 0.63 e | 39.68 ± 1.47 d | 14.68 ± 0.76 d |
| 2.10 mg/mL (+)-α-Pinene + 8.0 mg/mL conidia | 71.84 ± 2.05 a | 57.47 ± 1.02 c | 53.18 ± 1.46 b | 22.85 ± 0.88 b |
| Scheme IV: 80% (+)-α-Pinene: 20% Conidia | ||||
| 0.56 mg/mL (+)-α-Pinene + 0.8 mg/mL conidia | 53.89 ± 1.11 e | 25.73 ± 1.83 ij | 17.59 ± 1.01 h | 9.71 ± 0.54 e |
| 1.12 mg/mL (+)-α-Pinene + 1.6 mg/mL conidia | 57.13 ± 1.21 d | 43.08 ± 0.82 f | 25.29 ± 1.53 f | 13.11 ± 0.74 d |
| 1.68 mg/mL (+)-α-Pinene + 2.4 mg/mL conidia | 66.27 ± 1.19 bc | 52.42 ± 0.57 d | 40.33 ± 1.30 d | 19.89 ± 0.99 c |
| 2.24 mg/mL (+)-α-Pinene + 3.2 mg/mL conidia | 66.61 ± 1.20 bc | 63.89 ± 1.88 b | 53.91 ± 1.29 b | 22.20 ± 0.64 b |
| 2.80 mg/mL (+)-α-Pinene + 4.0 mg/mL conidia | 72.36 ± 1.89 a | 75.22 ± 2.02 a | 61.09 ± 0.89 a | 30.07 ± 1.09 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hussain, A. Compatibility of Beauveria bassiana and a Plant Secondary Metabolite: A Novel Modeling Approach to Invade Host Defense for Effective Control of Oligonychus afrasiaticus (McGregor) on Date Palms. J. Fungi 2021, 7, 334. https://doi.org/10.3390/jof7050334
Hussain A. Compatibility of Beauveria bassiana and a Plant Secondary Metabolite: A Novel Modeling Approach to Invade Host Defense for Effective Control of Oligonychus afrasiaticus (McGregor) on Date Palms. Journal of Fungi. 2021; 7(5):334. https://doi.org/10.3390/jof7050334
Chicago/Turabian StyleHussain, Abid. 2021. "Compatibility of Beauveria bassiana and a Plant Secondary Metabolite: A Novel Modeling Approach to Invade Host Defense for Effective Control of Oligonychus afrasiaticus (McGregor) on Date Palms" Journal of Fungi 7, no. 5: 334. https://doi.org/10.3390/jof7050334
APA StyleHussain, A. (2021). Compatibility of Beauveria bassiana and a Plant Secondary Metabolite: A Novel Modeling Approach to Invade Host Defense for Effective Control of Oligonychus afrasiaticus (McGregor) on Date Palms. Journal of Fungi, 7(5), 334. https://doi.org/10.3390/jof7050334