
Participation of the ABC Transporter CDR1 in Azole Resistance of Candida lusitaniae



Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Media and Primers
2.2. Case Report of C. lusitaniae Infection
2.3. Deletion of CDR1
2.4. Restoration of MRR1 Alleles
2.5. Rhodamine 6G Efflux
2.6. qPCR Assays
2.7. Minimum Inhibitory Concentrations (MIC) Assays
2.8. Sequencing
3. Results
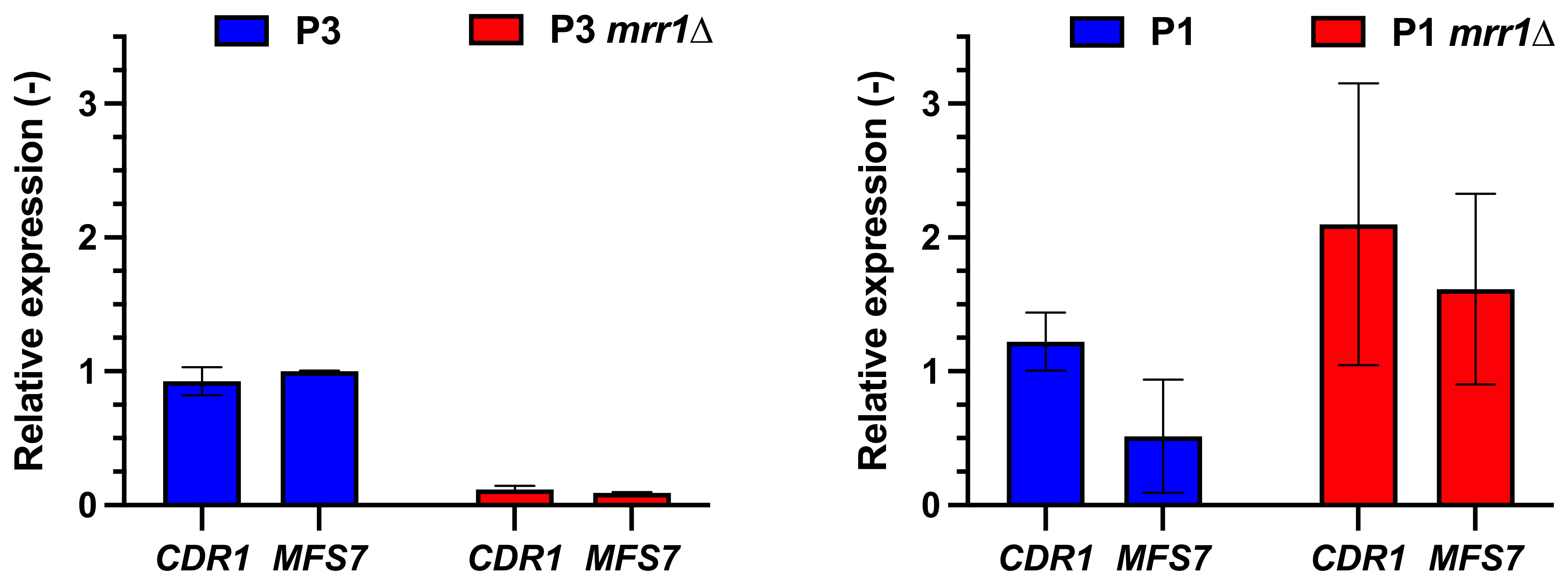
3.1. CDR1 Expression Patterns in C. lusitaniae
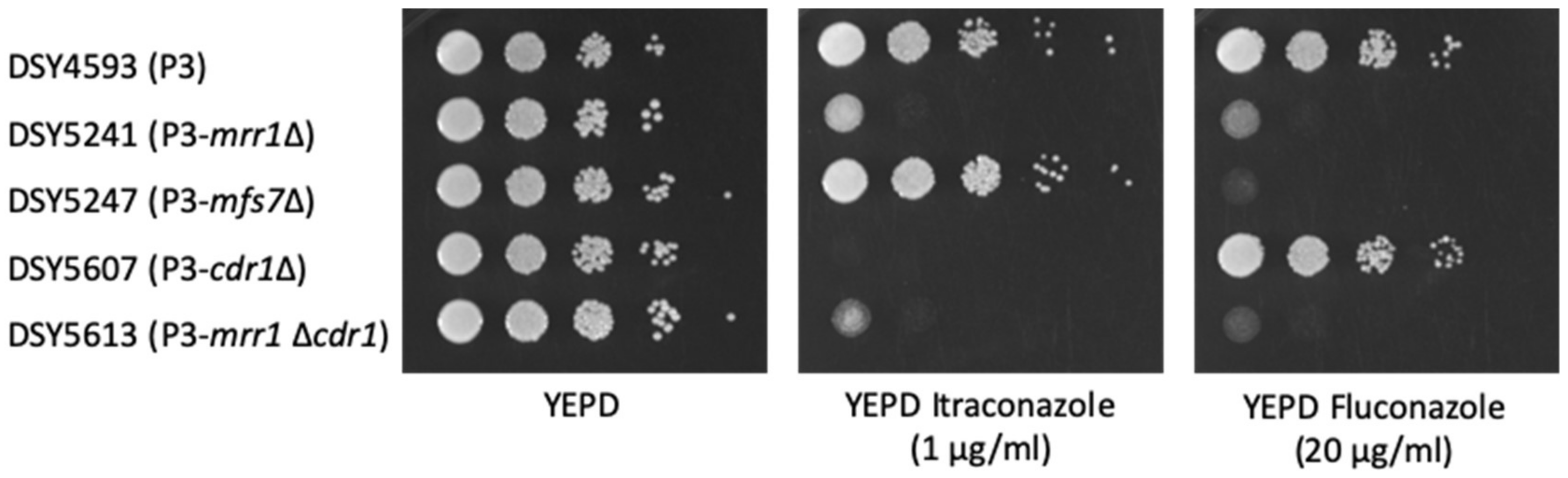
3.2. Effect of CDR1 on Antifungal Susceptibility
3.3. Probing Cdr1 Activity by R6G Efflux
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McCarty, T.P.; White, C.M.; Pappas, P.G. Candidemia and Invasive Candidiasis. Infect. Dis. Clin. N. Am. 2021, 35, 389–413. [Google Scholar] [CrossRef]
- Pfaller, M.A.; Andes, D.R.; Diekema, D.J.; Horn, D.L.; Reboli, A.C.; Rotstein, C.; Franks, B.; Azie, N.E. Epidemiology and Outcomes of Invasive Candidiasis Due to Non-albicans Species of Candida in 2,496 Patients: Data from the Prospective Antifungal Therapy (PATH) Registry 2004–2008. PLoS ONE 2014, 9, e101510. [Google Scholar] [CrossRef] [Green Version]
- Andes, D.R.; Safdar, N.; Baddley, J.W.; Playford, G.; Reboli, A.C.; Rex, J.H.; Sobel, J.D.; Pappas, P.G.; Kullberg, B.J.; Mycoses Study Group. Impact of Treatment Strategy on Outcomes in Patients with Candidemia and Other Forms of Invasive Candidiasis: A Patient-Level Quantitative Review of Randomized Trials. Clin. Infect. Dis. 2012, 54, 1110–1122. [Google Scholar] [CrossRef]
- Desnos-Ollivier, M.; Moquet, O.; Chouaki, T.; Guerin, A.M.; Dromer, F. Development of Echinocandin Resistance in Clavispora Lusitaniae during Caspofungin Treatment. J. Clin. Microbiol. 2011, 49, 2304–2306. [Google Scholar] [CrossRef] [Green Version]
- Demers, E.G.; Biermann, A.R.; Masonjones, S.; Crocker, A.W.; Ashare, A.; Stajich, J.E.; Hogan, D.A. Evolution of Drug Resistance in an Antifungal-Naive Chronic Candida lusitaniae Infection. Proc. Natl. Acad. Sci. USA 2018, 32, 201807698. [Google Scholar] [CrossRef] [Green Version]
- Asner, S.A.; Giulieri, S.; Diezi, M.; Marchetti, O.; Sanglard, D. Acquired Multidrug Antifungal Resistance in Candida lusitaniae during Therapy. Antimicrob. Agents Chemother. 2015, 59, 7715–7722. [Google Scholar] [CrossRef]
- Kannan, A.; Asner, S.A.; Trachsel, E.; Kelly, S.; Parker, J.; Sanglard, D. Comparative Genomics for the Elucidation of Multidrug Resistance in Candida lusitaniae. mBio 2019, 10, e02512-19. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, G.B.; Lorenz, A. What Do We Know about the Biology of the Emerging Fungal Pathogen of Humans Candida auris? Microbiol. Res. 2021, 242, 126621. [Google Scholar] [CrossRef]
- Sanglard, D. Emerging Threats in Antifungal-Resistant Fungal Pathogens. Front. Med. 2016, 3, 11. [Google Scholar] [CrossRef] [Green Version]
- Coste, A.T.; Karababa, M.; Ischer, F.; Bille, J.; Sanglard, D. TAC1, transcriptional activator of CDR genes, is a new transcription factor involved in the regulation of Candida albicans ABC transporters CDR1 and CDR2. Eukaryot. Cell 2004, 3, 1639–1652. [Google Scholar] [CrossRef] [Green Version]
- Morschhäuser, J.; Barker, K.S.; Liu, T.T.; BlaB-Warmuth, J.; Homayouni, R.; Rogers, P.D. The transcription factor Mrr1p controls expression of the MDR1 efflux pump and mediates multidrug resistance in Candida albicans. PLoS Pathog. 2007, 3, e164. [Google Scholar] [CrossRef] [Green Version]
- Demers, E.G.; Stajich, J.E.; Ashare, A.; Occhipinti, P.; Hogan, D.A. Balancing Positive and Negative Selection: In Vivo Evolution of Candida lusitaniae MRR1. Mbio 2021, 12. [Google Scholar] [CrossRef]
- Pappas, P.G.; Kauffman, C.A.; Andes, D.; Benjamin, D.K.; Calandra, T.F.; Edwards, J.E.; Filler, S.G.; Fisher, J.F.; Kullberg, B.-J.; Ostrosky-Zeichner, L.; et al. Clinical Practice Guidelines for the Management Candidiasis: 2009 Update by the Infectious Diseases Society of America. Clin. Infect. Dis. 2009, 48, 503–535. [Google Scholar] [CrossRef] [Green Version]
- Kaufman, D.; Boyle, R.; Hazen, K.C.; Patrie, J.T.; Robinson, M.; Donowitz, L.G. Fluconazole Prophylaxis against Fungal Colonization and Infection in Preterm Infants. N. Engl. J. Med. 2001, 345, 1660–1666. [Google Scholar] [CrossRef]
- Basso, L.R.; Bartiss, A.; Mao, Y.; Gast, C.E.; Coelho, P.S.R.; Snyder, M.; Wong, B. Transformation of Candida albicans with a synthetic hygromycin B resistance gene. Yeast 2010, 27, 1039–1048. [Google Scholar] [CrossRef] [Green Version]
- Jensen, M.A.; Fukushima, M.; Davis, R.W. DMSO and Betaine Greatly Improve Amplification of GC-Rich Constructs in De Novo Synthesis. PLoS ONE 2010, 5, e11024. [Google Scholar] [CrossRef] [Green Version]
- Grahl, N.; Demers, E.G.; Crocker, A.W.; Hogan, D.A. Use of RNA-Protein Complexes for Genome Editing in Non-albicans Candida Species. mSphere 2017, 2, e00218-17. [Google Scholar] [CrossRef] [Green Version]
- Coste, A.; Turner, V.; Ischer, F.; Morschhäuser, J.; Forche, A.; Selmecki, A.; Berman, J.; Bille, J.; Sanglard, D. A mutation in Tac1p, a transcription factor regulating CDR1 and CDR2, is coupled with loss of heterozygosity at chromosome 5 to mediate antifungal resistance in Candida albicans. Genetics 2006, 172, 2139–2156. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, K.; Niimi, M.; Niimi, K.; Holmes, A.R.; Yates, J.E.; Decottignies, A.; Monk, B.C.; Goffeau, A.; Cannon, R.D. Functional expression of Candida albicans drug efflux pump Cdr1p in a Saccharomyces cerevisiae strain deficient in membrane transporters. Antimicrob. Agents Chemother. 2001, 45, 3366–3374. [Google Scholar] [CrossRef] [Green Version]
- Sanglard, D.; Ischer, F.; Bille, J. Role of ATP-binding-cassette transporter genes in high-frequency acquisition of resistance to azole antifungals in Candida glabrata. Antimicrob. Agents Chemother. 2001, 45, 1174–1183. [Google Scholar] [CrossRef] [Green Version]
- Silva, L.V.; Sanguinetti, M.; Vandeputte, P.; Torelli, R.; Rochat, B.; Sanglard, D. Milbemycins: More than Efflux Inhibitors for Fungal Pathogens. Antimicrob. Agents Chemother. 2013, 57, 873–886. [Google Scholar] [CrossRef] [Green Version]
- Sanglard, D.; Kuchler, K.; Ischer, F.; Pagani, J.L.; Monod, M.; Bille, J. Mechanisms of Resistance to Azole Antifungal Agents in Candida albicans Isolates from AIDS Patients Involve Specific Multidrug Transporters. Antimicrob. Agents Chemother. 1995, 39, 2378–2386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monod, M.; Feuermann, M.; Salamin, K.; Fratti, M.; Makino, M.; Alshahni, M.M.; Makimura, K.; Yamada, T. Trichophyton Rubrum Azole Resistance Mediated by a New ABC Transporter, TruMDR3. Antimicrob. Agents Chemother. 2019, 63. [Google Scholar] [CrossRef] [Green Version]
- Walker, B.; Izumikawa, K.; Tsai, H.-F.; Bennett, J.E. Milbemycin A4 Oxime as a Probe of Azole Transport in Candida glabrata. FEMS Yeast Res. 2014, 14, 755–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cannon, R.D.; Lamping, E.; Holmes, A.R.; Niimi, K.; Baret, P.V.; Keniya, M.V.; Tanabe, K.; Niimi, M.; Goffeau, A.; Monk, B.C. Efflux-Mediated Antifungal Drug Resistance. Clin. Microbiol. Rev. 2009, 22, 291–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cowen, L.E.; Sanglard, D.; Howard, S.J.; Rogers, P.D.; Perlin, D.S. Mechanisms of Antifungal Drug Resistance. Cold Spring Harb. Perspect. Med. 2015, 5, a019752. [Google Scholar] [CrossRef]
- Liu, T.T.; Znaidi, S.; Barker, K.S.; Xu, L.; Homayouni, R.; Saidane, S.; Morschhäuser, J.; Nantel, A.; Raymond, M.; Rogers, P.D. Genome-wide expression and location analyses of the Candida albicans Tac1p regulon. Eukaryot. Cell 2007, 6, 2122–2138. [Google Scholar] [CrossRef] [Green Version]
- Schubert, S.; Popp, C.; Rogers, P.D.; Morschhauser, J. Functional dissection of a Candida albicans zinc cluster transcription factor, the multidrug resistance regulator Mrr1. Eukaryot. Cell 2011, 10, 1110–1121. [Google Scholar] [CrossRef] [Green Version]
- Arastehfar, A.; Hilmioğlu-Polat, S.; Daneshnia, F.; Pan, W.; Hafez, A.; Fang, W.; Liao, W.; Şahbudak-Bal, Z.; Metin, D.Y.; de Almeida Júnior, J.N.; et al. Clonal Candidemia Outbreak by Candida parapsilosis Carrying Y132F in Turkey: Evolution of a Persisting Challenge. Front. Cell. Infect. Microbiol. 2021, 11, 676177. [Google Scholar] [CrossRef]
- Paul, S.; Singh, S.; Sharma, D.; Chakrabarti, A.; Rudramurthy, S.M.; Ghosh, A.K. Dynamics of in Vitro Development of Azole Resistance in Candida tropicalis. J. Glob. Antimicrob. Resist. 2020, 22, 553–561. [Google Scholar] [CrossRef]
- Li, J.; Coste, A.T.; Liechti, M.; Bachmann, D.; Sanglard, D.; Lamoth, F. Novel ERG11 and TAC1b Mutations Associated with Azole Resistance in Candida auris. Antimicrob. Agents Chemother. 2021. [Google Scholar] [CrossRef]
- Rybak, J.M.; Muñoz, J.F.; Barker, K.S.; Parker, J.E.; Esquivel, B.D.; Berkow, E.L.; Lockhart, S.R.; Gade, L.; Palmer, G.E.; White, T.C.; et al. Mutations in TAC1B: A Novel Genetic Determinant of Clinical Fluconazole Resistance in Candida auris. mBio 2020, 11, e00365-20. [Google Scholar] [CrossRef]
- Mayr, E.-M.; Ramírez-Zavala, B.; Krüger, I.; Morschhäuser, J. A Zinc Cluster Transcription Factor Contributes to the Intrinsic Fluconazole Resistance of Candida auris. mSphere 2020, 5, e00279-20. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Rossi, J.M.; Myers, L.C. Candida albicans Zn Cluster Transcription Factors Tac1 and Znc1 Are Activated by Farnesol To Upregulate a Transcriptional Program Including the Multidrug Efflux Pump CDR1. Antimicrob. Agents Chemother. 2018, 62, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Coste, A.T.; Ramsdale, M.; Ischer, F.; Sanglard, D. Divergent functions of three Candida albicans zinc-cluster transcription factors (CTA4, ASG1 and CTF1) complementing pleiotropic drug resistance in Saccharomyces cerevisiae. Microbiology 2008, 154, 1491–1501. [Google Scholar] [CrossRef] [Green Version]
- Maesaki, S.; Marichal, P.; Bossche, H.V.; Sanglard, D.; Kohno, S. Rhodamine 6G efflux for the detection of CDR1-overexpressing azole-resistant Candida albicans strains. J. Antimicrob. Chemother. 1999, 44, 27–31. [Google Scholar] [CrossRef]
- Reboutier, D.; Piednoël, M.; Boisnard, S.; Conti, A.; Chevalier, V.; Florent, M.; Gibot-Leclerc, S.; Silva, B.D.; Chastin, C.; Fallague, K.; et al. Combination of Different Molecular Mechanisms Leading to Fluconazole Resistance in a Candida lusitaniae Clinical Isolate. Diagn. Microbiol. Infect. Dis. 2009, 63, 188–193. [Google Scholar] [CrossRef]
- Sanglard, D.; Ischer, F.; Monod, M.; Bille, J. Cloning of Candida albicans genes conferring resistance to azole antifungal agents: Characterization of CDR2, a new multidrug ABC transporter gene. Microbiology 1997, 143 Pt 2, 405–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Fold-Change (vs. Azole-Susceptible Isolates) | Experimental Condition | Reference |
|---|---|---|---|
| DSY4593 (P3) | 2,4 | RNAseq | [7] |
| DSY4661 (P4) | 4,7 | RNAseq | [7] |
| U04 | 2,6 a | RNAseq | [12] |
| DSY4593 (P3) | 5,0 | qPCR | [6] |
| DSY4661 (P4) | 3,5 | qPCR | [6] |
| Antifungal Drugs | MIC (µg/mL) | |||||
|---|---|---|---|---|---|---|
| P3 | P3-mfs7Δ | P3-cdr1Δ | P3-mrr1Δ | P3-mfs7Δcdr1Δ | P3-mrr1Δcdr1Δ | |
| Anidulafungin | 0.12 | 0.25 | 0.12 | 0.12 | 0.12 | 0.12 |
| Micafungin | 0.12 | 0.12 | 0.12 | 0.06 | 0.06 | 0.03 |
| Caspofungin | 0.12 | 0.25 | 0.12 | 0.12 | 0.12 | 0.12 |
| 5-Flucytosine | 64 | 4 | 32 | 0.5 | 0.5 | 0.5 |
| Posaconazole | 0.25 | 0.25 | 0.03 | 0.015 | 0.015 | 0.015 |
| Voriconazole | 0.5 | 0.12 | 0.25 | 0.015 | <0.008 | 0.015 |
| Itraconazole | 0.5 | 0.5 | 0.06 | 0.06 | 0.03 | 0.06 |
| Fluconazole | 32 | 4 | 32 | 1 | <0.12 | 1 |
| Amphotericin B | 0.5 | 0.5 | 0.5 | 0.25 | 0.25 | 0.25 |
| Antifungal Drugs | MIC (µg/mL) | |||||
|---|---|---|---|---|---|---|
| P1 | P1-mfs7Δ | P1-cdr1Δ | P1-mrr1Δ | P1-mfs7Δcdr1Δ | P1-mrr1Δcdr1Δ | |
| Anidulafungin | 0.12 | 0.06 | 0.12 | 0.12 | 0.06 | 0.12 |
| Micafungin | 0.06 | 0.06 | 0.06 | 0.03 | 0.12 | 0.06 |
| Caspofungin | 0.12 | 0.25 | 0.12 | 0.12 | 0.25 | 0.12 |
| 5-Flucytosine | 4 | 0.5 | 4 | 1 | 0.5 | 1 |
| Posaconazole | 0.03 | 0.015 | 0.03 | 0.015 | 0.015 | 0.015 |
| Voriconazole | <0.008 | <0.008 | <0.008 | <0.008 | <0.008 | <0.008 |
| Itraconazole | 0.06 | 0.06 | 0.06 | 0.06 | 0.03 | <0.008 |
| Fluconazole | 0.5 | 0.25 | 0.25 | 1 | <0.12 | 1 |
| Amphotericin B | 0.25 | 0.5 | 0.25 | 0.25 | 0.25 | 0.25 |
| Antifungal Drugs | MIC (µg/mL) | |||
|---|---|---|---|---|
| P1 | P1-AOx3 | P3 | P3-AOx3 | |
| Anidulafungin | 0.12 | 0.06 | 0.12 | 0.06 |
| Micafungin | 0.06 | 0.06 | 0.06 | 0.06 |
| Caspofungin | 0.12 | 0.25 | 0.12 | 0.06 |
| 5-Flucytosine | 4 | 0.5 | 64 | 64 |
| Posaconazole | 0.03 | 0.015 | 0.25 | <0.008 |
| Voriconazole | <0.008 | <0.008 | 0.5 | 0.25 |
| Itraconazole | 0.06 | 0.06 | 0.5 | <0.008 |
| Fluconazole | 0.5 | 0.25 | 32 | 16 |
| Amphotericin B | 0.25 | 0.5 | 0.5 | 0.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borgeat, V.; Brandalise, D.; Grenouillet, F.; Sanglard, D. Participation of the ABC Transporter CDR1 in Azole Resistance of Candida lusitaniae. J. Fungi 2021, 7, 760. https://doi.org/10.3390/jof7090760
Borgeat V, Brandalise D, Grenouillet F, Sanglard D. Participation of the ABC Transporter CDR1 in Azole Resistance of Candida lusitaniae. Journal of Fungi. 2021; 7(9):760. https://doi.org/10.3390/jof7090760
Chicago/Turabian StyleBorgeat, Valentin, Danielle Brandalise, Frédéric Grenouillet, and Dominique Sanglard. 2021. "Participation of the ABC Transporter CDR1 in Azole Resistance of Candida lusitaniae" Journal of Fungi 7, no. 9: 760. https://doi.org/10.3390/jof7090760
APA StyleBorgeat, V., Brandalise, D., Grenouillet, F., & Sanglard, D. (2021). Participation of the ABC Transporter CDR1 in Azole Resistance of Candida lusitaniae. Journal of Fungi, 7(9), 760. https://doi.org/10.3390/jof7090760